
Abstract
Although enrofloxacin (ENR) is widely used for therapy of bacterial infections in the veterinary clinical, bacterial resistance to ENR is becoming an increasing worldwide problem. The primary global response of Escherichia coli to ENR exposure before resistance is largely unknown on the proteomic level. The purpose of this study was to understand the physiological response of E. coli to a subinhibitory concentration of ENR using proteomic methods. Differentially expressed proteins of the whole-cell extracts were visualized by two-dimensional gel electrophoresis, and the selected proteins were purified and identified by MALDI-TOF/mass spectrometry analysis. The result showed that the number of proteins (mean±standard deviation) detected in the ENR-treated strains was significantly (p<0.05) reduced from 1115±25 to 732±19. In total, 42 differentially expressed proteins with more than twofold difference were identified, including 13 upregulated proteins (p<0.05) and 17 downregulated proteins (p<0.05), as well as the specific proteins expressed in the group with or without ENR-treated cells. The results show that the differentially expressed proteins identified in E. coli exposed to ENR included proteins involved with a classic resistance mechanism, such as bacterial cell membrane permeability mediated by OmpX and OmpW, and other adaptive changes that appear to represent the physiological basis and background of resistance to ENR.
Introduction
In E. coli, DNA damage induces the SOS response for DNA repair, which allows a higher mutation rate of gyrA, gyrB, parC, and parE coding the target enzymes, particularly in their quinolone resistance-determining regions (QRDRs), and induction of the SOS response after exposure to quinolones is also responsible for extensive bacterial cell filamentation. 30 The profound morphological alterations in the presence of ENR and the effect of ENR on cell growth were described in our previous study. 2 Most studies on fluoroquinolone resistance mechanisms focus on the mutations of the topoisomerase genes, the expression of efflux pumps, and decreased permeability of the outer membrane proteins.24,28,38 In our previous studies, we demonstrated that––regardless of the ENR concentration that bacteria were exposed to––their efflux pumps were triggered, that the expression of the SOS regulatory genes recA and umuC and the ENR target gene parC were significantly upregulated, and that the expression of the porin gene ompW was significantly downregulated. No mutations were detected in the QRDRs of E. coli treated with the subinhibitory concentration of ENR. 1 Further microarray analysis of E. coli ATCC 25922 exposed to a subinhibitory concentration of ENR for 5 hours showed that the expression of 519 genes was altered by the treatment, of which 239 were up- and 280 were downregulated. The functions of these differentially expressed genes included the SOS response, cell division, the stress response, biosynthesis, metabolism, transport, and transcription regulation. 2 However, on the protein expression profile, only limited information was available concerning the early global physiological response of susceptible bacteria to external drug stresses such as ENR.
In the present study, 2-D gel-based proteomic methods were used to identify the altered proteins of the whole-E. coli-cell lysates after ENR treatment, to explore the global physiological changes and the early protein expression profiles of E. coli exposed to ENR.
Materials and Methods
Bacterial strains, culture, and antibiotics
E. coli ATCC 25922 was purchased from the Institute of Microbiology, Chinese Academy of Sciences. Bacteria were routinely grown in a Luria-Bertani (LB) broth at 37°C. To measure bacterial growth, 1 ml of an overnight culture in LB was inoculated in 100 ml of LB, with or without a supplement of 1/2 minimum inhibitory concentration (MIC) of ENR, and the cultures were incubated at 37°C with shaking (160 rpm). Cultures treated with ENR were considered the experimental group, and cultures without treatment were used as the control. ENR was purchased from a commercial source (Amresco, Inc.) and of 99.5% purity.
Preparation of the total proteins of E. coli
Total proteins of ENR treated and untreated E. coli cells were extracted as described previously with a few modifications. 19 Briefly, the bacterial cells were harvested at the late exponential growth phase (5 hours of growth) by centrifugation at 6,000 g for 10 minutes at 4°C. The supernatant was discarded, and the cells were washed three times with ice-cold 40 mM Tris–HCl (pH 8.0) buffer and centrifuged at 6,000 g for 10 minutes each time. Then, the pellets were resuspended in 600 μl 40 mM Tris–HCl (pH 8.0) buffer containing 1 mM phenylmethylsulfonyl fluoride and 65 mM dithiothreitol (DTT). The suspension was mixed well and disrupted by ultrasonication in 3–5-second bursts for a total of 5 minutes with a cell disrupter (MODEL: VC-750; Ultrasonics) on an ice bath. The benzonase nuclease (Merck) at a final concentration of 1 mM was added to degrade nucleic acids at room temperature for 1 hour. The supernatant was lysed with lysis buffer [8 M urea, 40 mM Tris, 4% 3-[(3-Cholamidopropyl) dimethylammonio] propane sulfonate (CHAPS), pH 8.5] at a volume ratio of 1:4, mixed well, and sonicated with the ultrasound probe as above for 5 minutes again for a better dissolution. The suspension was collected and centrifuged at 35,000 g for 15 minutes at 4°C. The supernatants were filtered through a 0.45-μm membrane to remove residual bacteria or debris, divided into single-use aliquots, and stored at −70°C. The concentration of the total proteins in the final preparation was determined by a microplate modification of the Bradford assay, 8 and a calibration curve was made with 0.5 mg/ml of bovine serum albumin as standard. Three replicates of parallel preparation of total proteins for each sample were performed in our study.
2-DE proteomics and image analysis
The proteins were resuspended in an immobilized pH-gradient (IPG) rehydration buffer (8 M urea, 4% CHAPS, 50 mM DTT, 0.2% Bio-Lyte 4/7 ampholyte, 0.001% bromophenol blue; GE healthcare) and centrifuged at 10,000 g for 15 minutes at room temperature to remove undissolved materials. Samples equivalent to 200 μg total proteins were adsorbed onto an 18-cm Ready Strip IPG strips (linear, pI 4–7; Bio-Rad), and isoelectric focusing (IEF) was performed in the Bio-Rad Protein IEF Cell according to the manufacturer's instructions. The IPG strips were first rehydrated passively at 20°C for 8 hours and were then automatically focused using the following parameters: 50 V, 4 hours; 500 V, 1 hour; 1,000 V, 1 hour; 8,000 V, linear, 1 hour; 8,000 V, rapid; and 72,000 V-hour. The isoelectric focused strips were incubated for 15 minutes in an equilibration buffer (6 M urea, 30% glycerol, 2% sodium dodecyl sulfate [SDS], and 0.375 M Tris, pH 8.8) containing 1% DTT and then incubated for 15 minutes in an equilibration buffer containing 2.5% iodoacetamide. The equilibrated IPG strips and molecular weight standards (14–97 kDa; Bio-Rad) were subjected to 12% SDS–polyacrylamide gel electrophoresis at 80 V for 45 minutes and then 200 V until the dye front reached the bottom of the gels. Three replicate gels for each sample were performed in our study.
The gels were stained by the modified silver staining method compatible with MS 39 and scanned at a resolution of 500 dots/inch, using the Uniscan D3000 scanner (Tsinghua). Spot detection, spot matching, and quantitative intensity analysis were performed using PDQuest 2-D analysis software (Bio-Rad). Altered spots were compared based on their volume percentages in the total spot volume over the whole gel image. Significantly changed spots were selected if their intensity had changed (increased/decreased) by at least twofold. 15
Identification of differentially expressed proteins by MALDI-TOF/MS and database searches
For MALDI-TOF/MS analysis, the selected protein spots were manually excised from the silver-stained gels and transferred into V-bottom 96-well microplates containing 100 μl of 25 mM NH4HCO3 (pH 8.0) in 50% acetonitrile (ACN) per well. After destaining for 1 hour, gel plugs were dehydrated with 100 μl of 100% ACN for 20 minutes and then thoroughly dried in a SpeedVac concentrator (Thermo Savant) for 30 minutes. The dried gel particles were rehydrated at 4°C for 45 minutes with 2 μl per well 0.1 μg/μl sequencing-grade-modified trypsin (Promega) in 25 mM NH4HCO3 (pH 8.0), and then incubated at 37°C for 12 hours.
The in-gel-digested peptide fragments were extracted from the gel segments using 100 μl of 5% trifluoroacetic acid (TFA) in 50% ACN aqueous mixture, followed by vortex mixing for 1 hour. After three repeated washing, the solute materials, including the peptide fragments, were dried by vacuum centrifugation. A zip-tip column (Millipore), in which C18 resin was fixed at the tip end, was used to eliminate the sample impurities. The peptide solution was prepared using an equal volume of saturated R-cyano-4-hydroxycinnamic acid solution in 50% ACN and 0.1% TFA on a sample plate of the MALDI-TOF/MS. The MS analysis was obtained on an Ultraflex MALDI-TOF (Bruker). The annotation of peptide mass fingerprinting (PMF) was performed by the MASCOT search engine (Matrix Science). The search was done against the E. coli database. 36 The parameters used were Taxonomy: Escherichia coli; Type of search: Peptide Mass Fingerprint; Enzyme: Trypsin; Fixed modifications: Carbamidomethyl(C); Variable modifications: Gln->pyro-Glu (N-term Q), Oxidation (M); Mass values: Monoisotopic; Protein Mass: unrestricted; Peptide Mass Tolerance:±100 ppm; Peptide Charge State: 1+; Max Missed Cleavages: 1. The identification of a protein with respect to theoretical parameters (pI, molecular mass, etc.) was accepted if the peptide mass matched within a mass tolerance of 100 ppm.
Bioinformatic analysis for subcellular location and statistical analysis
The subcellular location of proteins identified by MALDI-TOF/MS was determined by the Program PSORTb Version 2.0 (http://psort.org/psortb/).
Two-tailed Student's test was used for statistically analyzing the data extracted from the comparison window of Image Master Software that normalized volumes for each protein spot. All the statistical analyses were carried out with Statistical Package for the Social Sciences 13.0 for Windows software. p-Value<0.05 was considered a significant difference, and p-value<0.01 was considered as very significant.
Results
2-DE profiles of the total proteins of E. coli cultures treated with ENR
We analyzed nine pairs of samples, including nine gels for the experimental group and nine gels for the control group to evaluate the differentially expressed proteins. The percentage of common spots detected from two replicate gels using the identical sample was more than 90%. The gel-to-gel reproducibility was credible, and the matching rate of the same protein spot interclass reached more than 90%. Figure 1 shows the staining spots separated by 2-DE with sharp focusing as well as wide distribution along pI 4–7. The number of proteins (mean±SD) derived from the ENR-treated strain was significantly (p<0.05) reduced from 1115±25 to 732±19, and a total of 101 proteins showed more than twofold difference in relative spot volumes when the gel carrying ENR-treated samples (Gel A) was matched with the reference gel (Gel B) carrying control samples.
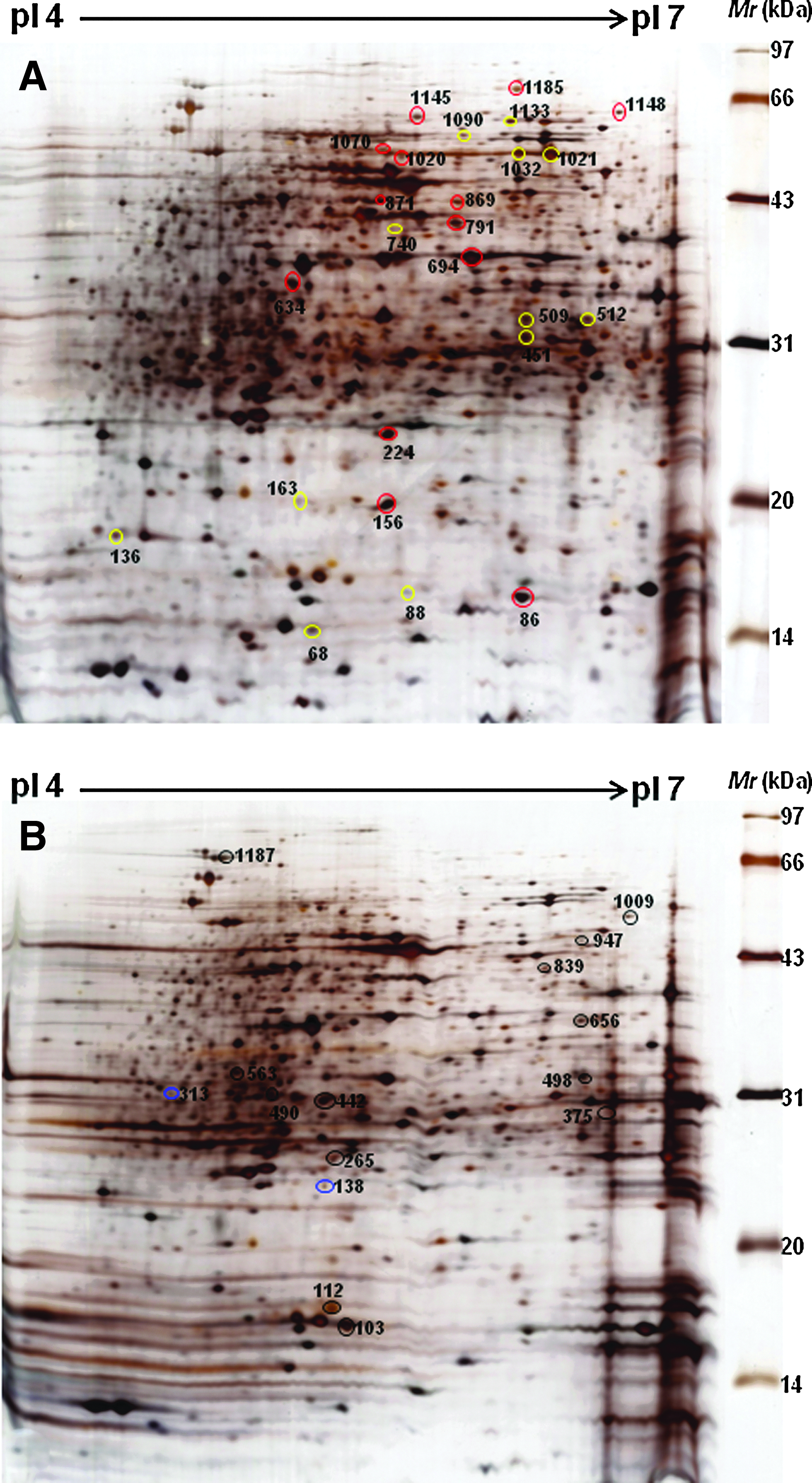
2-DE profiles of the total proteins from Escherichia coli ATCC 25922 with or without ENR treatment. Equal amounts (200 μg) of cell lysates were separated on the 17-cm pI 4–7 linear-gradient IPG strips and by 12% denaturing (SDS) PAGE.
Identification and subcellular location of differentially expressed proteins
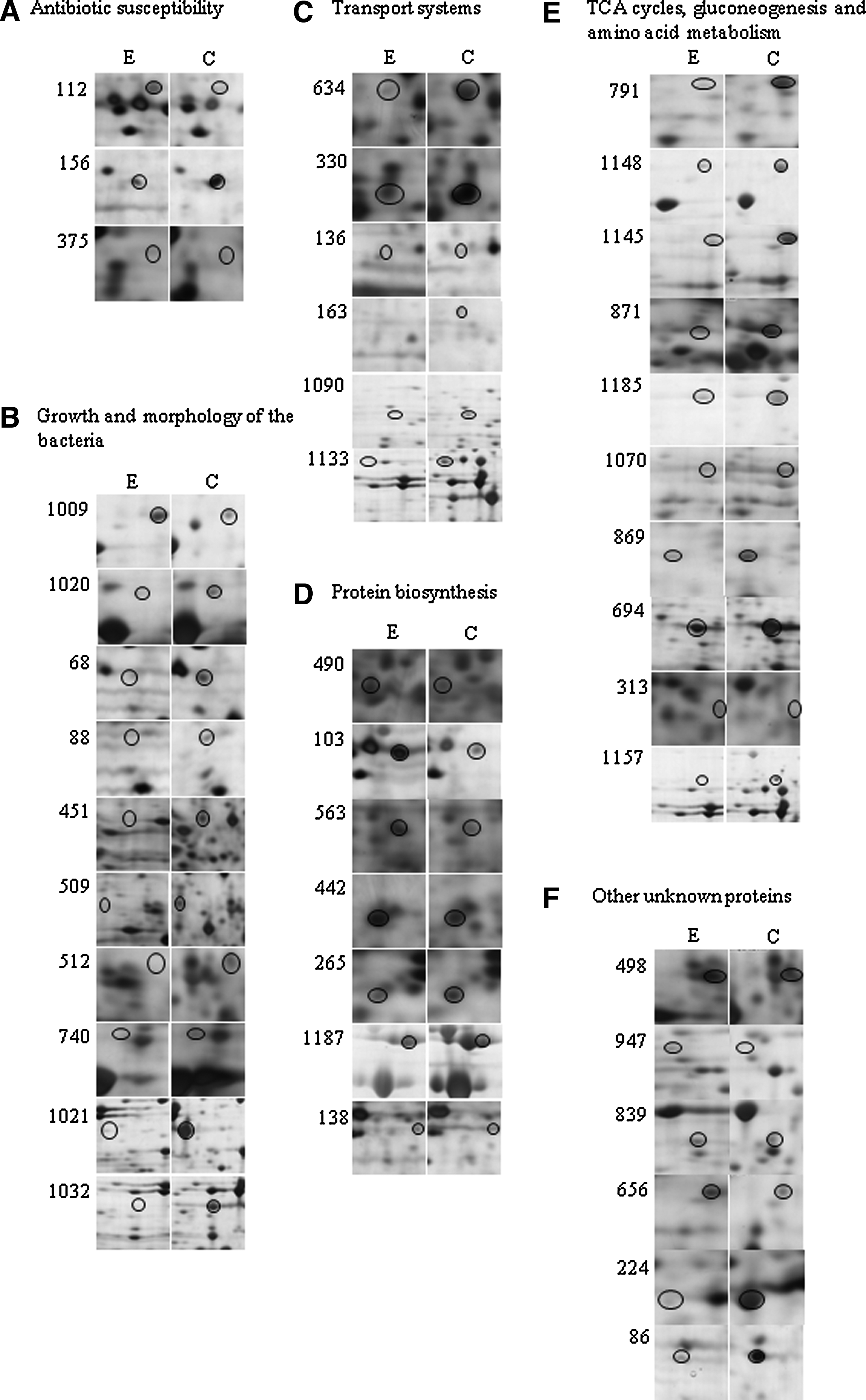
To identify the differentially expressed proteins on 2-DE images, a total of the matched 101 protein spots were excised from these 2-DE gels and subjected to in-gel trypsin digestion and subsequent MALDI-TOF identification. Of these, 42 protein spots were of reliably high quality that allowed matching with the SWISSPROT protein database. These proteins included 13 significantly upregulated protein spots (>2-fold) and 14 significantly downregulated protein spots (>2-fold), two spots specific to the ENR treatment group and 13 spots specific to the control group (Tables 1 and 2). Based on the annotations from the UniProt Knowledgebase (SwissProt/TrEMBL) and Gene Ontology Database, these proteins were involved in bacterial resistance, protein biosynthesis, amino acid metabolism, transport systems, or cell division. Figure 2 shows a comparative analysis of the differential protein expression in the ENR-treated cultures and control cultures according to their biological functions. According to the Program PSORTb Version 2.0 (http://psort.org/psortb/), 26 of the proteins were cytoplasmic; four were periplasmic, two were outer membrane proteins; one was an inner membrane protein; and nine were unknown proteins (Tables 1 and 2).
Comparative 2-DE profiles of the differentially expressed proteins in E. coli ATCC 25922 after ENR treatment. Circles indicate the differentially expressed protein spots.
A threshold of ≥2-fold was fixed, and differentially expressed protein spots were identified on two-dimensional gels. Altogether, 27 protein spots were identified, of which 13 were upregulated, and 14 were downregulated. In terms of subcellular location, two were outer membrane proteins, 17 cytoplasmic proteins, two periplasmic proteins, and six unknown proteins.
Ggbp, glucose-/galactose-binding protein.
There were two protein spots seen only in the samples from ENR treatment, and 13 were seen only in the control samples. Subcellular analysis showed that one was an inner membrane protein, nine cytoplasmic proteins, two periplasmic proteins, and three unknown proteins.
ENR, enrofloxacin.
Discussion
In our study, differentially expressed proteins were identified using 2D gel analysis and MALDI-TOF/MS. Functional analyses of the differentially expressed proteins of E. coli exposed to ENR were related to antibiotic susceptibility, growth, and morphology of the bacteria, transport system, protein biosynthesis, the tricarboxylic acid (TCA) cycles, gluconeogenesis, and amino acid metabolism.
Proteins related to antibiotic susceptibility
Adjustments in the expression levels of the different outer membrane proteins are considered to be an adaptive response to antibiotic exposure as exemplified by a recent report. 25 For example, OmpX can regulate the uptake of antibiotics such as β-lactams and fluoroquinolones because of the role in regulating membrane permeability. 14 An increased synthesis of OmpX accompanied by a reduction in the major porins was linked to multidrug resistance, including resistance to fluoroquinolones, in two clinical strains of Enterobacter aerogenes. 18 In our study, OmpX was upregulated 6.6±0.1-fold, whereas OmpW was downregulated 3.3±0.4-fold after exposure to ENR for 5 hours. The results suggest that the early regulation of OmpX may represent a preliminary response to ENR and the initial line of defense of bacteria against antibiotic therapy.
Previous studies suggested that members of the OmpW family were involved in the transport of small hydrophobic molecules across the bacterial outer membrane and in the protection of bacteria against various forms of environmental stress. 20 Downregulation of Tsx and OmpW and upregulation of OmpX are known to be required for iron homeostasis in E. coli, 24 a characteristic feature in E. coli surviving in an iron-deficient medium. Lin et al. reported that expression of the ompW gene was downregulated by exposure to chlortetracycline, and its deletion resulted in a decrease in survival capabilities. 25 The downregulated expression of OmpW in the study presented here is consistent with the downregulated gene expression of ompW, demonstrated in our previous study. 2 In contrast to these reports, OmpW was said to be upregulated in E. coli after exposure to nalidixic acid, another gyrase inhibitor like ENR. 23 It will be interesting to study the molecular interactions between ENR and OmpX and OmpW, especially in view of the changes in the size and shape of ENR-treated cells, which may influence the structure of these proteins.
The reasons why OmpF and OmpC, 31 the major outer membrane proteins in E. coli, have not been detected in our study is not clear. Possibly, OmpX and OmpW may play a dominant role in the early response to environmental stress such as ENR treatment. Moreover, the use of CHAPS, a less-denaturing zwitterionic detergent, might have been a hindrance to the solubilization of the missing major outer membrane proteins.
Another protein upregulated in the ENR-treated cells was the guanylate kinase encoded by the gmk gene is a target of an antitumor drug, 6-thioguanine, and is required for activation of some of the antiviral drugs such as acyclovir and ganciclovir.6,33 An earlier study showed that gmk was also transcriptionally regulated by heat stress during nutrient starvation and growth phase in the gram-positive bacterium, Staphylococcus epidermidis, such that the transcript levels of gmk declined after the mid-exponential phase. 35 In the present study, sampling was done in the late-exponential phase when the gmk transcriptional activity would be expected to be low, but we found a higher level (4.3±0.3-fold upregulated) of protein expression in the presence of ENR. It will be interesting to investigate if this enzyme has a role to play in the mode of action of this fluoroquinolone drug.
Proteins related to growth and morphology of the bacteria
Primase is one of the essential enzymes of chromosomal and plasmid DNA replication that initiates leading-strand synthesis once and lagging-strand synthesis multiple times during the course of replication. 7 Primase-catalyzed reactions are known to be inhibited under conditions of starvation or stress, which leads to an arrest of DNA replication and retardation in growth rate. 26 MinE is a cell division topological specificity factor in gram-negative and gram-positive bacteria. Disruption of the minE gene in Sinorhizobium meliloti resulted in large, swollen, and branched cells that could not enter into a symbiotic association with the host plant. 10 In our study, spots corresponding to primase and MinE were found only in untreated control culture samples.
Yet, another protein induced by ENR treatment was the bifunctional N-acetylglucosamine-1-phosphate-uridyltransferase/glucosamine-1-phosphate acetyltransferase, the product of the glmU gene. This enzyme catalyzes a two-step reaction yielding UDP-N-acetylglucosamine, an important precursor in cell wall (peptidoglycan) biosynthesis, and hence is considered an excellent drug target. 29 In a recent long-term evolution study in populations of E. coli, a single base change in the glmUS operon has led to reduced expression of these genes and conferred an improved fitness on the population. 32 Hence, it is intriguing to see that the amount of this protein increased 3.7±0.3-fold after ENR treatment, although the growth rate was reduced. Whether this effect is an adaptive response to ENR or is related to the dramatic changes in the cell morphology because of the role of this enzyme in cell wall synthesis remains to be investigated.
Tryptophanase, encoded by the tnaA gene, degrades
Proteins related to transport systems
NikA is a periplasmic binding protein involved in nickel uptake and transport in E. coli. It plays a dual role as the primary binding protein for Ni in the uptake process and also in negative chemotaxis in conditions of excess of Ni. 16 The oligopeptide-binding protein, OppA, binds to oligopeptide substrates and facilitates their uptake via the membrane-associated oligopeptide permease (Opp). 3 Here we found up to three- to fivefold reduction in the expression of proteins involved in active transport, for example, the NikA and OppA discussed above, as well as the glucose-/galactose-binding protein and molybdate transporter periplasmic protein, in extracts from cells treated with ENR. It is believed that importing of amino acid and other carbon sources via periplasmic binding proteins is essential for E. coli cells during energy crisis. 37
Proteins related to protein biosynthesis
Alterations in the expression of proteins related to protein biosynthesis, in a particular elongation factor complex EF-Tu, EF-Ts, and 30S ribosomal protein, were also demonstrated in ENR-treated cells. A two- to fivefold upregulation of the elongation factor complex (protein spot nos. 490, 563, 442, and 265) was recorded (Table 1). In prokaryotes, three elongation factors are required for translation: EF-Tu, EF-Ts, and EF-G. Apart from their essential role in protein synthesis, the elongation factors also play role in stress response. For instance, EF-Tu acts as a chaperone to assist in folding of proteins damaged due to stress. 9 Other proteins such as the 30S ribosomal proteins were seen to be upregulated: S6 (spot no. 103) and S1 (spot no.1187) showed increases in expression of about fivefold and twofold, respectively, after treatment with ENR, whereas one spot, no. 138 that also corresponded to S6, was detected only in the ENR-treated cells. In some reports, the ribosomal S1 protein was shown to be downregulated under oxidative stress, while its expression was unaltered under other environmental stresses such as heat or acidic pH. 34 However, in the present study, this protein was clearly upregulated during growth in the presence of ENR. Similarly, the ribosomal protein S6 is known to be induced under various environmental stress conditions such as heat shock 12 and was upregulated 5.3±0.3-fold in ENR-treated cells.
Proteins related with the TCA cycles, gluconeogenesis, and amino acid metabolism
The introduction of ENR into the growth medium appeared to influence the carbon flux by repressing the expression of related enzymes for the TCA cycle, gluconeogenesis, and fatty acid synthesis. For example, malate dehydrogenase and succinate dehydrogenase are important enzymes in the TCA cycle and were repressed after ENR treatment (Table 1; Fig. 1). Fumarate hydratase was downregulated with ENR treatment or expressed only in the control cultures. In addition, phosphoenol pyruvate carboxykinase involved in gluconeogenesis was repressed in the ENR-treated cultures (Table 1; Fig. 1). 6-Phosphogluconate dehydrogenase involved in the hexose monophosphate shunt was detected only in the extracts from ENR-treated cultures. The phosphotransferase enzyme glycerol kinase showed downregulation with ENR treatment. Another important enzyme, asparagine synthetase A (AsnA), was downregulated 10.8±0.3-fold, which was the largest fold change seen for proteins of known functions in this study.
Other proteins
There were a number of other proteins showing significantly higher expression (between 3.1±0.3 and 5.9±0.2-fold) in the group with ENR treatment, for example, F0F1-ATP synthase subunit-alpha (a response to fluoroquinolone 11 ), queuine tRNA-ribosyl transferase, 4-hydroxy-3-methylbut-2-en-1-yl diphosphate synthase, and methionyl-tRNA formyltransferase, which have been mentioned in other studies as stress- or growth-regulated proteins.4,17 Two of the downregulated proteins, spot nos. 224 and 86, were putative proteins with unknown functions, of which the former showed the highest level of change (11.8±0.2-fold).
Presumably, some of the differentially expressed proteins may be related to the reduced growth rate or to the changes in cell morphology. Some other proteins may be specifically regulated by ENR, for example, due to its effect on DNA gyrase and DNA topoisomerase IV. We did not detect the proteins associated with the SOS response that can be induced by fluoroquinolone drugs as was demonstrated in our microarray result. 2 The reasons for this discrepancy are not clear at the present time. Further studies on individual proteins will be necessary to identify those that may be involved in development of resistance to ENR.
Conclusion
This study adopted a gel-based comparative proteomic approach to analyze and identify the differentially expressed proteins of E. coli ATCC 25922 during growth in a medium containing ENR at a concentration of 1/2 MIC (0.025 μg/ml). The proteomic analysis revealed the early protein expression profiles in E. coli on exposure to ENR and identified 42 differentially expressed proteins. An upregulated expression (p<0.05) of 13 proteins was detected, including OmpX, F0F1-ATP synthase subunit-alpha, and elongation factor complex proteins, Ef-Tu and EF-Ts. Downregulated expression (p<0.05) of 17 proteins was detected, including OmpW, AsnA, and tryptophanase. MinE and primase, which are involved in cell shape and division, were absent from the proteome of ENR-treated cells.
Our results demonstrate on the level of protein expression––the global response of E. coli to ENR exposure. The proteins involved included such classic components of resistance mechanisms as OmpX and OmpW. The other adaptive changes may represent the physiological basis and background of resistance to ENR. The data generated in this study will serve a base to launch future studies for detailed analysis of the role of each of the potentially interesting proteins in bacterial adaptive responses to and development of resistance to fluoroquinolone drugs.
Footnotes
Acknowledgments
This project was supported by the National Natural Science Foundation of China (No. 30770044). The authors would like to thank Mr. Tang Chong (Beijing Genomics Institute), Mr. Wang Guibin (Beijing Genomics Institute), and Ms. Zheng Xiaojuan (Zhejiang University) for technical assistance and experimental help.
Disclosure Statement
No competing financial interests exist.