
Abstract
Staphylococcus aureus is a major pathogen associated to bovine mastitis and has the ability to form a slow-growing population termed the small colony variants (SCVs). From 20 samples of 5 chronic S. aureus cases, 1 SCV isolate (SCV102) was recovered simultaneously with 1 of 8 S. aureus isolates. SCV102 showed auxotrophy for thymidine and had a slow growth rate. Intracellular persistence in human mammary epithelial cells (HBL100cell line) monolayer revealed that SCV102 isolate had minimal cytopathological effects compared with its parent strains. SCV102 isolate and its parent strain S. aureus 101 indicate similar resistant pattern to four antibiotics. On the contrary, the minimal inhibitory concentrations values for chloramphenicol and sulfadimethoxine were much higher in SCV102 than that of S. aureus 101. To the best of our knowledge this is the first time the isolation of S. aureus SCV102 from a persistent bovine mastitis has been reported in Beijing (China). This study suggests that SCV102 isolate may be an important contributor to persistent bovine mastitis.
Introduction
SCVs are often associated with persistent or recurrent infections. They differ from the S. aureus normal-phenotype not only in their small colony sizes but also by decreased pigmentation and hemolysis. Further, SCVs can be divided by two main auxotrophism subgroups, the inability to synthesize thymidine or by deficient electron transport activity caused by their auxotrophism for hemin or menadione.4,25,27,30
An additional key property of SCVs is their increased ability to survive within nonprofessional phagocytes, such as epithelial cells, fibroblasts, osteoblasts, and endothelial cells. 27 S. aureus is not traditionally considered to be an intracellular pathogen. However, over the last decade, published evidence of SCVs' ability to survive intracellularly has increased from a few sporadic reports to, at present, many model systems that report bacterial uptake and intracellular persistence. Most of these studies reported cytopathological effects of intracellular S. aureus, but only a few of these studies addressed the underlying mechanisms of intracellular survival. SCVs may serve as reservoirs for recurrent infections in cases of chronic infection. 11 Intracellular survival is thought to be a critical feature of the ability of SCVs to cause chronic and persistent infections because the intracellular location may shield these bacteria from host defenses and limit exposure to certain antibiotics.18,25,37,38
In comparison with the S. aureus normal-phenotype strains, clinical SCVs often have higher minimal inhibitory concentrations (MICs) values to antibiotics. 25 This can be due to an electrochemical gradient, which is required for the import of positively charged molecules, such as aminoglycosides and some antibiotics, into the bacterium. 2 In addition, the slow growth of these organisms reduces the effectiveness of cell wall active antibiotics, such as β-lactam.7,13 In general, it can be stated that the antibiotic treatment of S. aureus infections becomes more and more challenging. 5
Clinical isolates are often found in mixed populations of SCV and the parental strain, and even when SCVs are found in high percentage, they are easily overgrown in overnight broth culture, presenting a challenge to antimicrobial testing. Moreover, these morphological and biochemical changes make SCVs difficult to identify by diagnostic laboratories. 40 While S. aureus SCVs have received much attention in human medicine,23,36,44 they have been underestimated and overlooked in veterinary medicine.1,33,34 That can be explained due to uncommon morphological and physiological properties of the bacterium. SCVs are characterized by a strong reduction in growth rate, an atypical colony morphology, and unusual biochemical characteristics, which causes them to be frequently undetected or misidentified by standard clinical microbiology procedures. 27
The aim of this study is to characterize obtained SCVs strains from chronic S. aureus mastitis with regard to auxotrophism, growth curves performance, intracellular persistence, and antibiotic resistance pattern and to compare the SCVs with their parent isolates.
Materials and Methods
Isolation and identification of S. aureus and SCVs from milk
Raw milk samples (∼20 mL/quarter) were aseptically collected from five Chinese Holstein dairy cows (20 samples) with a history of chronic S. aureus mastitis (two acute events of S. aureus mastitis in 12 month period) from a commercial herd in Beijing, China. Consequently, obtained samples were cultured on Trypticase Soya Agar (TSA; BBL, Becton Dickinson, Sparks, MD) supplemented with 5% sheep blood, on Columbia blood agar, and on 110 agar medium plates (Difco, Becton Dickinson). The plates were incubated at 37°C and examined at 24 hr and 48 hr. Primary identification of S. aureus was done by picking the characteristic colonies (large, creamy, and strongly hemolytic on TSA with 5% sheep blood). Potential SCVs colonies were identified by tiny, nonpigmented, nonhemolytic colonies on TSA with 5% sheep blood. All these potential S. aureus and SCVs isolates were subcultured to be identified by routine methods. 1 Isolates of potential SCVs were subcultured on TSA for ten generations to assess stability. 1 SCVs and their parent strains were initially tested for routine biochemical properties. Confirmation of identification was done by sequence analysis of the 16S rRNA gene 43 and all the isolates underwent PCR analysis for the nuc gene. 6 The identified SCV S. aureus was termed as “SCV102” and its parental strain as S. aurues101 by us in the present study. Relatedness between SCV102 and parent strain of S. aureus101 were analyzed by Multiple Loci VNTR Analysis (MLVA). Hereby, five tandem repeat loci (spa, clfA, sdr, SIRU01, and SIRU21) were used for multiplex PCR as described previously,16,28 except that the annealing temperature was 52°C. 15 For further analysis, all isolates were stored at −80°C in cryoprotection medium of Trypticase Soya Broth (TSB) supplemented with 25% volumes of glycerin.
Auxotrophism assay
Auxotrophy test of the SCVs was performed on Mueller–Hinton agar (Difco BD, Beijing/China), with standard disks impregnated with 1 μg/mL of hemin (Sigma, Shanghai/China) and disks impregnated with 15 μL of 100 μg/mL of thymidine (Sigma) and 10 μg/mL of menadione sodium bisulfite (Sigma) as described previously. 18 Auxotrophism was identified if a zone of growth around the disks was detectable after incubation overnight at 37°C. 4
Bacterial growth curves
SCV102 and their parent strains were analyzed in triplicate for growth curve characteristics.1,20 Briefly, a loopful of the cryoprotected frozen stock culture was inoculated into 10 mL TSB broth and grown overnight at 37°C. One mL of overnight culture was transferred to a flask containing 200 mL TSB. Inoculated flasks were incubated at 37°C on a rotary shaker (150 rpm). Bacterial growth was determined every hour by measuring the OD600nm.
Quantitative RT-PCR
Prior to mRNA extraction, S. aureus and SCVs were grown in brain heart infusion broth medium until the stationary phase. The mRNA was extracted from the bacteria with an RNeasy mini-kit (Qiagen, China) as described in the manufacturer's protocol. Then, cDNA was synthesized from mRNA using a quantitect reverse transcription (RT) kit (Transgen, China) by following the manufacturer's recommendations.
Real-time amplification was performed using specific primers for thyA, spa, and hla genes as described previously. 10 Each reaction mixture contained 10 TaqMix(Tianjin, China)+1μl from each primer+7μl dd.water and 25 ng of cDNA as the template, and was carried out with an iCycler iQ real-time PCR system (AB Applied Biosystems 7500) using iQ SYBR green Supermix (TianGen). The levels of mRNA expression of the different genes were normalized using the expression of the internal control gene yqiL (encoding acetyl-coenzyme A acetyltransferase), which is a housekeeping gene successfully used for quantitative RT-PCR.21,30
The levels of transcripts were expressed as increases (n-fold) relative to the values for the internal control. 8 Total of 40 cycles were run under the following conditions: cDNA denaturation at 95° for 10 min, primer annealing at 55° for 30 sec, and cDNA extension at 68° for 10 min.
The intracellular persistence assay
The assay of intracellular persistence for SCV102 and the parent strain was performed according to the method used for the phagocytosis assay for bovine aortic endothelial cells. 2 Briefly, HBL100 cell monolayers were grown in Dulbecco's modified Eagle Medium (DMEM; Debco, Shanghai/China) with 10% fetal bovine serum to confluence (2x105cells) in 24-well tissue culture plates. The numbers of washed bacteria were adjusted to be nearly equal to 50 bacteria/cell and were added to the washed monolayers. The infected monolayers were incubated for 3.5hr at 37°C in 5% CO2 to allow adhesion and phagocytosis of the bacteria. The monolayers were then washed thrice with PBS to remove nonattached bacteria; 1 mL of medium containing 10 mg of lysostaphin/mL was added to eliminate extracellular bacteria. Incubation in the presence of lysostaphin was continued for 30 min, 3hr, 9hr, 24hr, and 48hr. At these time points, the monolayers were washed thrice with DMEM to remove lysostaphin, followed by addition of 1 mL of ice sterile water to disrupt epithelial cells and to release intracellular bacteria. Serial dilutions were made in sterile water. The number of intracellular colony-forming units (CFU) was determined by plating 100 μL aliquots on TSA supplemented with 5% sheep blood in duplicate. The detection limit was 10 CFU. The number of intracellular CFU at each time point was determined in triplicate.
Antimicrobial susceptibility testing
Antimicrobial susceptibility for SCV102 and the parent strain was determined for 11 antibiotics by identifying the MICs as described previously. 1 A standard broth microdilution method were used according to Clinical and Laboratory Standards Institute (CLSI) guidelines (CLSI, 2006). 12 The standard S. aureus strain (ATCC29213) was used as quality control (QC) strain and the MICs values were examined in triplicate. Results were read on a microplate reader after incubation for 16hr and 48hr, for the parent strain and SCV102, respectively.
Results
Isolation and identification of S. aureus101 and SCV102 from milk
Eight isolates of suspected typical S. aureus101 were recovered from 20 samples of five selected China Holstein dairy cows with a history of chronic S. aureus mastitis. Subcultured at 37°C for 16hr on TSA with 5% sheep blood, all eight isolates presented a morphological phenotype of large (∼4 mm), β-hemolytic, and creamy colonies (Fig. 1B), and bacteria were gram-positive coccus and arranged in grape-like clusters under light microscope (Fig. 2B). Catalase tests of incubates on pure TSA at 37°C for 16hr were strong positive. Positive rate of coagulase was 100% by the commercial kit (Staphylase test kit, OXOID, Beijing, China).

Colony morphological of SCV102 with pinpoint colonies on trypticase soya agar with 5% sheep blood
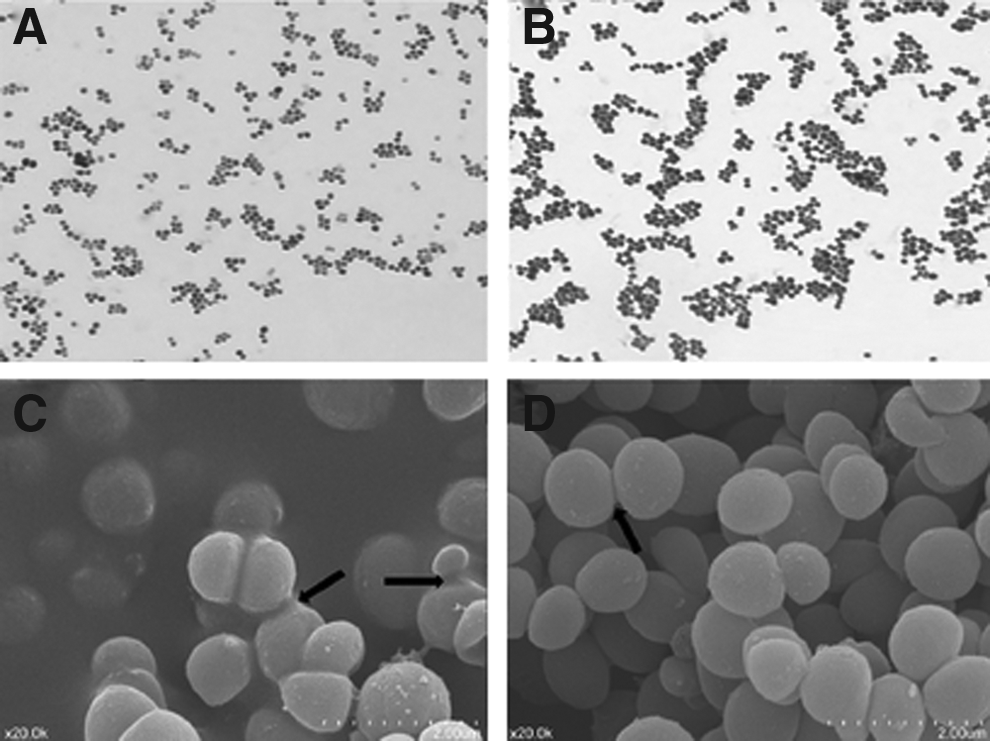
Gram stains micrographs
One putative SCV102 isolate was simultaneously recovered with one of the eight S. aureus isolates from the same raw milk sample. Colonies of SCV102were pinpoint (∼0.3 mm), nonpigmented, and failed to present an extremely slight hemolysis until 24hr of incubation at 37°C on TSA with 5% sheep blood (Fig. 1A). Under the light microscope, these potential SCV102 appeared to be identical to typical S. aureus (Fig. 2A, B). For SCV102, catalase test was also positive, while coagulase was not detected. In the course of a continuous passage for ten generations these results were stable until the 10 generation passage. Additionally, using scanning electronic microscope (×20.0 k) a minor difference in the amounts of intercellular substance was observed, whereas SCV102 showed higher amount of it than the S. aureus101(Fig. 2C, D).
All isolates were positive to nuc gene PCR test. The sequence analysis of 16S rRNA showed that gene of SCV102 and their parents are homologous with published S. aureus sequence (Accession No.: DQ997837). The MLVA typing method demonstrated that SCV102 and their parent belong to the same molecular type (Fig. 3).Analysis of the underlying auxotrophism of SCV102 revealed a thymidine dependence (Fig. 4), In contrast, the SCV102 was negative for both hemin and menadione.
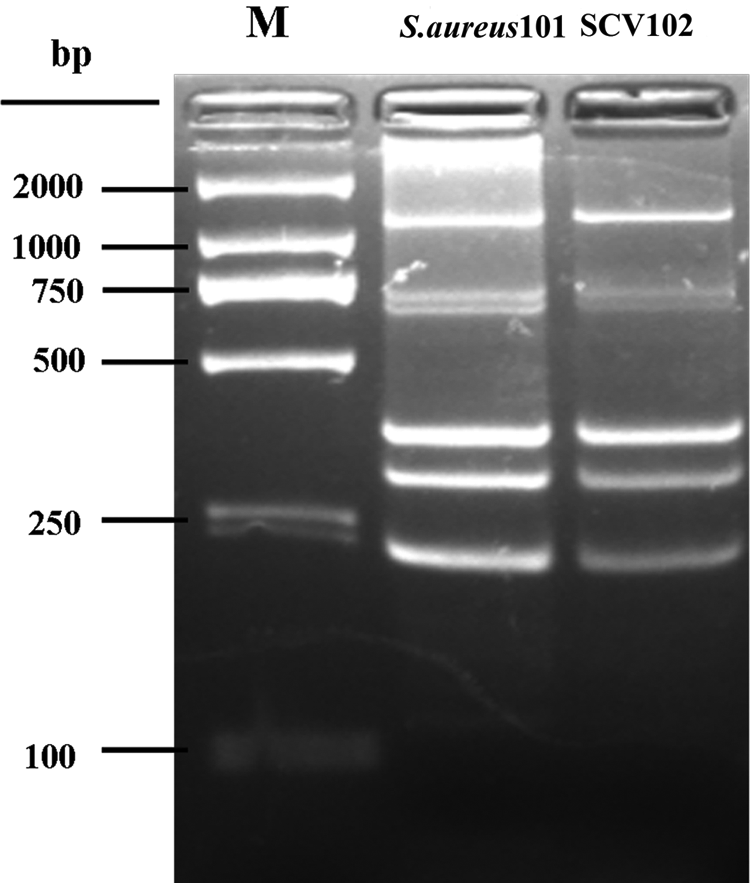
Molecular typing of S. aureus101 and its SCV102 isolated from cases of bovine mastitis by multiple loci VNTR analysis. Multiplex PCR demonstrates no variation in the five loci for the two isolates. Molecular markers: 50 and 100 bp ladder (Takara, Japan).

SCV102 growth zone on Mueller–Hinton agar with the disk impregnated with thymidine.
Bacterial growth curves
Growth curves of SCV102 and their parent strain (hereafter S. aureus101) cultured in pure TSB were drawn (Fig. 5). S. aureus101showed a typical bacterial-growth curve measured by the optical density with four phases: the lag phase (∼1 hr), the log phase (∼3 hr), followed by the stationary and decline phase. In comparison, the growth curve of SCV102 had an extended lag phase. Moreover, the division between the lag and log phase cannot be distinguished as it do not show a breakpoint like the S. aureus101. Moreover, the SCV102 do show a linear and not a logarithmic growing. Additionally, the OD600nm reached different values where it was up to 2.5 for S. aureus101 and 1 for the SCV102.
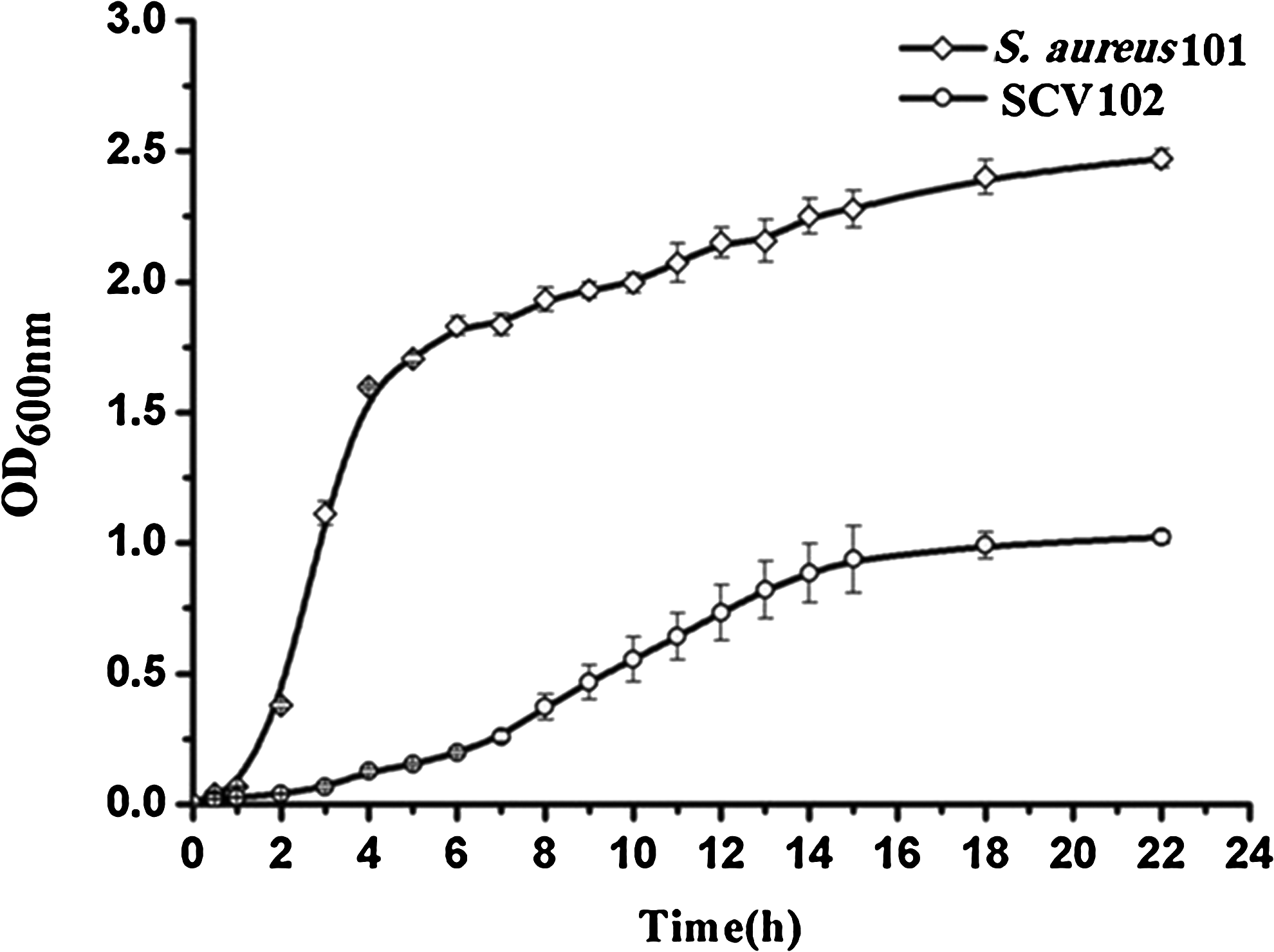
Growth curves of small colony variant SCV102 in trypticase soya broth in comparison with their parent strain.
Quantitative RT-PCR
In general, all measured mRNA of thyA, spa, and hla showed lower transcription levels in thymidine-SCV 102 (TD-SCV102) compared with S. aureus101 (Fig. 6). Whereas, the transcription level of thyA in TD-SCV102 was about 20-fold lower than the S. aureus101.
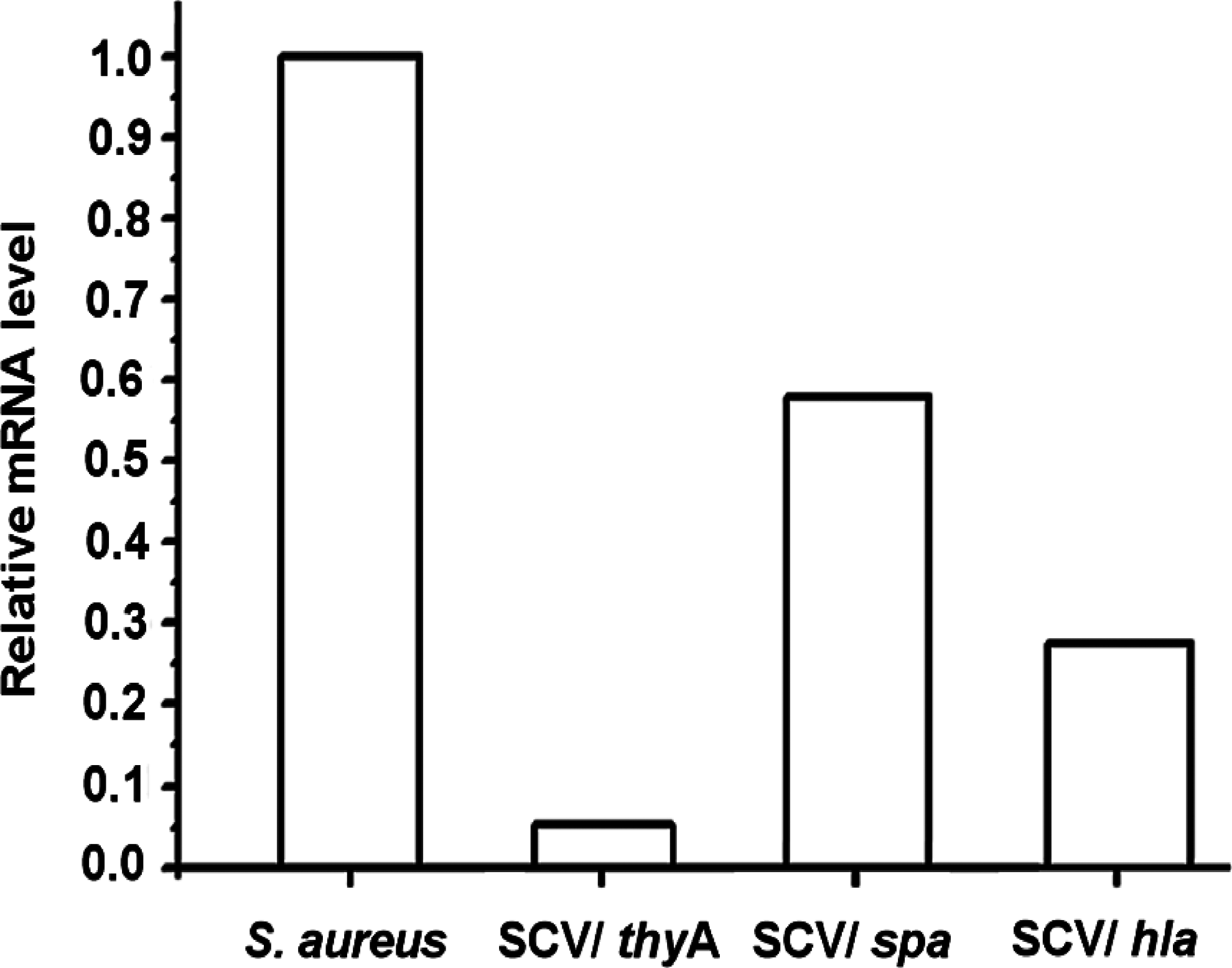
Real-time PCR quantification of expression in SCV102 of thyA, spa, and hla after overnight culture. The levels of mRNA expression of the mentioned genes were normalized using the expression of an internal control, and the transcript quantities are expressed as increases (n-fold) relative to the values for the internal control. The data are mean and standard errors of the mean obtained from three independent experiments.
Intracellular assay
Assays for SCV102 and S. aureus101 were done in triplicate to compare and quantify their ability to intracellularly persist within HBL100 cell line. Measurement of CFU was conducted at the time points 0.5 hr, 3 hr, 9 hr, 24 hr, and 48 hr. Hereby, the experimental strains showed similarity of persistent rate up to the 3 hr time point. The following four time points (9 hr, 12 hr, 24 hr, and 48 hr) show increasing differences of both strains (Fig. 7). Whereas, SCV102 showed relatively stable CFU rates compared with the S. aureus101, which drop toward the x-axis (Fig. 8).
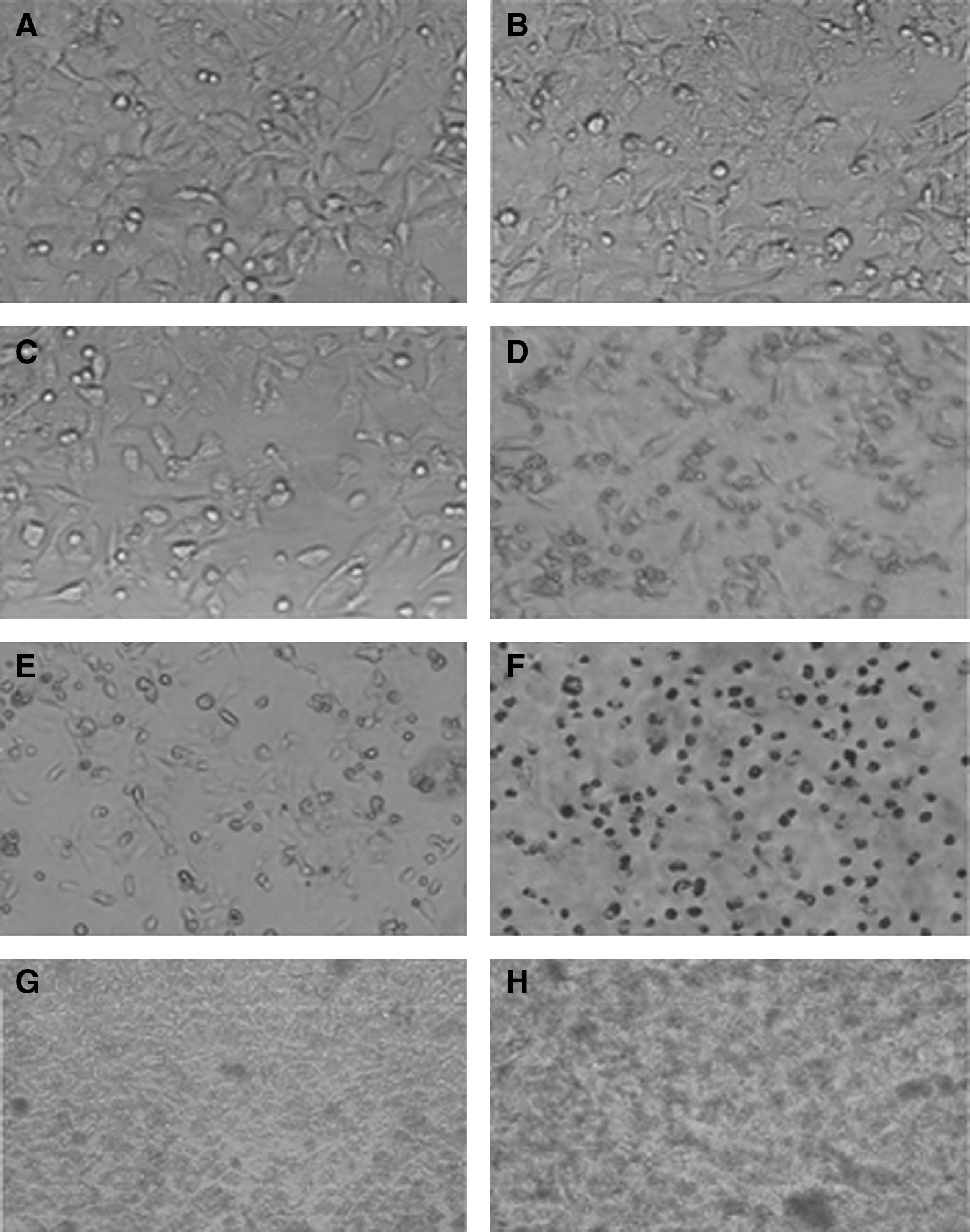
(40×) Morphological changes indicating cell damage, detachment, and rounding of HBL100 cell line infected with small colony variant SCV102, and their parent strain after 3 hours

SCV102 CFU/mL (colony-forming units/mL) recovered from HBL100 cell line at several time points compared with their parent strain.
Antimicrobial susceptibility testing
All MICs values for QC strain (ATCC29213) were within the expected ranges. Whereas the MICs values for SCV102 and S. aureus101 were much higher than the QC values for several antibiotics. Moreover, both of the experimental strains showed resistance to four antibiotics (penicillin G, tetracycline, gentamicin, and chloramphenicol) belonging to different antibiotics groups. Further, the MICs values for chloramphenicol and sulfadimethoxine were much higher in SCV102 than that of S. aureus101 (Table 1).
Median minimal inhibitory concentration (MIC) of three tests.
Breakpoints were based on sulfisoxazole.
According to Clinical and Laboratory Standards Institute (CLSI).
QC, quality control.
Discussion
The ability of S. aureus to exist as SCVs could play a part in the difficulty in eliminating S. aureus mastitis, which accounts for up to 30% of all cases of mastitis that continues to be one of the most economically important disease to the dairy industry. 32 It has to be taken into account that heterogeneity among S. aureus SCVs18,19,24,26,33,38 makes it difficult to compare results from studies that use different strains.
SCVs have been well documented and studied in humans with chronic and recurrent infections like cystic fibrosis.24,29,41 In veterinary medicine the association of SCVs with chronic and persistence mastitis are less documented. However, early studies reported a high occurrence of SCVs as the causative agent of bovine mastitis in Israel.33,34 Recently, the occurrence and association of SCVs with chronic mastitis was reported. 1 This finding is in agreement with our results, as the occurrence of SCVs during chronic mastitis at a dairy farm in Beijing is described here. While our isolate (SCV102 and S. aureus101) strains show very similar genetic phylogeny by MLVA pattern, they showed different phenotype characterization with regard to growth curves. Hereby, SCV showed extended, adaptable, and log phase of its growth curve performance and that is in accordance with previous studies.1,10 This slow growth and small phenotype is the consequence of metabolic defects. However, the investigated SCV strain in this study showed auxotrophy for thymidine. Unlike a previous study, it was found that SCV isolate from mastitis in Israel showed thiamine-auxotrophy. 33 In accordance to that difference, previous study reported that SCV isolate from persistent mastitis in Canada was identified as menadione and hemin auxotrophy, 1 indicating that all known subtypes of SCVs can be found in mastitis.
Both of the experimental strains showed resistance to four antibiotics (penicillin G, tetracycline, gentamicin, and chloramphenicol). This resistant pattern of S. aureus toward penicillin G and tetracycline is in accordance with previous study 15 in the same geographical region of Beijing. Moreover, SCVs are recognized as having higher MICs toward aminoglycosides and β-lactamase.25,42,45
SCV102 showed a high resistance pattern toward trimethoprim–sulfamethoxazole, and this can be related to thymidine-auxotrophic S. aureus SCVs, which normally interferes with the tetrahydrofolic acid pathway. Thymidylate synthase (thyA) requires tetrahydrofolic acid as a cofactor to catalyze the last step of the thymidine biosynthesis pathway, which is responsible for the formation of dTMP. Thus, mutations in thyA provide a possible explanation for thymidine auxotrophy and thereby resistance to the above mentioned antibiotics.4,9,10
The presented results indicate that SCV102 strain has the ability to intracellularly survive for a prolonged time compared with the parent strain (Fig. 8). These results of the intracellular persistence assay are confirmed by previous studies.2,29,39
Recovery of viable SCVs from endothelial cell lysates are reported to be inoculum and time dependent. 1 The ability of SCVs to persist longer without profound damage to monolayer cells may be explained by the production of small quantities of α-toxin 24 that are not sufficient to enhance apoptosis together with lack of essential nutrients such as thymidine. The persistence of SCVs within the cells may explain why an antibody-mediated immune response (AMIR) alone is not always effective and emphasizes the necessity of combined cell mediated immune response and AMIR against persistent strains of this pathogen.17,35
To the best of our knowledge this is the first time the isolation of S. aureus SCVs from persistent bovine mastitis in Beijing, China is reported. This suggests that SCVs strains may be important contributors to persistent bovine mastitis. Also, it demonstrates that such bovine SCVs strains possess different characteristic attributes than the parent strain with regard to antibiotics resistance patterns, which indicates that for successful antibiotic treatment of chronic mastitis cases the resistance patterns of SCVs has to be taken into consideration. There is little understanding of the pathophysiology of SCVs and its clinical importance in veterinary medicine, particularly in persistent bovine mastitis. So, further studies to better understand the role of SCVs in the pathogenesis of S. aureus mastitis are warranted.
Footnotes
Acknowledgments
This research was supported by the Chinese Twelfth “Five-Year” National Science and Technology Support Project (No. 2012BAD12B03) and National Education Ministry's Major Project (No. 313054).
Disclosure Statement
The authors declare that they have no conflict of interest.