
Abstract
Telavancin is a novel semisynthetic lipoglycopeptide derivative of vancomycin with a dual mode of action. This study sought to understand the mechanisms of decreased telavancin susceptibility in a laboratory-derived Staphlococcus aureus mutant TlvDSMED1952. There were extensive changes in the transcriptome of TlvDSMED1952 compared to the susceptible parent strain MED1951. Genes upregulated included cofactor biosynthesis genes, cell wall-related genes, fatty acid biosynthesis genes, and stress genes. Downregulated genes included lysine operon biosynthesis genes and lrgB, which are induced by telavancin in susceptible strains, agr and kdpDE genes, various cell surface protein genes, phenol-soluble modulin genes, several protease genes, and genes involved in anaerobic metabolism. The decreased susceptibility mutant had somewhat thicker cell walls and a decreased autolytic activity that may be related to decreased proteolytic peptidoglycan hydrolase processing. Membrane fatty acid changes correlated with increased membrane fluidity were observed. It seems likely that there are multiple genetic changes associated with the development of decreased telavancin susceptibility. The TlvDS mutant showed some similar features to vancomycin-intermediate S. aureus and decreased daptomycin susceptibility strains, but also exhibited its own unique features.
Introduction
One of these novel agents is telavancin, a lipoglycopeptide developed from the parent molecule vancomycin. 19 This agent exhibits potent activity against methicillin-sensitive S. aureus (MSSA), MRSA, VISA, and VRSA both in vitro and in vivo.5,10 Telavancin, marketed as ATIVIV™, has been approved by the Food and Drug Administration for the treatment of complicated skin and skin structure infections, and in Europe for treatment of nosocomial pneumonia known or suspected to be caused by MRSA. This molecule has a dual mode of action on S. aureus causing the inhibition of peptidoglycan biosynthesis and membrane depolarization.11,23,39
Although we are unaware of the development of telavancin-resistance or decreased susceptibility in clinical strains, the potential fate of a novel antimicrobial agent that is introduced into use is the development of resistance or decreased susceptibility to varying degrees and over varying timescales. For example, while frank resistance may be rare, modest increases in minimum inhibitory concentrations (MICs) have occurred to vancomycin and daptomycin resulting in vancomycin and daptomycin decreased susceptibility isolates.12,25 It is desirable then to have knowledge of the mechanisms involved in resistance or decreased susceptibility to each antimicrobial agent, especially an agent with a dual mode of action.
In a recent study, Kosowska-Shick et al. 17 reported on their attempts to select telavancin-resistant strains in MRSA. One stable mutant was obtained after 43 days in multistep resistance selection studies from one of the ten MRSA strains tested, which was also a vancomycin-susceptible strain (VSSA). In this article, we describe a variety of approaches we have taken to try to understand, for the first time, the mechanisms involved in decreased telavancin susceptibility.
Materials and Methods
Strains studied and growth conditions
The strains studied were the telavancin-susceptible parent strain MED1951 and telavancin decreased susceptibility (TlvDS) mutant MED1952 kindly provided by Dr. Peter Applebaum, which were designated as parental SA248 and passage day 43 isolate, respectively, in the original publication describing these strains. 17 The strains were grown in Mueller-Hinton broth supplemented with 25 mg of Ca2+ liter−1 and 12.5 mg of Mg2+ liter−1. The growth rates of the strains were assessed by determining the mean generation times of cultures in the exponential phase of growth. Antibiotic MICs were determined as described by Kosowska-Shick et al. 17 or by the E-test.
Transcriptional profiling
The strains were grown to an OD600 of 0.4 (50 ml of culture in a 250-ml Erlenmeyer flask) with shaking (210 rpm) at 37°C. Transcriptional profiling was carried out using S. aureus genome microarrays version 8.0 provided by the Pathogen Functional Genomics Resource Center (PFGRC) of the National Institutes of Allergy and Infectious Diseases (NIAID) as described in previous publications from this laboratory.26,39 The full genome array consists of 70-mer oligonucleotides representing 4589 ORFs from the S. aureus strain COL, Mu50, MW2, N315, MRSA252, MSSA476, USA300-FPR3757, and pLW043. Each ORF is printed three times on the array. The data discussed in this publication have been deposited in NCBI's Gene Expression Omnibus (GEO) (www.ncbi.nlm.nih.gov/geo/) and are accessible through the GEO Series accession number GSE40697.
Microarray validation by real-time reverse transcription polymerase chain reaction
This was carried out as described by Song et al. 39 The primers used are shown in Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/mdr).
Autolysis, preparation of cell walls, and peptidoglycan hydrolase profiles
Triton X-100 stimulated autolysis of whole cells grown in Ca2+- and Mg2+-supplemented MHB was determined as described by Gustafson et al. 8 Crude cell walls retaining autolytic activity (CCW) and purified cell walls (PCW), which have been boiled and digested with trypsin and do not retain autolytic activity, were prepared as described previously.16,30 An autolysin extract was prepared by suspending washed exponential phase cells in a 0.01 M K2PO4-K2HPO4 buffer (pH 7.2) and subjecting them to three freeze–thaw cycles (−80°C for 1 hr at 37°C for 10 min).16,30 The suspension was centrifuged (13,380 g, 10 min) and the supernatant was used as the freeze–thaw autolysin extract for digestion of PCW. The peptidoglycan hydrolase profiles of these extracts were determined by zymography by renaturation following sodium dodecyl sulfate–polyacrylamide gel electrophoresis in a gel containing 0.2% (wt/vol) Micrococcus luteus cells (Sigma) as previously described.16,20
Fatty acid composition
Washed exponential phase cells were saponified and methylated and fatty acid methyl esters were analyzed on an Agilent 5890 dual-tower gas chromotograph. Fatty acids were identified using the MIDI microbial identification system (Sherlock 4.5 Microbial identification system) at Microbial ID. 50
Total carotenoid determination
This was determined by measuring the OD465 of cells extracted with methanol at 55°C for 3 min. 18
Measurement of membrane fluidity
This was determined as described previously. 38 In brief, midexponential phase cells (OD600 0.6) were washed twice with 0.85% NaCl. Then, the cells were resuspended in 0.85% NaCl containing 2 μM 1,6-diphenyl-1,3,5-hexatriene (DPH; Sigma) to an OD600 of 0.3 and incubated at 30°C for 1 hr. A 1 mM DPH solution was prepared in tetrahydrofuran, and 200 μl was added to 50 ml of 0.85% NaCl. DPH fluoresces in the hydrophobic regions of the lipid bilayer, but does not fluoresce in an aqueous environment. 1 Excess tetrahydrofuran was removed by flushing with nitrogen. Fluorescence polarization was measured using a QuantaMasterTM40 spectrofluorometer (Photon Technology International, Inc.). The excitation and emission wavelength for DPH were 360 and 426 nm, respectively. The higher the value is, the lower the membrane fluidity. The experiment was performed twice, and the mean polarization values were compared for significant differences by using the t test.
Transmission electron microscopy
Preparation and examination of S. aureus cells by transmission electron microscopy were performed as described previously. 45 Thin sections stained with uranyl acetate and lead citrate were examined in a Zeiss 10-C transmission electron microscope operating at 60 kV. The cell wall thickness was determined using photographic images at 50,000×final magnification. Thirty five cells of each strain with nearly equatorial cut surfaces were measured, and results are expressed as mean values±standard deviation.
δ-Hemolysis assay for Agr dysfunction
δ-hemolysin production was measured by streaking the strains adjacent to a β-hemolysin disk (Remel) on a Tryptic Soy Agar plate with 5% sheep blood. 34 Results were observed after overnight incubation at 37°C. Agr dysfunction was evaluated by the absence of δ-hemolysis within the β-hemolysis zone due to lack of synergistic hemolysis.
Results and Discussion
Characteristics of the strains
The telavancin MICs of the strains were MED1951 0.12 μg ml−1 and TlvDSMED1952 1 μg ml−1, an eightfold increase in MIC. TlvDSMED1952 was derived through multistep resistance selection with a subinhibitory concentration of telavancin at day 43. The elevated MIC of the mutant was stable in that it remained the same after ten passages in the absence of an antibiotic. There was also a fourfold increase in vancomycin MIC going from 0.5 to 2.0 μg ml−1; a twofold increase in daptomycin MIC going from 1.0 to 2.0 μg ml−1; a 2.6-fold increase in oxacillin MIC going from 12 to 32 μg ml−1 in strains MED1951 and TlvDSMED1952, respectively. The seesaw effect of decreasing oxacillin susceptibility seen in daptomycin decreased susceptibility isolates 48 was not observed here. TlvDSMED1952 did not demonstrate heteroresistance to telavancin or vancomycin in the population analysis profile. The mutant grew at a somewhat slower rate, which was about 67% of that of the parent strain. This is in contrast to what can be found in some VISA and daptomycin-resistant strains, where growth is often markedly slower than their susceptible parent strains.12,37
Changes in gene expression in TlvDSMED1952 compared to telavancin-susceptible parent strain MED1951
Compared to the strain MED1951, 330 genes were increased in expression and 311 genes were decreased in expression using a twofold cut off in the strain TlvDSMED1952. Changes in gene expression of selected genes are shown in Table 1 and the full transcriptome is shown in Supplementary Tables S2 and S3 in the Supplementary material. Considerable number of genes in the categories energy metabolism, cellular processes, cell envelope, regulatory functions, and transport and binding proteins were upregulated (Supplementary Table S2 Supplementary material), and the distribution between these different categories is given in Supplementary Table S4.
Upregulated genes
Some of the most highly upregulated genes were as follows: SAV2569 isaA 5.3-fold; SAV2299 ssaA 9.7-fold; SACOL2291 staphyloxanthin biosynthesis protein 6.3-fold; SACOL2280 ureA 33.8-fold plus other ure operon member genes; SAV2530 putative L-serine dehydratase 15.1-fold and SAV2531 putative beta-subunit of L-serine dehydratase 12-fold; SAV0017 purA 14.3-fold; SAV2095 similar to SceD precursor 25.1-fold.
In the cell envelope category, there were 2–2.3-fold increases in the expression of murF, murC, pbp2, and llm (tarO), the first enzyme in teichoic acid biosynthesis, and drp35, a gene encoding a protein with the Ca2+-dependent lactonase activity that is upregulated by cell wall antibiotics.41,46 Two genes involved in metabolism of the peptidoglycan interpeptide bridge were upregulated: femB, which is involved in adding a glycine residue to the bridge, and lytM, a glycylglycine endopeptidase. lytM, isaA, and ssaA, which were also upregulated, are members of the WalKR regulon. The WalKR (YycGF) two-component system is essential in S. aureus, and plays a significant role in the control of the expression of several peptidoglycan hydrolase genes. 6 LytM is a glycyl-glycyl endopeptidase. SsaA and IsaA are CHAP-domain (Cysteine, Histidine-dependent Aminohydrolases/Peptidases) peptidoglycan hydrolases. Delaune et al. 4 have suggested that these enzymes play a role in relaxation of crosslinking in the peptidoglycan cross bridge, especially in the absence of WalKR. The sceD precursor gene was upregulated 25.1-fold. SceD is a peptidoglycan lytic transglycosylase. 40 However, despite upregulation of these peptidoglycan hydrolase genes, overall autolysis of TlvDSMED1952 was lower compared with MED1952, see below.
bacA, undecaprenyl pyrophosphate phosphatase; mvaK1, mevalonate kinase; mvaS, isoprenoid synthesis, ppS (undecaprenyl diphosphate synthase) were upregulated. These genes are involved in the biosynthesis of undecaprenyl phosphate, which is a critical component of lipid-I and lipid-II peptidoglycan intermediates, and lipid intermediates involved in teichoic acid biosynthesis. This may be a correlate of enhanced or altered cell wall synthesis in TlvDSMED1952. Similar changes in expression of isoprenoid biosynthesis genes have also been noted in a mutant showing decreased susceptibility to the membrane-active household disinfectant pine oil, 18 which, like the lipoglycopeptide telavancin, is membrane active.
Genes involved in the biosynthesis of several cofactors, prosthetic groups, and carriers, including pantothenate and coenzyme A, thiamine, heme, porphyrin, cobalamin, biotin, menaquinone and ubiquinone, pyridoxine, and glutathione, were upregulated. This suggests that there is a significant change in metabolism in this strain with possibly a greater dependence on the biosynthesis of these molecules.
betA and gbsA, encoding choline and glycine betaine aldehyde dehydrogenase, respectively, were upregulated. These enzymes metabolize choline to glycine betaine, 14 a compatible solute involved in osmoregulation that has beneficial effects on the protein structure. 31 In addition, there was upregulation of the heat shock protein genes clpP, htrA, and groES (about 1.9-fold). With the upregulation of genes involved in accumulation of protective solutes and proteins to deal with stress, TlvDSMED1952 may be primed to deal with the challenge posed by exposure to telavancin.
Several genes involved in the fatty acid metabolism were upregulated—acetyl-CoA acetyl transferase homolog, fabG1, fabH, accB, acetyl CoA carboxylase, and fabD. TlvDS MED 1952 shows an altered fatty acid composition from MED1951, see below.
A variety of intermediary metabolism genes were upregulated. ure member genes of the urease operon were highly upregulated. Urease degrades urea to NH3 and CO2. One possible role of NH3 production may be to neutralize any excess acidity that may be produced through changes in the metabolism of the mutant. Also, a putative L-serine dehydratase, which breaks serine down to pyruvate and ammonia, was upregulated 15.1-fold.
arg, encoding arginase, and rocA, Δ’-pyrroline-5-carboxylate dehydrogenase, showed an increased expression and are part of the arginase pathway whereby arginine can be utilized as a source of carbon and energy resulting in the production of glutamate. 42
Several genes of the purine biosynthesis operon were overexpressed. This is consistent with decreased expression of purR, the negative repressor of the pur operon.
A variety of genes involved in protein synthesis, including ribosomal protein genes, tRNA and rRNA base modification genes, and tRNA aminoacylation genes, were upregulated suggesting some changes in protein synthesis in the mutant.
Various regulatory genes were upregulated, including sarA, rot, treR (1.9), tcaR (1.7), and ccpA (1.6). SarA is involved in the control of expression of agr, 32 a quorum-sensing locus in S. aureus. Rot is a global regulator with both positive and negative effects on S. aureus gene expression, 33 and some members of the Rot regulon were changed in expression.
A range of transporter genes were upregulated. Also, genes of various hypothetical proteins were upregulated.
Downregulated genes
The distribution of downregulated genes in various functional categories is given in Supplementary Table S4. Considerable numbers of cell envelope, cellular processes, energy metabolism, regulatory function, and transport and binding proteins were downregulated.
Some of the most highly downregulated genes were as follows: SAV215 probable transmembrane protein SmpB −28.1-fold; SAR0136 sasD −17.8-fold; SAV0158 capJ −13.6-fold and other cap operon genes; SAS1940a delta-hemolysin precursor −30.5-fold; agrB −8.8-fold; SAV0990 similar to peptide-binding protein OppA −16.7-fold; SAS0202 putative pyruvate formate-lyase activating enzyme −13-fold; pflB −12.2-fold; hutU −10.3-fold; SAV2514 probable transport protein −28.9-fold; SAR0071 potassium-transporting ATPase B chain −27.7-fold; SAR0070 potassium-transporting ATPase submit A; oppC −10.3-fold; SACOL1187 phenol soluble modulin −40-fold; SACOL1186 phenol soluble modulin −29.1-fold; spa immunoglobulin G binding protein A precursor −26.1-fold; aur zinc metalloproteinase aureolysin −20.4-fold.
In the category of cell envelope genes, various genes of the capsular polysaccharide biosynthetic operon (cap) were strongly downregulated.
Genes of the lysine biosynthetic operon were downregulated, as was lrgB, an anti-holin protein (Table 1). Interestingly, these genes are overexpressed in response to the membrane-depolarizing and membrane-active effects of telavancin, daptomycin, bacitracin, and various other antimicrobial peptides.21,26,39,44 Although upregulation of these genes by the cell is viewed as a response to combat telavancin challenge, 39 perhaps their downregulation makes the mutant less responsive to telavancin in some way.
The delta hemolysin precursor gene was downexpressed 30.5-fold. Expression of delta hemolysin is a marker of the Agr system, which is a quorum-sensing system that controls the expression of virulence and other genes.13,28 agrA, agrB, agrC, and agrD were also markedly downregulated. This suggested that TlvDSMED1952 was likely to be Agr- and indeed this was confirmed by the blood agar plate assay by lack of synergistic hemolysis within the β-hemolysin zone, 34 in contrast to MED1951, which was Agr+(Fig. 1). There is also an association between the VISA phenotype and defects in Agr. 43
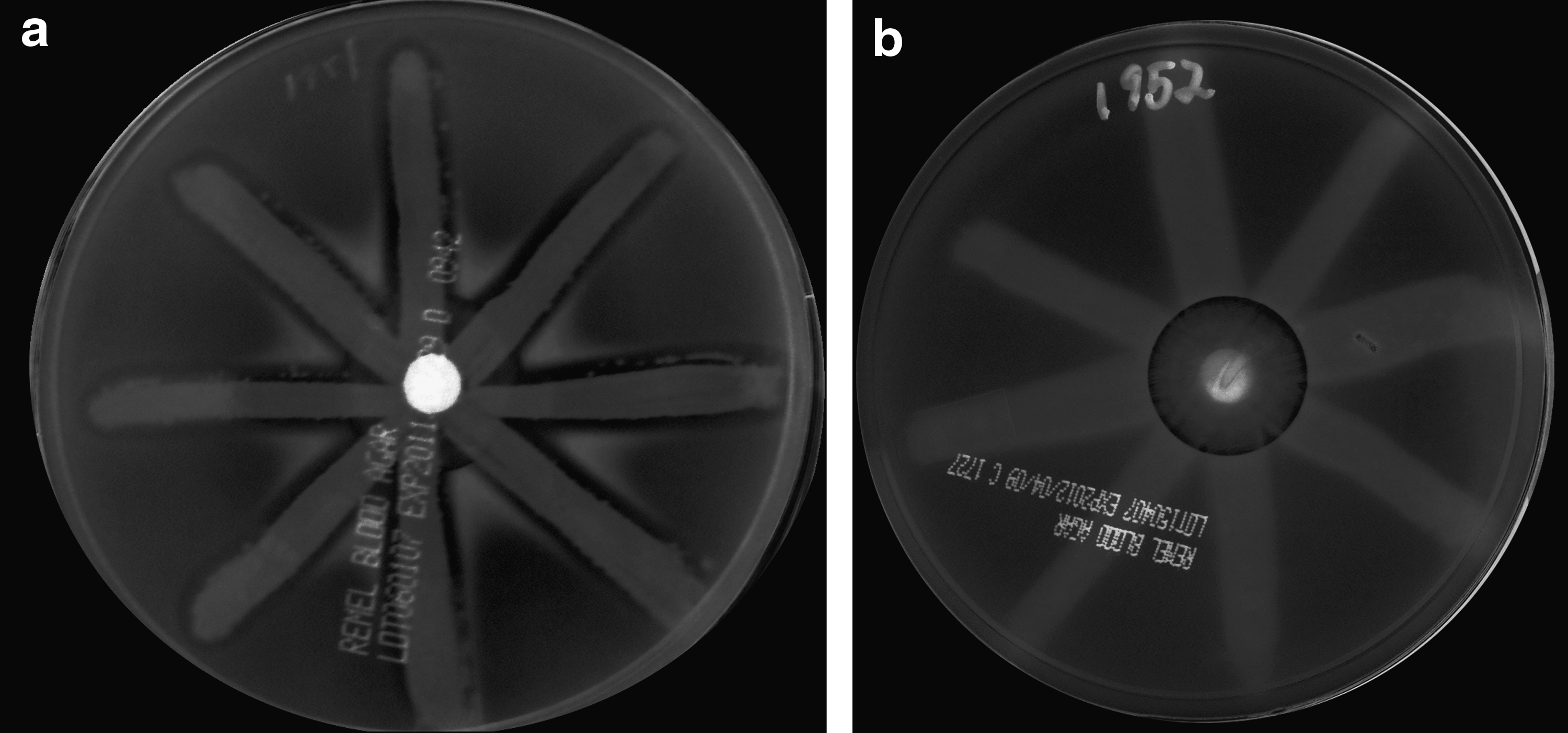
Visualization of agr dysfunction by δ-hemolysin assay.
Several protease genes were downregulated: sspB2, cysteine protease precursor; sspA, cysteine protease; sspC, cysteine protease; and aur, zinc metalloproteinase aureolysin. Peptidoglycan hydrolase profiles of TlvDSMED1952 autolysin extract showed decreased proteolytic processing of peptidoglycan hydrolases in this strain (see below), which may be related to decreased transcription of protease genes. A sspC S. aureus mutant showed a diminished autolytic activity. 35
Various cell surface-associated protein genes were under expressed: sasD, clfB, sdrE, isaB, and spa, protein A precursor. It would seem that TlvDSMED1952 would present a significantly different cell surface than its parent strain. Downregulation of spa gene expression is a common characteristic of VISA strains. 12 This may be related to possible alterations in the peptidoglycan structure in TlvDSMED1952, particularly involving the interpeptide bridge in that lytM and femB are increased in expression.
Several genes involved in anaerobic metabolism were downregulated, including nar genes involved in nitrate and nitrite metabolism, pflB, formate acetyltransferase; formate lyase-activating enzyme; and ldh1, lactate dehydrogenase. Possibly, TlvDSMED1952 has a greater dependence on aerobic metabolism than its parent strain.
kdpD and kdpE, a potassium two-component sensor and response regulator were downregulated. Xue et al. 47 have provided evidence that the main function of KdpDE is not in K+ transport via the KdpFABC system, but is in the regulation of the virulence factor expression. Genes SAR0071 and SAR0070 encoding proteins of the kdpFABC gene cluster are controlled by kdpDE. Genes regulated include spa, cap, hla, aur, and hlgB. Also, Agr/RNAIII strongly activated the transcription of kdpDE. 47 Clearly, downregulation of the agr system may well be involved in downregulation of kdpDE, and thereby, further genes affected by it. Gardete et al. 7 have noted downregulation of major virulence determinants in a clinical VISA.
oppC and oppF were strongly downregulated, as well as a gene similar to oppA. Transport of peptides may be impaired in TlvDSMED1952. Various other transport genes were also downregulated.
SACOL1187 and SACOL1186, encoding phenol soluble modulins, were highly downregulated. Phenol-soluble modulins are small peptides of about twenty amino acids that destroy neutrophils at low concentrations, 3 with a significant role in S. aureus pathogenesis.
smpB was significantly downregulated. SmpB, small protein B, is a probable transmembrane protein that binds to transfer-messenger RNA (tmRNA).9,36 Decreased transcription of smpB may lead to decreased cellular levels of SmpB and may result in altered regulation of gene expression.9,36
Verification of transcriptional profiling results by real-time reverse transcription polymerase chain reaction
The fold changes in gene expression of selected up- and downregulated genes as indicated by microarray and real-time reverse transcription polymerase chain reaction (RT-PCR) results are shown in Supplementary Table S5. RT-PCR confirmed the up- or downregulation of the selected genes, with some quantitative differences in the magnitude of gene expression in some cases.
Ultrastructure
The cell walls of TlvDSMED1952 were about 30% thicker than those of MED1951 (32.4±0.75 nm versus 25.0±3.0 nm, Fig. 2). Markedly thicker cell walls are often a characteristic of VISA and daptomycin-resistant strains.29,48 This may be related to the upregulation of some cell wall biosynthesis genes noted above. However, otherwise, the cells appeared normal, with normal division septa.
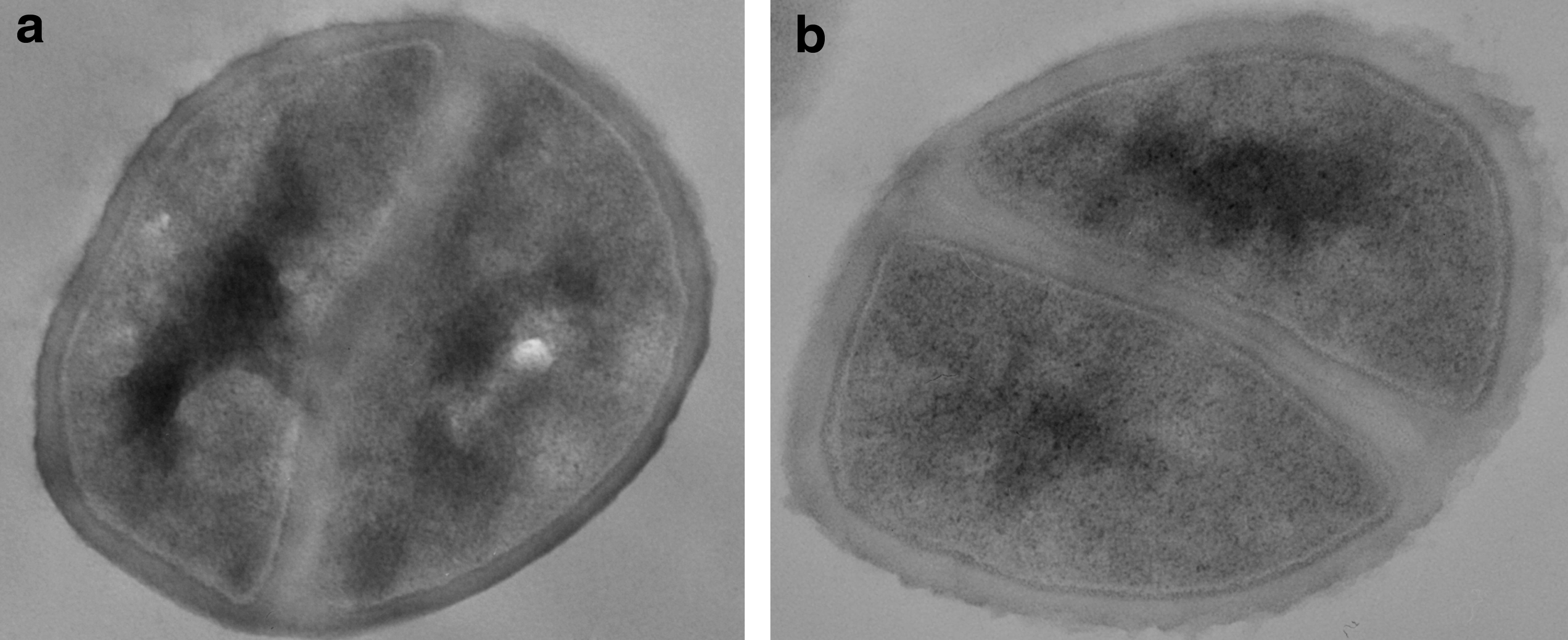
Ultrastructure of the strains.
Autolysis
It is a common characteristic of VISA and daptomycin-resistant strains that they show a decreased autolytic activity compared to their susceptible parent strains.12,16,37 TlvDSMED1952 showed a somewhat decreased Triton X-100-stimulated autolysis (Fig. 3a), although some VISA and daptomycin-resistant strains show a more severe reduction in the autolytic activity.12,16,37 In an effort to investigate decreased autolysis in TlvDSMED1952 further, we determined the autolytic activity of isolated CCW that retain autolytic activity. The CCW from the strain TlvDSMED1952 also had a lower autolytic activity than those from the strain MED1951 (Fig. 3b). This indicates that the decreased autolysis is due to either alterations in the structure of the peptidoglycan, which is the autolysin substrate, or in the autolysin amount or activity. We next examined the susceptibility of PCW, which have no autolytic activity due to inactivation of autolysins by boiling, to freeze–thaw autolysin extract from the strains. The PCW of TlvDSMED1952 were digested slowly by its own autolysin extract (Fig. 3c), but significantly more rapidly by the autolysin extract from the strain MED1951. The autolysin extract from this strain digested its own PCW rapidly (Fig. 3c). This indicates that major changes in the peptidoglycan structure in TlvDSMED1952 sufficient to explain the decreased autolytic activity in this strain are unlikely. In support of this there was little difference in the lysostaphin digestion of PCW from the two strains (data not shown).
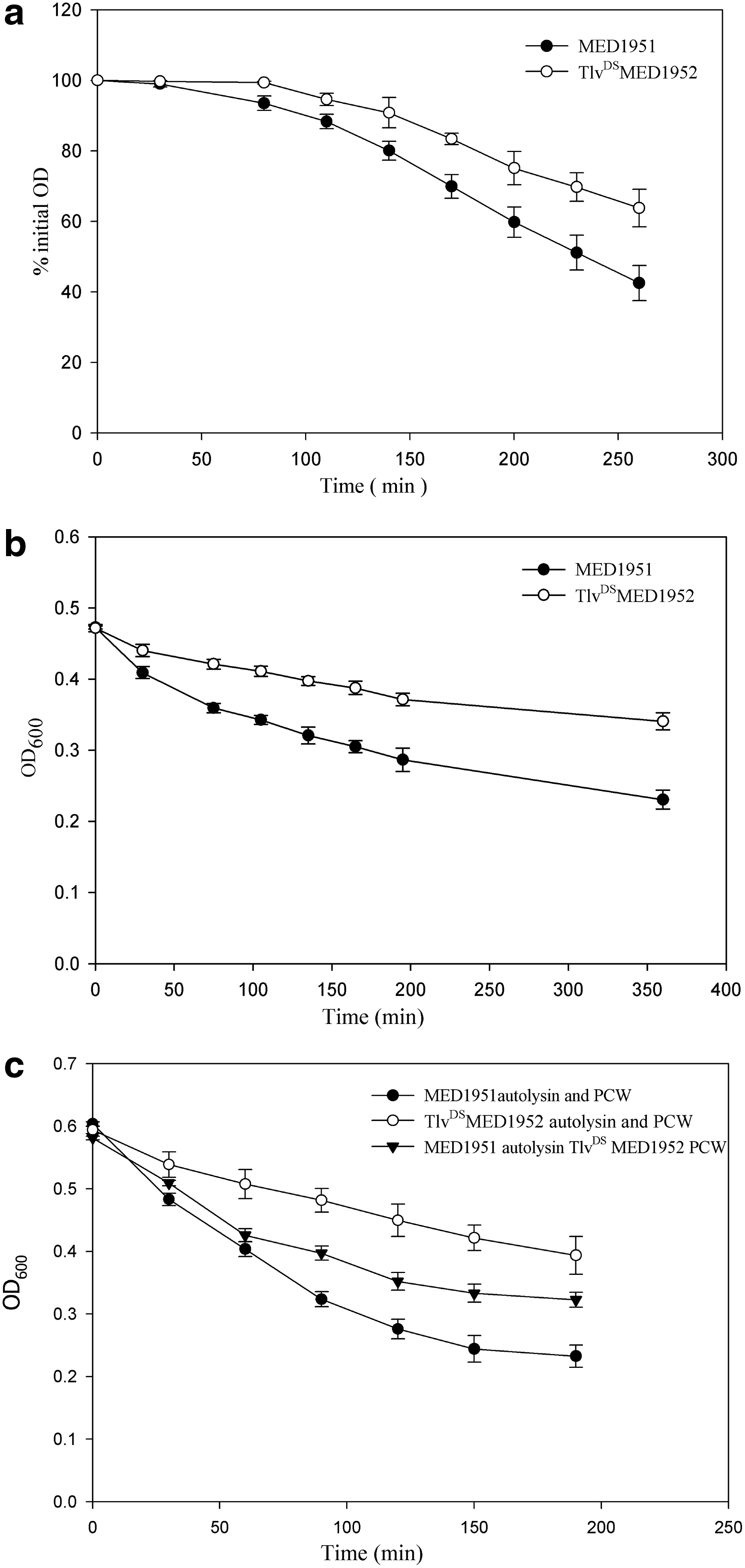
The autolysis activity of MED1951 and TlvDSMED1952.
We further characterized the freeze–thaw autolysin extract by studying the peptidoglycan hydrolase profile by zymography. When the peptidoglycan hydrolase profile of the autolysin freeze–thaw extracts of the strains were compared at equal protein concentrations, the TlvDSMED1952 extract showed different characteristics than the MED1951 extract (Fig. 4). The bands of the TlvDSMED1952 extract were less intense; it was devoid of lower molecular weight bands and had higher molecular weight bands, showing deficiency in autolysin processing.16,20 This is consistent with downregulation of protease genes (Table 1 and Supplementary Table S3 Supplementary material) that may be involved in peptidoglycan hydrolase processing.

Peptidoglycan hydrolase profiles of freeze–thaw autolysin extracts. Eight micrograms of protein were loaded on each lane.
Fatty acid composition, carotenoid content, and membrane fluidity
The total fatty acid compositions of the strains were determined and are shown in Supplementary Table S6. The content of straight-chain fatty acids was decreased to 9.4% in TlvDSMED1952 from 17.5% in MED1951, and the sum of branched-chain fatty acids increased to 90.6% from 82.5% in TlvDSMED1952 compared to MED1951. The anteiso:iso ratio was increased to 6.8 in TlvDSMED1952 compared to 3.4 in MED1951. These changes are indicative of increased membrane fluidity in strain TlvDSMED1952. 49 Perhaps, counter intuitively, increased membrane fluidity has been associated with decreased susceptibility to membrane-active molecules in S. aureus, including organic solvents and daptomycin.1,24,27,38
The OD465 value of the warm methanol extracts of TlvDSMED1952 were 73% of those of MED1951. This indicates no major changes in the carotenoid content, which can impact membrane fluidity.22,24
A statistically significant difference (p<0.05) in the polarization value (0.5±0.019) was detected for MED1951 compared to the polarization value of TlvDSMED1952 (0.46±0.016). The lower polarization value indicates a more fluid membrane in TlvDSMED1952.1,38
In conclusion, there were multiple changes in the transcriptome of the decreased susceptibility mutant that may be involved in its decreased telavaancin susceptibility. Also, expression of various virulence factors was decreased. There were significant cell envelope changes in the mutant involving thicker cell walls, decreased autolysis, and membrane fatty acid composition.
Footnotes
Acknowledgments
This work was funded by a grant from Astellas and grant 1R15AI084006 from the National Institutes of Health to BJW. The DNA microarrays were obtained through NIAID's Pathogen Functional Genomics Resource Center, Division of Microbiology and Infectious Diseases, NIAID, NIH.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.