
Abstract
The Haemophilus parasuis respiratory tract pathogen poses a severe threat to the swine industry despite available antimicrobial therapies. To gain a more detailed understanding of the molecular mechanisms underlying H. parasuis response to tilmicosin treatment, microarray technology was applied to analyze the variation in gene expression of isolated H. parasuis SH0165 treated in vitro with subinhibitory (0.25 μg/ml) and inhibitory (8 μg/ml) concentrations. Tilmicosin treatment induced differential expression of 405 genes, the encoded products of which are mainly involved in the heat shock response, protein synthesis, and intracellular transportation. The subinhibitory and inhibitory concentrations of tilmicosin induced distinctive gene expression profiles of shared and unique changes, respectively. These changes included 302 genes mainly involved in protein export and the phosphotransferase system to sustain cell growth, and 198 genes mainly related to RNA polymerase, recombination, and repair to inhibit cell growth. In silico analysis of functions related to the differentially expressed genes suggested that adaptation of H. parasuis SH0165 to tilmicosin involves modulation of protein synthesis and membrane transport. Collectively, the genes comprising each transcriptional profile of H. parasuis response to tilmicosin provide novel insights into the physiological functions of this economically significant bacterium and may represent targets of future molecular therapeutic strategies.
Introduction
The H. parasuis SH0165 strain has emerged as the dominant serovar in China, and is recognized for its remarkably high virulence. 58 However, several other serovars, as well as nontypeable isolates, persist in China and around the world, limiting the efficacy of available vaccines and complicating the development of a cross-protective vaccine. 36 Thus, the swine industry continues to rely on antibiotic therapy for the treatment of Glässer's disease. It is not surprising that antimicrobial-resistant strains of H. parasuis have been isolated, including those resistant to one of the preferred veterinary antibiotics for treating respiratory pathogens, tilmicosin (TIM, Pulmotil®).10,59 TIM, a semisynthetic tylosin that is produced as a 16-membered macrolide during the fermentation of Streptomyces fradiae, 37 is known to function as a ribosomal inhibitor, inducing the transcription of bacterial-encoded ribosomal genes. 34 Gaining a detailed understanding of the global and context-specific transcriptional response of H. parasuis to TIM will likely reveal promising therapeutic targets to combat TIM resistance.
DNA microarrays represent a useful tool for rapidly identifying large numbers of genetic determinants simultaneously. In microbiology, DNA microarrays have provided detailed genomic level information related to context-specific pathogenic mechanisms, including identification of virulence factors and antibiotic-resistance genes. 55 It is well known that the extent of a bacterium's mechanistic response to an antibiotic attack can be time- and dose-dependent.19,50 In this study, we used a whole-genome DNA microarray to profile the transcriptional response induced in H. parasuis SH0165 upon exposure to TIM at subinhibitory and inhibitory concentrations and durations. These context-specific data provide insights into the molecular pathways underlying H. parasuis resistance to TIM.
Materials and Methods
Strains and antibiotics
The H. parasuis SH0165 strain used for the Genome Chip analysis was isolated from the lung of a diseased piglet in Shijiazhuang, China, in 2001. 58 The strain was treated with various concentrations and durations of tilmicosin (China Institute of Veterinary Drugs Control, Beijing, China) before transcriptional profiling, as detailed below. The tilmicosin stock was prepared in 95% ethanol, and working concentrations were prepared by dilution with sterile water.
Determination of the subinhibitory and inhibitory concentrations of tilmicosins
Before the transcriptional profiling study, we determined the subinhibitory concentration and the inhibitory concentration for H. parasuis SH0165 using a method previously described by Muthaiyan et al. 32 and the MIC (2 μg/ml) for H. somnus strain ATCC® 700025 published by the National Committee for Clinical Laboratory Standards. 33 The determinations were carried out using the fastidious medium [a tryptic soy broth (TSB; Difco Laboratories, Detroit, MI), 5% bovine serum, and 10 μg/ml NAD (Sigma, St. Louis, MO)]. Cultures were grown overnight at 37°C with shaking (180 rpm) until the optical density at 600 nm (OD600) reached 0.05 (Synergy HT spectrophotometer; Bio-tek, Winooski, VT). At that time, an aliquot was used to inoculate media solutions supplemented with 0.25, 1, 2, or 8 μg/ml tilmicosin. The bacteria were again grown at 37°C with shaking (180 rpm) and 50-μl samples were collected every 2 hrs over a 12-hr period for OD600 measurement. Each determination was made in triplicate.
Transcriptional profiling
Overnight cultures of H. parasuis SH0165 were inoculated (1% vol/vol) in the TSB medium (20 ml) and grown at 37°C with shaking (180 rpm). Growth was measured at regular intervals until the OD600 reached >0.3 (∼6×106 colony-forming units), at which time the cells were added to a medium with the maximal inhibitory concentration (8 μg/ml) of tilmicosin and incubated at 37°C for 50 min, 45 or a medium containing the subinhibitory concentration (0.25 μg/ml) of tilmicosin for ∼16 hrs.28,29 Control cultures consisted of H. parasuis SH0165 at OD600 >0.3 that were inoculated in the medium alone, without tilmicosin, and grown in similar conditions until the OD600 reached >0.3.
RNA isolation and microarray processing
Bacterial cultures (subinhibitory- and inhibitory-treated, and untreated control) were centrifuged for 5 min (2300 g at 4°C). Total RNA was extracted from the bacterial pellets by treatment with the TRIzol Reagent (Invitrogen Life Technologies, Carlsbad, CA) according to the manufacturer's instructions, and purified using the Qiagen RNeasy® Mini Kit (Hilden, Germany). RNA concentration and integrity were evaluated by A260/A280 spectrophotometer readings and agarose gel electrophoresis, respectively.
Microarray probes used in this study were in situ synthesized by the Agilent eArray program (Santa Clara, CA) at the Shanghai Bio Corporation (Shanghai, China). The probes were designed based on the genome sequences of H. parasuis SH0165 serotype 5 (GenBank accession numbers: CP001321). Each probe was represented twice on the array. Briefly, 2 μg RNA was reverse-transcribed into cDNA, and then transcribed into cRNA. After purification, 4 μg of the cRNA was labeled with the amino-group reactive dye, Cy3 NHS ester (GE Healthcare, Waukesha, WI), and purified.
Hybridization was performed using the Gene Expression Hybridization Kit (Agilent) according to the manufacturer's instructions. Briefly, after fragmentation, 1 μg of cRNA was hybridized to the slide by incubating at 65°C with a 10-rpm rotation for 17 hr. After sequential washings with Wash Buffers 1 and 2, the arrays were scanned by the Agilent G2565BA Microarray Scanner System with a resolution of 5 μm. The scanner used 100% and 10% post-translational modification and automatically combined the data from these two sources.
Data analysis
The signal intensities were normalized using Agilent's Feature Extraction Software and transformed into log2 values. The coefficients of two repeated spots in the same array for each gene were calculated using Microsoft Excel to estimate intra-array reproducibility. The coefficients of the two repetitions between arrays were calculated to estimate interarray reproducibility. Data were analyzed using GeneSpring 5.0 software. Genes with all signals present (flag=P) were selected for analysis. Since each gene present on the array had three corresponding probe sequences, any genes with abnormal signal intensities (≥10 or ≤5) were excluded from further analysis. The signal intensities of the remaining genes were averaged to obtain the unique signal intensity for each gene. Since a 1.5-fold change in the transcription level has been indicated as biologically significant by previous studies,15,24 the genes with a fold-change of ≥1.5 were selected for analysis. The data from this study were deposited in the NCBI Gene Expression Omnibus (GEO) (www.ncbi.nlm.nih.gov/geo/) under the accession number GSE34744.
The regulated genes were analyzed by searching through the Cluster of Orthologous Groups (COG) and Kyoto Encyclopedia of Genes and Genomes (KEGG) using the NCBI gene (www.ncbi.nlm.nih.gov/gene/) and the KEGG pathway (www.kegg.jp/kegg/pathway.html) programs, respectively.
Real-time reverse transcription–polymerase chain reaction
Reverse transcription was performed with random primers using the ReverTra Ace qPCR RT Kit (Toyobo, Osaka, Japan). The real-time reverse transcription–polymerase chain reaction (RT-PCR) gene-specific primers were designed by Primer Premier 5 software (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/mdr). RT-PCR of cDNA (1:8 dilution) was performed in triplicate with an ABI 7500 Fast Real-Time PCR System (Applied Biosystems, Foster City, CA) using reagents from a SYBR Green I PCR kit (Toyobo). The thermal cycler conditions were denaturation at 95°C for 10 sec, annealing at 60°C for 20 sec, and extension at 72°C for 20 sec. The resulting data were analyzed by the comparative Ct method for relative gene expression quantification. Specificity of the expected products was demonstrated by melting curve analysis. The fold-changes were calculated by the 2−ΔΔCt formula. 20
Results
Antibiotic concentrations suitable for transcriptional profiling
Before the transcriptional profiling, the subinhibitory and inhibitory concentrations of tilmicosin were determined as described in the Materials and Methods section. Figure 1 shows the effects of different concentrations of tilmicosin on the growth of H. parasuis SH0165. Compared with the untreated control group, tilmicosin treatments of 0.25 μg/ml for ∼16 hr (until OD600 >0.3)28,29 produced the slightest inhibition of growth, while 8 μg/ml tilmicosin for 50 min (when OD600 >0.3) 45 produced nearly absolute inhibition of growth.

Tilmicosin treatment effects on the growth of Haemophilus parasuis SH0165. (●, untreated control; ■, 0.25 μg/ml; ▲, 1 μg/ml; □, 2 μg/ml;
Whole-genome microarray profiling of H. parasuis SH0165 genes differentially expressed in response to subinhibitory and inhibitory concentrations of tilmicosin
Exposure to the subinhibitory concentration of 0.25 μg/ml tilmicosin led to differential expression of 302 genes (vs. untreated control), among which 157 were upregulated and 145 downregulated (Supplementary Table S2). Exposure to the inhibitory concentration of 8 μg/ml led to differential expression of only 198 genes (vs. untreated control; p<0.05), among which 94 were upregulated and 104 were downregulated (Supplementary Table S3). Comparison of the differentially expressed genes between the two tilmicosin treatment groups revealed that 56 of the upregulated genes and 39 of the downregulated genes were common (Fig. 2A).
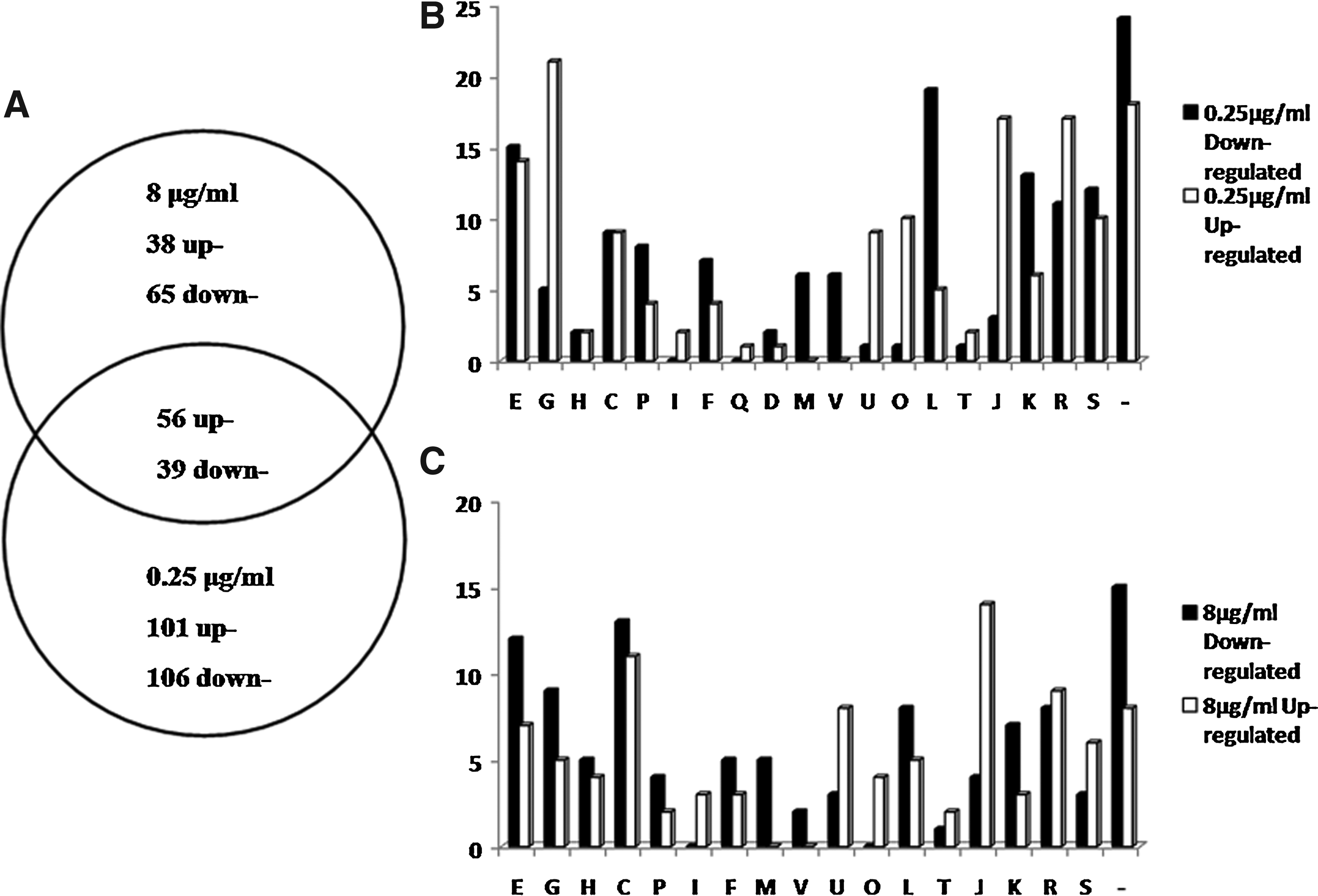
Differentially regulated genes grouped by functional classification according to H. parasuis SH0165 genome annotation.
The entire sets of differentially regulated genes in the subinhibitory and inhibitory groups were categorized by a known function, to understand the overall bacterial response to tilmicosin and gain insights into the possible mechanisms related to sensitivity (0.25 μg/ml; Fig. 2B) or resistance (8 μg/ml; Fig. 2C). Among the entire set of differentially regulated genes (n=405), the occurrence represented COG functional categories were (i) amino acid transport and metabolism, (ii) carbohydrate transport and metabolism, (iii) inorganic iron transport and metabolism, (iv) replication, recombination and repair, (v) transcription, (vi) general function, (vii) unknown function, and (viii) unrecognized genes.
Quantitative RT-PCR validation of tilmicosin-induced differential expression
Fourteen of the most differentially expressed genes between the subinhibitory and inhibitory groups and representing different COG were analyzed by quantitative RT-PCR (qRT-PCR) from the same RNA samples used in the microarray analysis. All 14 showed a similar trend in significant differential expression by qRT-PCR in both the subinhibitory (Supplementary Fig. S1A) and inhibitory (Supplementary Fig. S1B) tilmicosin treatment groups (vs. untreated control).
Tilmicosin-induced upregulation of heat shock proteins
Some of the tilmicosin-induced genes were categorized as heat shock proteins (HSPs). Induction of the major HSPs, such as chaperones, proteases, transcriptional regulators, and other structural proteins, are known to function in repair and prevention of damage caused by accumulation of unfolded proteins. 11 Previous microarray analysis of Streptococcus pneumoniae demonstrated that streptomycin induces the heat shock regulon. 34 As shown in Table 1, five heat shock genes and three chaperone proteins were significantly upregulated by the subinhibitory concentration of tilmicosin, while only three chaperone proteins were upregulated by the inhibitory concentration of tilmicosin.
Upregulated expression indicated by (+) values, and downregulated expression indicated by (−) values.
—, data not available or no change.
NF, indicates gene not found in the KEGG pathway.
Tilmicosin-induced upregulation of ribosomal protein synthesis genes
The ribosome is a large, template-directed enzyme with sophisticated proof reading capabilities, and its ligands are tRNA and mRNA. The ribosome mainly consists of a large subunit (50S) and a small subunit (30S), with the 50S ribosomal subunit catalyzing peptide bond formation during protein synthesis. Additionally, the 30S ribosomal subunit works with the 50S subunit to move the tRNAs and the associated mRNA precisely by one codon in a process referred to as translocation. 6 Treatment with the subinhibitory concentration of tilmicosin led to upregulated expression of 11 ribosomal proteins. Among these genes, rpsP (2.39-fold), rpsI (1.76-fold), and rpsE (1.55-fold) encode the 30S ribosomal protein, and rpmA (2.05-fold), rplQ (1.88-fold), and rpmI (1.57-fold) encode the 50S ribosomal protein (0.25 μg/ml; Table 1). In contrast, treatment with the inhibitory concentration of tilmicosin led to upregulated expression of eight ribosomal proteins. Among these genes, rpsP (2.47-fold), rpsB (1.61-fold), and rpsF (1.55-fold) encode the 30S ribosomal protein, and rpmA (2.54-fold), rpIU (1.69-fold), and rplQ (1.56-fold) encode the 50S ribosomal protein (8 μg/ml; Table 1). The two tilmicosin-treated groups share six ribosomal-related genes, including the rimM, trmD, and tsf genes that were upregulated.
Tilmicosin-induced downregulation of protein synthesis-associated genes
As shown in Table 1, KEGG pathway analysis identified several of the differentially expressed genes related to the mechanisms of mismatch repair and homologous recombination, and RNA polymerase; all of which were downregulated in a response to tilmicosin treatments. For example, the subinhibitory and inhibitory concentrations led to 2.5-fold and 4.9-fold downregulation, respectively, of the rarA gene of mismatch repair, while the oxidative damage repair protein, mutT/L/S, was significantly more downregulated than the subinhibitory concentration (vs. 8 μg/ml tilmicosin treatment group). Seven genes involved in homologous recombination were downregulated. Among these genes, the exodeoxyribonuclease V (RecB/C/D) gene and the DNA replication and repair RecF gene were uniquely downregulated by the subinhibitory concentration, while the ATP-dependent DNA helicase rep gene and the primosome assembly protein (priA) gene were uniquely and significantly downregulated by the inhibitory concentration. Among the differentially expressed RNA polymerase genes, the inhibitory concentration uniquely and significantly downregulated rpoB (3.4-fold) and rpoC (2.0-fold) genes.
Tilmicosin-induced downregulation of cell wall biogenesis-associated genes
Exposure to subinhibitory concentration of tilmicosin led to downregulation of six cell wall biogenesis genes and two cell cycle control genes (Table 1), while the inhibitory concentration also led to downregulation of six cell wall biogenesis genes, but no cell cycle control genes (Supplementary Table S3). Both concentrations downregulated the vimF, wecG, and pbpB cell wall biogenesis genes. The cell division protein ftsA/Q and the cell cycle protein MesJ were only inhibited by the subinhibitory concentration of tilmicosin.
Tilmicosin-induced effects of genes on transport mechanisms
Tilmicosin challenge affected genes encoding ATP-binding cassette (ABC) transporters, as well as factors involved in protein export and the phosphotransferase system (PTS). Twenty-two ABC transporter system genes were differentially expressed upon treatment with the subinhibitory concentration of tilmicosin (Table 1). These genes are known to couple the transport of the amino acid, inorganic ion, and carbohydrate to ATP hydrolysis through the cell membrane. Both concentrations of tilmicosin downregulated the expression of oppB/C/F, afuB, and fbpC genes, and modulated eight protein export genes. The majority of the known general secretory pathway (SecA/D/F/Y) genes were found to be differentially expressed in response to tilmicosin treatments. Among these genes, the preprotein translocase subunits SecA and SecY were downregulated by the inhibitory concentration, while the SecDF and lepB were upregulated by both concentrations. In addition, a total of 12 PTS genes were changed in the two groups, with seven being upregulated by the subinhibitory concentration and two being upregulated by the inhibitory concentration. Meanwhile, both concentrations led to upregulation of the sucrose-specific PTS gene (ptsB;>2.5-fold) and downregulation of the PTS operon mlc gene (<2.0-fold).
Furthermore, tilmicosin upregulated two genes in the TolA-TolQ-TolR membrane-spanning transport complex, which is believed to function in antibiotic delivery to Gram-negative bacteria. 13 These genes, TolQ and TolR, were selected for and passed verification by qRT-PCR (Supplementary Figs. S1A, B). Finally, two genes encoding permeases of the drug/metabolite transporter (DMT) superfamily, HAPS_2067 and HAPS_2068, were found to be significantly upregulated (both >3.4-fold) by tilmicosin treatment. The differential expression of these cell membrane-associated transporter genes, in particular, may represent important factors relating to antibiotic resistance.
Discussion
Gene expression profiling of H. parasuis SH0165 demonstrated the response of this Gram-negative bacterium to tilmicosin and its adaptation to the antibiotic environments of subinhibitory and inhibitory concentrations. The majority of genes found to be involved in the tilmicosin-related processes encode HSPs, and proteins related to ribosomal protein biosynthesis and transport systems.
In general, the HSPs are known to play a specific role in surface stress responses of single cells, which implies the existence of a distinct intracellular mechanism regulating protein folding and degradation that may be modulated in response to tilmicosin exposure.49,47 The tilmicosin treatment also modulated the expression of two major bacterial chaperones involved in protein folding: GroE, which has been shown to provide a protective microenvironment for protein folding, and DnaK, which binds to and protects exposed regions of unfolded or partially folded protein chains.41,48 In addition, the inner membrane cochaperone, DjlA, was modulated by tilmicosin, and this gene has been previously implicated in acquired resistance to environmental stress by another Gram-negative bacteria, Legionella dumoffii. 35
Distinct differential expression profiles were obtained in our study for the subinhibitory and inhibitory concentrations of tilmicosin. Treatment with 0.25 μg/ml tilmicosin caused unique upregulation of the HSPs, htpG, htpX, and clpP. This finding may indicate a particular mechanism that is triggered in H. parasuis that promotes resistance to the antibiotic at the subinhibitory concentration. In bacteria, htpG has been characterized as a protease, whose chief role is to degrade misfolded membrane proteins. 38 Likewise, studies of clpP in Baccilus subtilis revealed an essential role for this HSP in many of the stationary-phase adaptive responses, including motility, competence, degradative enzyme synthesis, and sporulation, 31 and demonstrated the ability of this protein to resolve accumulation of denatured proteins. 17 Considering these previously defined characteristics of the tilmicosin-induced H. parasuis HSPs, it is possible that Gram-negative bacteria upregulate expression of HSPs and chaperone proteins to counter the intracellular protein damage induced by antibiotics.
In our study, tilmicosin was also found to have a significant effect on H. parasuis genes encoding the ribosomal protein 50 subunit, including upregulation of rpmA and rplQ, and rpsP, which have been previously studied and characterized in other Gram-negative bacteria. Zouine et al. reported that overexpression of the ribosomal protein rplQ (L17) in B. subtilis remarkably perturbed nucleoid morphology and segregation. 60 Other bacteria studies have shown that the ribosomal protein rpmA (L27) mediates activity of the peptidyl transferase by interacting with its active site, possibly through a mechanism that promotes correct positioning of the tRNA substrates at the catalytic site. 25 In Escherichia coli, the ribosomal protein S16 (rpsP) has been characterized as essential for bacterial viability 40 and shown to play an important role in the assembly of the 30S ribosomal subunits. 12 While the E. coli rimM protein only weakly associates with the 30S ribosomal subunits, 22 it does bind strongly to S19, and rimM strains are partially suppressed by mutations in S13 and S19. 23 Finally, the trmD protein, which renders tRNA methyltransferase temperature-sensitive both in vivo and in vitro, is the structural gene of the tRNA methyltransferase in E. coli K-129. 4 Therefore, as Dennis demonstrated in his study of the E. coli response to chloramphenicol, when Gram-negative bacteria, such as H. parasuis, experience low levels of translation inhibition, a compensatory mechanism may be triggered in which the synthesis of ribosomal proteins is initially upregulated, but as the inhibition stress increases this compensation fails to keep pace and the cells succumb to antibiotic killing. 7
Homologous recombination is a key pathway to maintain genomic integrity between meioses during ontogenic development. The RecBCD- and RecF DNA repair pathways process broken DNA molecules, along with the RecA protein, which binds to and acts upon the single-stranded DNA. 52 In our study, the RecBCD- and RecF pathways were downregulated by subinhibitory concentrations of tilmicosin, while the inhibitory concentrations led to unique downregulation of other unrelated DNA repair genes, priA, Rep, and UvrD. The priA gene encodes a protein that binds with high specificity to D loops, thereby directing the assembly of a replication fork on a nick, double-stranded, and circular DNA template carrying a D loop. 18 Therefore, just as Boubakri et al. reported, tilmicosin-induced deceases in the helicases Rep and UvrD may exert synergistic effects on replication blockage at an inverted rRNA locus. 3
The mismatch repair mechanism is another important regulator of genome stability, though precisely which nucleotide mismatches, small insertions or deletion loops introduced by DNA polymerases are repaired in the replicated DNA strand remains unknown. The mgsA (rarA) gene, which was obviously downregulated by the inhibitory concentration of tilmicosin, was previously shown to play an important role in maintaining the genomic stability of E. coli. 51 In addition, the mismatch repair genes mutT, mutL, and mutS were downregulated in H. parasuis by tilmicosin, but to a greater extent by the subinhibitory concentration. This differential response pattern suggests a possible function for these genes in limiting intrachromosomal exchanges, while the bacteria are actively attempting to overcome antibiotic stress. 39
The rpoB and rpoC genes encoding β and β subunits of RNA polymerase were obviously downregulated by the inhibitory concentration of tilmicosin. In bacteria, RNA polymerase is responsible for the synthesis of mRNA, rRNA, and tRNA, and its catalytic function is largely carried out by the β and β subunits. 57 The tilmicosin-induced decrease in rpoB and rpoC gene expression is expected to inhibit the overall protein syntheses mediated by RNA polymerase. Such a response has previously been reported by Friedman et al. in Staphylococcus aureus, wherein altered susceptibility to daptomycin was shown to occur through mutations in rpoB and rpoC. 9
The bacteria cell envelope is a target of many antibiotics, and disruption of its structure inhibits the transmembrane transport functions and impairs normal physiological functions. The two key transport systems critical for bacterial viability and survival are the general secretory (Sec) pathway and the ABC transporter pathway. 53 In our study, the inhibitory concentration of tilmicosin induced a robust effect on the KEGG pathway protein export genes, including those involved in intracellular trafficking, secretion, and vesicular transport. Among these, the preprotein translocase subunit SecD/F genes have been proposed to be important for the release of translocated proteins from the translocase, 27 and for the prevention of backward sliding of the translocated proteins. 8 In addition, the lepB signal peptide has been demonstrated as involved in targeting preproteins for translocation via the Sec system. 46 Meanwhile, the subinhibitory concentration of tilmicosin affected the composition of large ABC transporters, and possibly regulated amino acid transport and metabolism, inorganic ion transport and metabolism, and carbohydrate transport and metabolism (Table 1). ABC transporters are involved in the secretion of the antibiotic through the cell membrane and also contribute to acquired antibiotic resistance. 30 In fact, ABC transporters were the first proteins to be implicated in the mechanism of resistance to macrolides, as described in antibiotic-producing actinomycetes. 5
Essential components of the bacterial cell wall include glycosyltransferases (GTs) and transpeptidases (TPs; also known as penicillin-binding proteins (PBPs)), which are considered excellent drug targets due to their accessibility from the periplasm and vital role in bacterial physiology and structural integrity. 21 Tilmicosin treatment of H. parasuis induced downregulation of the TP enzyme pbpB gene. It is known that the final stage of peptidoglycan synthesis depends on the high molecular weight PBPs, which polymerize disaccharide precursors into glycan chains, and then covalently crosslink the chains to form mature peptidoglycans.16,26 The cell wall biogenesis genes vimF and wcfQ encode GTs and were found to be downregulated by the subinhibitory concentration of tilmicosin. Hernandez et al. previously reported that the action of macrolide GTs in Streptomyces leads to a glycosylated inactive oleandomycin, 14 while Vanterpool et al. showed that the vimF gene itself may be involved in post-translational regulation of gingipain expression, subsequently perturbing the necessary degradation of cytokines. 54
Furthermore, several PTS system genes, such as the manX/Y/Z genes and ptsEIIB/C/D genes, were upregulated by the subinhibitory concentration of tilmicosin. The PTS system is responsible for the transport of a variety of carbohydrates in prokaryotes. PTS components participate in signal transduction, chemotaxis, and the regulation of essential physiological processes, such as carbohydrate utilization, catabolite repression, and coordination of carbon and nitrogen metabolism.61,43 However, the mlc PTS-related gene was downregulated remarkably in H. parasuis by both subinhibitory and inhibitory concentrations of tilmicosin, suggesting that the mlc gene may encode a yet unrecognized global regulator of carbohydrate metabolism, or generic PTS that acts as a nonspecific scavenger of sugars derived from cell wall breakdown.42,44
Collectively, the data obtained from our transcriptional profiling of H. parasuis SH0165 treated with tilmicosin at different concentrations and durations provide novel insights into the complex mechanisms underlying the general response to tilmicosin treatment. In addition, the distinctive differential expression pattern related to the subinhibitory concentration of tilmicosin indicates that survival under this antibiotic stress condition involves protein export, the PTS system, and ABC transporters, which would promote cell growth and survival. However, the inhibitory concentration of tilmicosin was characterized by a downregulation in genes encoding the RNA polymerase and proteins involved in recombination and repair, which would inhibit cell growth. Importantly, the specific transcriptional profiles of H. parasuis SH0165 under the various antibiotic conditions may provide promising targets for future therapeutic strategies.
Footnotes
Acknowledgments
We would like to thank Professor Hanchang Zhu for his help in revising the language of this article. The project was supported by grants from National Nature Science Foundation of China (No. 31201953), China's Agricultural Research System (No. CARS-36) and National Basic Research Program of China (973 Program, 2012CB518802).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.