
Abstract
We investigated the prevalence of plasmid-mediated quinolone resistance (PMQR) qnr genes by the polymerase chain reaction (PCR) in antibiotic-resistant bacteria isolates collected from aquatic environments in Jinan during 2 years (2008.3–2009.11). Genes were identified to variant level by PCR restriction fragment length polymorphism analysis or sequencing. qnrA1, qnrB2, qnrB4, qnrB6, qnrB9, qnrS1, and the new qnrB variant qnrB26 were detected in 31 strains from six genera (Klebsiella spp., Escherichia coli, Enterobacter spp., Proteus spp., Shigella spp., and Citrobacter spp.), four of which contained double qnr genes. Other PMQR genes, aac(6′)-Ib-cr and qepA, were found in 12 (38.7%) and 5 (16.1%) of 31 isolates, respectively; while qepA was found in Shigella spp. for the first time. Eight types of β-lactamase genes and eight other types of resistance genes were also present in the 31 qnr-positive isolates. The detection rate for five β-lactamase genes (blaTEM, blaCTX, ampR, blaDHA, and blaSHV) was >45%. Class 1 integrons and complex class 1 integrons were prevalent in these strains, which contained 15 different gene cassette arrays and 5 different insertion sequence common region 1 (ISCR1)–mediated downstream structures. qnrA1, qnrB2, and qnrB6 were present in three ISCR1-mediated downstream structures: qnrA1–ampR, sapA-like–qnrB2, and sdr–qnrB6. We also analyzed the horizontal transferability of PMQR genes and other resistance determinants. The qnr genes and some integrons and resistance genes from 18 (58.1%) of the 31 qnr-positive strains could be transferred to E. coli J53 AziR or E. coli DH5α recipient strains using conjugation or transformation methods. The results showed that a high number of qnr genes were associated with other resistance genes in aquatic environments in Jinan. This suggests that we should avoid over-using antibiotics and monitor aquatic environments to control the spread of antibiotic resistance genes.
Introduction
Enterobacteria are widely distributed in natural environments, clinics, and hospitals.12,20,33,45 Pathogens and opportunistic pathogens of the Enterobacteriaceae family cannot be ignored in human health, because symbiotic bacteria found in the human body may become opportunistic killers on occasions. Escherichia coli is the most common symbiotic organism but it is the main factor in UTIs, 31 while frequent culprits of pneumonia are Klebsiella spp. and Enterobacter spp. 3 The occurrence of integrons, PMQRs, and β-lactamase determinants have been reported widely in antimicrobial-resistant Enterobacteria and qnr genes are widely distributed in isolates of the Enterobacteriaceae family around the world.12,22,32,35,40,55,60,61
At present, quinolones are commonly used in clinical applications, animal husbandry, and aquaculture. As soon as antibiotics, antibiotic-resistant bacteria, and antibiotic resistance genes are released into the natural environment, they become susceptible to transfer via water. The aquatic environment may enrich the population of resistant bacteria and facilitate the transfer of resistance genes to human pathogens. 27 This may further exacerbate the problem of the development of clinical drug resistance. In this study, we analyzed the prevalence of PMQR qnr genes (qnrA, qnrB, and qnrS) in antimicrobial-resistant Enterobacteriaceae isolates collected from the aquatic environment during 2 years. Another two PMQR genes, aac(6′)-Ib-cr and qepA, and extended-spectrum β-lactamase (ESBL) or other β-lactamase (AmpC) genes were also identified in the qnr-positive isolates. We also analyzed the gene cassette arrays of class 1 integrons and their ISCR1 element-mediated resistance genes.
Materials and Methods
Sampling and bacterial isolation
Wastewater samples were collected at seven sampling points from surface and ground water in Xiaoqing River (XQR and GXQR), effluents at Thousand Buddha Hill Hospital (TBHH), Qilu Hospital (QLH), Shandong Traditional Chinese Medicine Hospital (STMH), the pigpen of Qilu pharmaceutical factory (PQPF), and No. 2 wastewater treatment plant at Jinan (WWTP), Shandong Province, China. The sampling sites were selected to achieve good coverage of the Jinan City area. After treatment in WWTPs, the water was discharged into XQR, before passing through different land use areas, including urban, agricultural, parkland, residential areas, and heavy and light industrial zones. TBHH, QLH, STMH, and PQPF were the main regions in which antibiotics, antibiotic-resistant bacteria, and antibiotic resistance genes were released into natural environments.
In general, qnr genes only reduce the susceptibility or mediate low-level resistance to quinolones, 49 so using quinolones for screening resistant strains may lead to a relatively low detection rate and even miss some important information. Previous studies found that qnrA and qnrB genes are related to complex class 1 integrons containing the transferable element ISCR1.14,56 However, we focused on the relationship between integron- and the PMQR genes; so, we used gatifloxacin (GAT) as a representative quinolone and six other antibiotics (see below in the same paragraph) related to integron-associated gene cassettes 16 to screen qnr-positive isolates. Each water sample (500 ml) was filtered through a sterile 47-mm-diameter membrane (0.22 μm; Millipore Corp.), which was then transferred into 100 ml of Luria–Bertani (LB) broth and incubated with shaking at 200 rpm at 37°C overnight. The cultures were diluted to 10−6, and 100 μl aliquots were spread onto MacConkey agar plates containing one of the qnr gene-related antibiotics, such as ampicillin (AMP, 100 μg/ml), ceftazidime (CAZ, 30 μg/ml), chloramphenicol (CHL, 30 μg/ml), GAT (5 μg/ml), kanamycin (KAN, 30 μg/ml), tetracycline (TET, 30 μg/ml), or trimethoprim (TMP, 30 μg/ml). The antibiotic ranges used in the antibiotic susceptibility tests of Enterobactereaceae followed the Clinical and Laboratory Standards Institute (CLSI) guidelines. 9 Individual colonies were picked based on different morphology and cultured routinely in 4 ml LB broth with the same antibiotics for further studies.
The polymerase chain reaction detection of qnr genes in isolates
The total DNA was extracted as described by Wilson. 59 Primers qnrAF/R, qnrBF/R, and qnrSF/R21,65 were used for the products of 516 bp qnrA gene, 469 bp qnrB gene, and 566 bp qnrS gene, respectively, to identify qnr genes in 391 antibiotic-resistant isolates. The genus of qnr-positive isolates was identified by sequencing the 16S rRNA using the primer pair K90 and K94. 64 The polymerase chain reaciton (PCR) was carried out using 50 pM of each forward and reverse primer with 1×PCR buffer (2 mM of magnesium sulfate, 50 mM potassium chloride, and 10 mM Tris-Cl, at pH 8.3), 200 μM of each dNTP, 0.5 U of Easy-Taq polymerase (TransGen Biotech), and 100 ng template DNA in a total volume of 20 μl. After amplification, the products were analyzed using 0.8% agarose gel electrophoresis and visualized by ethidium bromide staining.
Antibiotic susceptibility testing
The susceptibility of the qnr-positive strains and their transconjugants or transformants to antibiotics was tested using the disc diffusion method on Mueller–Hinton agar (MHA) plates, according to the CLSI guidelines. 9 Antimicrobial disks were purchased from BD Diagnostic Systems, which contained AMP (10 μg/ml), CAZ (30 μg/ml), cefepime (FEP, 30 μg/ml), cefoperazone (CFP, 75 μg/ml), cefotaxime (CTX, 30 μg/ml), CHL (30 μg/ml), CIP (5 μg/ml), fosfomycin (FOS, 200 μg/ml), GAT (5 μg/ml), KAN (30 μg/ml), nalidixic acid (NAL, 30 μg/ml), streptomycin (STR, 10 μg/ml), sulfisoxazole (300 μg/ml), TET (30 μg/ml), and TMP (5 μg/ml).
Identification of qnr variants by PCR restriction fragment length polymorphism analysis and DNA sequencing
The qnrB genes were further analyzed using the primers qnrBXF/BR and qnrB645XF/645BR (Table 1). To differentiate between the variants of qnrB, we analyzed the amplification products of qnrB using the primers qnrB645XF/645BR with a modified PCR restriction fragment length polymorphism (RFLP) method using the enzymes BglII, HindIII, and EcoRII. 51 Any PCR products of qnrB that we failed to identify using this primer set and RFLP were analyzed further by sequencing. The products of qnrB obtained using the primers qnrBXF/BR were also analyzed by enzymatic digestion and identified by sequencing. The amplification products of qnrA and qnrS were cloned directly into the pMD19-T vector (TA clone Kit; TaKaRa), and transformed into E. coli DH5α for sequencing and confirmation.
PCR, polymerase chain reaction; ISCR1, insertion sequence common region 1.
Analysis of aac(6′)-Ib-cr, qepA, and β-lactamase genes, and other resistance genes
We also screened for other resistance genes in the qnr-positive strains with the same primers that were used in previous studies for the PMQR gene qepA, 24 ESBL genes (blaSHV, blaTEM, blaCTX, blaCMY, and blaPER),5,6 AmpC genes (ampR, ampC, and blaDHA), 34 TET resistance genes (tetA, tetB, tetC, tetD, tetE, and tetG),25,26,30,67 CHL resistance gene catA, 15 aminoglycoside resistance genes [aadB, aphA, and aac(6′)-Ib], 13 and STR resistance genes strA-strB, 50 by PCR amplification in the same conditions as earlier, except for differences in the annealing temperature. To determine the PMQR gene aac(6′)-Ib-cr, the gene aac(6′)-Ib was amplified using the primer pairs aac(6′)-IbF/R, and the positive PCR product was analyzed further by digestion with FokI and direct sequencing. 24
Class 1 integron characterization and ISCR1 element analysis
Primers intI1F/1R were used to detect class 1 integrase, and the relative gene cassette arrays were amplified with the 5′ CS primer hep58 and 3′ CS primer hep59. 16 The ISCR1 element was checked by PCR using the primers 341A/341B 43 to produce an amplicon measuring 452 bp, or 513mF/513mR to produce another amplicon of 1,234 bp. ISCR1-mediated downstream fragments were obtained by PCR using the primers 513BF/qacEΔ1-B or self-formed adaptor PCR (SEFA PCR which can be used for chromosome walking). 58 The gene cassette arrays and ISCR1-mediated downstream fragments were analyzed by PCR-RFLP by EcoRII digestion, while PCR amplification products with different RFLP profiles, considered to be different gene cassette arrays, were ligated to the pMD19-T vector (TA clone Kit; TaKaRa) and transformed into E. coli DH5α for sequencing. The respective primers are described in Table 1.
Conjugation and transformation experiment
Conjugation and transformation experiments were performed with the qnr-positive strains to analyze the horizontal transferability of the detected qnr genes and other resistance genes. Conjugation experiments were carried out in a liquid mating-out assay with E. coli J53AziR as the recipient strain, according to the method of Wang, 56 and the transconjugants were screened on MHA plates supplemented with sodium azide (300 μg/ml) plus AMP (50 μg/ml) and TMP (30 μg/ml), and CIP (0.025 μg/ml), respectively. The plasmids of qnr-positive strains that failed in conjugation were purified and transformed into the home-made competent cells (E. coli DH5α) by the calcium chloride method 44 or electroporation using a Micropulser (Bio-Rad). 44 The transformants were selected on LB agar plates with AMP (50 μg/ml), TMP (30 μg/ml), and CIP (0.025 μg/ml).
Results
Prevalence of qnr genes and antimicrobial susceptibility
Of the total 391 resistant strains, we found 31 qnr gene-positive isolates, which were from the TBHH (11), QLH (14), STMH (4), and XQH (2) samples. The sequencing results for the 16S rRNA gene showed that they had spread among six different genera: Klebsiella spp. (16), E. coli (6), Enterobacter spp. (4), Proteus spp. (2), Shigella spp. (2), and Citrobacter spp. (1) (Table 2). Among the 31 qnr-positive isolates, the qnrA genes were present in 50.0% (1/2) of Proteus spp. and 25.0% (1/4) of Enterobacter spp. The qnrB genes were present in 87.5% (14/16) of Klebsiella spp., 100.0% (2/2) of Shigella spp., 50.0% (2/4) of Enterobacter spp., and 50.0% (1/2) of Proteus spp. The qnrS genes were present in 100.0% (6/6) of E. coli, 31.3% (5/16) of Klebsiella spp., and 25.0% (1/4) of Enterobacter spp. Three strains (0.8%) contained both qnrB and qnrS. Antimicrobial susceptibility testing with 15 agents showed that all 31 isolates were resistant to >3 antibiotics, and 17 were resistant to >8 antibiotics, as shown in Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/mdr). Of the 31 qnr-positive strains, the frequencies of resistance to quinolones were 23 (74.2%) for NAL, 19 (61.3%) for CIP, and 16 (51.6%) for GAT. The remaining isolates had intermediate resistance or were sensitive. The qnrA and qnrS genes were identified as qnrA1 and qnrS1 by sequencing.
T and C indicate transformants in E. coli Top 10 and transconjugants in E. coli J53, respectively.
Indicates resistance.
PMQR, plasmid-mediated quinolone resistance; ESBL, extended-spectrum β-lactamase; TET, tetracycline; CHL, chloramphenicol; STR, streptomycin.
Identification of qnrB variant
qnrB amplification fragments (681 bp) were detected in 4 out of 20 qnrB-borne strains (IK5, ICL11, J4, and LK4), while 645 bp fragments were found in all of the qnrB-positive strains except LK4. RFLP profiles with related restriction enzymes digestion of the qnrB genes are shown in Fig. 1A and B, and the process of qnrB variant identification is illustrated in Fig. 2. According to the original primer design, if a 681 bp qnrB fragment was successfully amplified, the 645 bp amplicon should be obtained as well. However, LK4 was an exception and it may contain a new qnrB variant. The sequencing results for the 681 bp qnrB fragment of LK4 showed 99% nucleotide sequence similarity and four nucleotide differences from the qnrB6 of pWCH-LM2 (EF520349) in Pantoea agglomerans. The deduced amino-acid sequence of this qnr gene had 226 amino acids, showing 99% similarity with qnrB6 except for two amino-acid differences (Glu-8-Val and Ser-23-Gly) and differed from qnrB1 by four amino acids. This gene was referred to as qnrB26 and its nucleotide sequence has been deposited into GenBank database with the accession number HQ386846, while its protein was designated QnrB26 based on the qnr nomenclature of Jacoby. 18 The phylogenetic tree was constructed to analyze qnrB26's relationship with the other qnrB variants (Supplementary Fig. S1). The 681 bp amplicons from the other three strains have the same RFLP profiles, and the sequencing results showed that all of them were qnrB6 (Fig. 1A).
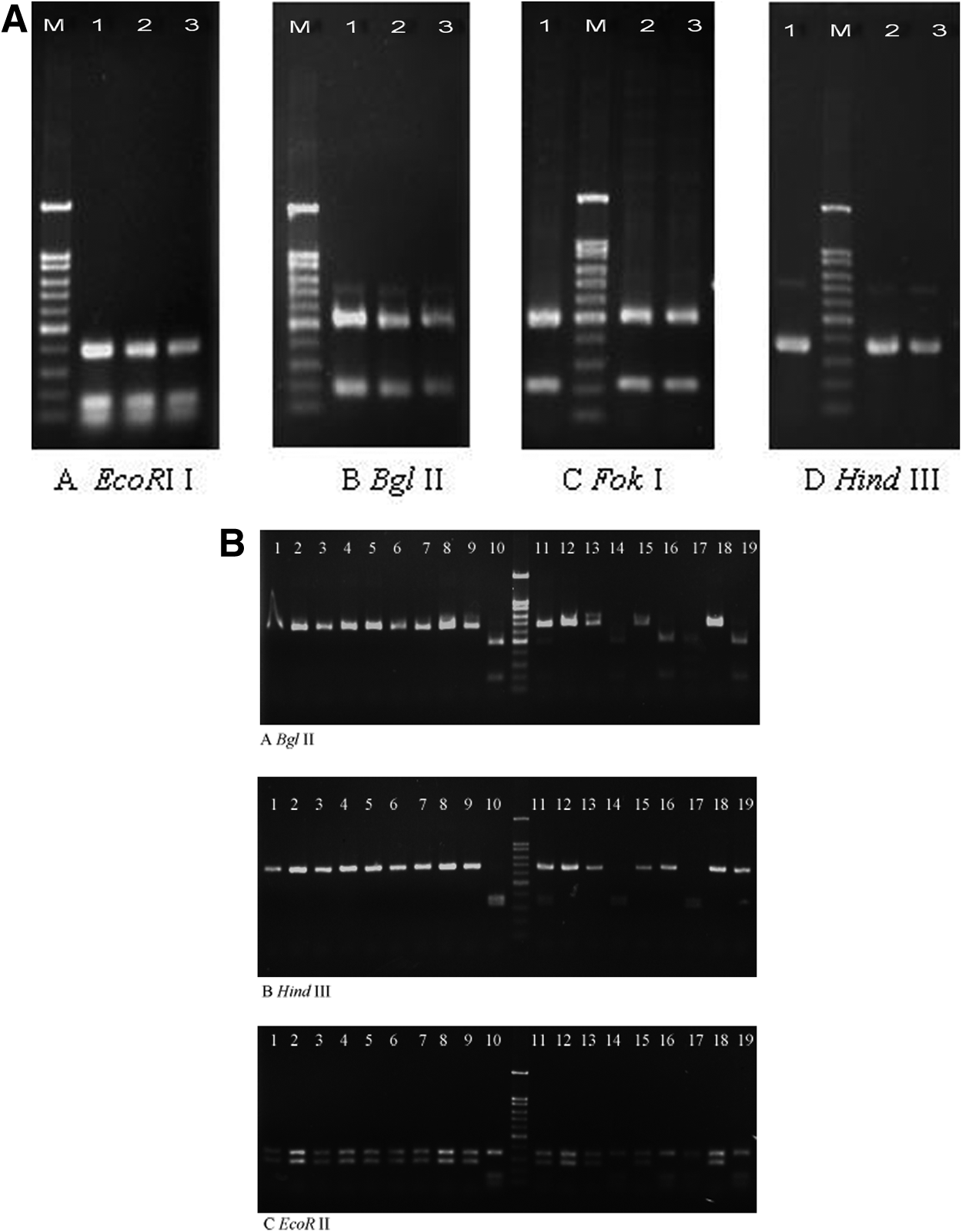
Restriction fragment length polymorphism (RFLP) profiles after EcoRII, BglII, FokI, and HindIII digestion of qnrB gene amplification products, where M indicates the marker (1.5, 1.0, 0.9, 0.8, 0.7, 0.6, 0.5, 0.4, 0.3, 0.2, and 0.1 kb).

The qnrB variant identification approach based on sequencing or the polymerase chain reaction-RFLP using BglII, HindIII, and EcoRII. B2, B4, B7, B8, B9, B11, B12, B19, B21, and B22 are abbreviations of the qnrB variants qnrB2, qnrB4, qnrB7, qnrB8, qnrB9, qnrB11, qnrB12, qnrB19, qnrB21, and qnrB22.
The BglII digestion results showed that IK11 might contain two qnrB variants, which were identified as qnrB4 and qnrB9 by sequencing. Further RFLP analysis of the 645 bp amplicons from the other 15 strains indicated that ISf1 and LTe6 contained qnrB2, while the remainder had qnrB4 (Figs. 1B and 2).
In conclusion, using two pairs of primers and RFLP analysis, the qnrB genes were identified to yield five variants, that is, qnrB2, qnrB4, qnrB6, qnrB9, and a new variant named qnrB26, which comprised 9.5% (2/21) qnrB2, 66.7% (14/21) qnrB4, 14.3% (3/21) qnrB6, 4.8% (1/21) qnrB9, and 4.8% (1/21) qnrB26.
aac(6′)-Ib-cr, qepA, β-lactamases, and other antibiotic resistance genes
The other PMQR genes, aac(6′)-Ib-cr and qepA, were detected in 12 and 5 of the 31 qnr-positive strains, respectively (Table 2). At least one type of β-lactamase (ESBL or AmpC) gene was detected in 27 out of 31 qnr-positive strains, including blaTEM 54.8% (17/31), ampR 51.6% (16/31), both blaCTX and blaDHA 48.4% (15/31), blaSHV 45.2% (14/31), and ampC, blaCMY, and blaPER 3.2% (1/31), respectively. Aminoglycoside resistance genes aphA1-Iab, aac(6′)-Ib, and aadB spread in 32.3% (10/31), 9.7% (3/31), and 6.5% (2/31) of the 31 qnr-positive strains, while the STR resistance gene strA-strB and CHL resistance gene catA were present in 71.0% (22/31) and 12.9% (4/31), respectively. TET resistance determinants, including tetA 45.2% (14/31), tetB 6.5% (2/31), and tetD 19.4% (6/31), were detected in 67.7% (21/31) of qnr-positive strains, including one strain that was positive for both tetA and tetB (Supplementary Table S1).
Class 1 integron characterization and ISCR1 element analysis
We found that 28 (90.3%) strains were positive for class 1 integrase (Table 2), while 25 (80.6%) strains contained 15 different gene cassette arrays, including 28 gene cassettes, as shown in Supplementary Table S1, most of which consisted of antibiotic resistance genes. These were TMP resistance genes (dfrA1, dfrA12, dfrA16, dfrA117, dfrA25, and dfrA27), aminoglycoside resistance genes [aadA1, aadA2, aadA5, aadA16, aadA23, aadB, aac3, aac(6′)-Ib, aac(6′)-II, and aac(6′)-IIc], rifampicin resistance genes (arr and arr3), CHL resistance genes (catB5 and catB3), an erythromycin resistance gene (ereA2), β-lactamases resistance genes (blaOXA-1 and blaOXA-10), and a quinolone resistance gene [aac(6′)-Ib-cr]. The remaining were a transposase gene (tnpA), an insertion sequence IS26, and two genes of unknown function (orfC and orfF). The sequences of these gene cassette arrays were 100% identical to previously reported arrangements. 16
Of the 31 qnr-positive strains, 21 (67.7%) strains, including 2 qnrA-, 18 qnrB-, and 1 qnrS-bearing strains, contained the ISCR1 element, which was confirmed by PCR (Table 2). To detect ISCR1-mediated downstream structures, four different-sized PCR amplicons (2.5, 2.6, 2.9, and 1.3 kb) with different RFLP profiles for eight strains and one representative strain of each profile were cloned for sequencing. In the other 13 strains, we used the SEFA PCR method to obtain one type fragment (2.4 kb) with the same RFLP profiles in four strains, and the representative was also cloned for sequencing. The remaining nine strains contained an ISCR1 element but lacked amplicons in the downstream region according to the two methods described earlier. As shown in Fig. 3, the 2,529 bp amplicon in the downstream region of ISCR1 in the two qnrA-positive strains (AK2 and IFs4) was the arrangement qnrA1–ampR and 100% identical to that of pKP96 (EU195449) in Klebsiella pneumoniae KP96 in terms of the nucleotide sequence. 46 A 2,645 bp PCR amplicon in the downstream region of ISCR1 was found in strains IK5, ICL11, and J4. They shared the same RFLP profile according to EcoRII digestion, and they were identified as having two open reading frames (ORFs), including an sdr gene encoding a 302 amino-acid short-chain dehydrogenase/reductase and qnrB6 encoding a 226 amino-acid quinolone resistance protein. This sequence shared 99% similarity with that of the plasmid pLC2 (EU543271) in K. pneumoniae. 24 A 2.9 kb amplicon was detected in qnrB-positive strains LTe6 and ISf1, which had 100% identity with a part of the sequence of the plasmid pKP1334/05-1 (FJ943244) from K. pneumoniae and the complex sul1-type integron (AM234698) from Salmonella enterica. 14 This fragment contained two ORFs, including a sapA-like gene encoding 180 amino acids, similar to a peptide transport periplasmic protein, and a qnrB2 gene encoding 214 amino acids quinolone resistance protein. Proteus vulgaris LK4 containing the new variant qnrB26 was amplified with a 1,322 bp ISCR1 element-mediated downstream fragment, while sequencing analysis detected an open reading frame containing 177 amino acids. A 2,406 bp amplicon of tnpU–armA was detected in four strains by SEFA PCR, which shared 99% identity with the partial sequence of the plasmid pNDM-HK (HQ451074) from E. coli strain HK-01. 17 Of these, three strains (Bsr3, GQr7, and GAp1) were qnrB positive, while one strain (GTa5) harbored both qnrB and qnrS.
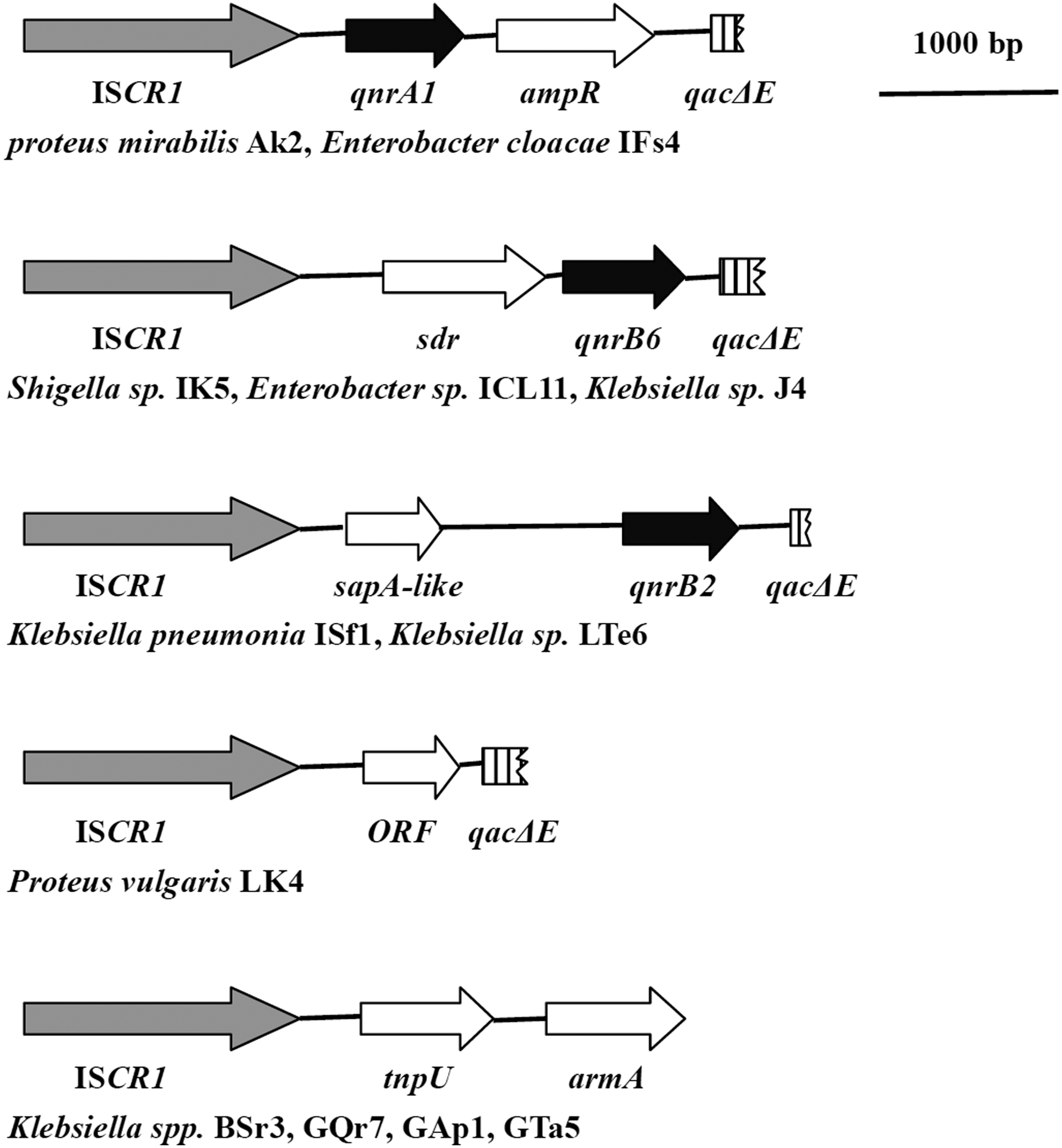
The qnr-positive strains with the insertion sequence common region 1 (ISCR1)–mediated downstream structures.
Horizontal gene transfer of quinolones and other antibiotic resistance genes
The results of the conjugation and transformation experiments shown in Table 2 and Supplementary Table S2 indicated that qnr genes could be transferred successfully from 18 (58.1%) strains, suggesting that they may be located in a plasmid. Of these, 15 were transferred by conjugation, including 1 qnrA gene from IFs4, 8 qnrB genes from GSr6, GAp1, ISr4, IK5, IK11, ICf1, ICf7, and J5, and 6 qnrS genes from Kp5, DQr1, ETe1, J14, ICf3, and LTe3. The remaining three qnr genes could be transferred by transformation, that is, one qnrB from LTe6 and two qnrS genes from ISr7 and LTe7. Strains IK11 or Kp5 contained two qnr genes (qnrB4 and qnrB9 or qnrB4 and qnrS1), but only one qnr gene could be transferred from either. This suggested that the two qnr genes had different locations in both strains. Other PMQR genes, aac(6′)-Ib-cr, from six (50%) strains could be co-transferred with the qnr genes, indicating that they may be located in the same plasmid with the qnr genes; whereas neither aac(6′)-Ib-cr nor qepA was transferred in isolation in this study. The resistance level of transconjugants and transformants to quinolones antibiotics NAL and CIP are described in Supplementary Table S2. Eight different class 1 integron gene cassette arrays, that is, dfrA16–aadA2, dfrA12–orfF–aadA2, aac(6′)-Ib-cr–arr3–dfrA27–aadA16, ereA2–arr–aac3–tnpA–ereA2–aac(6′)-IIc, aadA2, dfrA1–orfC, dfrA17–aadA5, and aadA23, from 12 strains were co-transferred with qnr genes; while the array dfrA12–orfF–aadA2 could also be transferred in isolation from 2 strains, GSr6 and BSr3. Some β-lactamase (ESBL or AmpC) genes were co-transferred with qnr genes from 48.1% (13/27) strains (i.e., blaSHV, blaTEM, blaCTX, blaDHA, blaCMY, and ampR) except that blaTEM and blaCTX from BSr3 were transferred without qnr genes. Apart from the 8 strains containing transferable qnrS genes, the ISCR1 element was found in the other 10 qnr-transferrable strains that could be co-transferred with qnrA or qnrB. In addition, 36.4% (8/22) of strA–strB, 40.0% (4/10) of aphA1-Iab, 33.3% (1/3) of aac(6′)-Ib-cr, 21.4% (3/14) of tetA, and 83.3% (5/6) of tetD displayed horizontal gene transfer with qnr genes.
Discussion
Our previous studies in Jinan detected the presence of a variety of class 1 and class 2 integrons from antibiotic-resistant Gram-negative bacteria in hospital wastewater, WWTPs, and even in rivers.16,63 In this study, we report the molecular characterization of a variety of PMQR determinants of multidrug-resistant Gram-negative bacteria from hospital wastewaters and receiving waters in this region, which expands our knowledge of drug resistance. The prevalence of plasmid-mediated quinolone resistant qnr genes was as high as 7.9% in 391 antibiotic-resistant strains isolated from aquatic environments in Jinan. The results showed that hospital wastewater is a major source of qnr-positive bacteria and should probably be treated more intensively. Previous reports have often focused on the presence of the qnr gene in cephalosporin antibiotic-resistant or β-lactamase-producing strains, as well as quinolone antibiotic-resistant strains.19,24,40,55,56 We extended the screening scope to other strains and found that the positive rate of the qnr gene is similar (8%, 6/78) to that in CIP-resistant isolates of E. coli from Shanghai, China, 56 which indicated that the qnr gene is related to diverse antibiotic-resistant strains and the prevalence of multidrug resistance. According to Jiang, 19 the positive rate of the qnr gene in 362 ESBL-producing strains collected from six provinces or districts in China was also 8%, which indicates that the prevalence of the qnr gene in Jinan is similar to the average in other areas of China. However, all the isolates in the earlier two reports of the qnr gene in China were clinical strains. The similar detection rate in isolates from water samples in our study, which were released into natural environments, reminds us of the antibiotic resistance crisis in the aquatic environment and the necessity for monitoring in a timely fashion to control the spread of antimicrobial resistance genes. Moreover, the prevalence of the qnr gene (7.9%) in our study was the same as that in 101 isolates from companion and food-producing animals in Guangdong, China. 24 The quinolones are used mostly in human and veterinary clinical medicine, as well as in livestock. The results of this study demonstrated the similar prevalence of qnr-positive isolates in water-related environments and those from human or animals. This highlights the antibiotic resistance crisis and the urgent need to ensure the rational use of antibiotics, as well as the scientific analysis of residual antibiotics.
According to the nomenclature system of qnr sequences, 37 qnrA has seven type variants and qnrS has eight types. In our study, the strains detected comprised only qnrA1 and qnrS1. A clinical surveillance study by Wu et al. 61 of Enterobacter cloacae in a Taiwanese hospital detected a very similar distribution to our study (the predominance of qnrB, followed by qnrS and a minority of qnrA). The most prevalent determinant, qnrB, has been reported in clinically isolated K. pneumoniae and other Enterobacteriaceae species in Asian countries.47,53,61 We also found that a high percentage of the environmental samples had various qnrB genes. The finding of qnrB26 complements the integrity and diversity of the qnrB information in a way and the number of qnrB variants has increased quickly from 23 in the year 2010 to 71 by now, 37 which will aggravate quinolones crisis without any doubt and indicates the necessity to monitor the qnr genes in time. The first qnrA was found in K. pneumoniae during 1994 in the United States, 28 whereas in this study we found no qnrA genes in any of 16 Klebsiella spp. strains and 6 E. coli strains, which was probably due to the geographical distribution of qnr genes. All of the E. coli isolates harbored the qnrS1 gene, which shows that the qnrS1 genes might be situated on plasmids that are prone to transfer between E. coli strains in this area. The prevalence of other two PMQR genes aac (6′)-Ib-cr and qepA was 38.7% and 16.1%, respectively, in combination with qnr genes, which could improve the survival prospects of bacteria in low-concentration quinolone environments, which will gradually accumulate mutations to produce high-level quinolone resistance to a greater extent. The efflux pump gene qepA has been reported in E. coli, Enterobacter spp., and Klebsiella spp. 24 and it is reported for the first time in Shigella spp. In this study, only one strain (IK5) was positive to one qnr gene (qnrB6) and other two PMQR genes [aac (6′)-Ib-cr and qepA].
In general, PMQR determinants have been reported to be associated with ESBLs or other β-lactamases. 39 It has been reported that qnr and aac(6′)-Ib-cr genes were detected in 8.0% and 9.9% of clinical ESBL-producing E. coli and K. pneumoniae, respectively, which were collected from six provinces or districts in China. 19 The horizontal transfer of plasmids harboring genes encoding ESBLs and PMQR determinants may have promoted this coresistance. 23 In the present study, the ESBL and AmpC genes blaCTX, blaSHV, blaTEM, blaCMY, blaPER, ampR, ampC, and blaDHA were identified in 87.1% (27) of the qnrA-, qnrB-, and qnrS-positive isolates, which are consistent with their close correlation. In addition, to the β-lactam antibiotics, 100% of the qnr-positive isolates were found to be resistant to AMP, and 71.0% (22) of the qnr-positive isolates were found to be resistant or intermediate-resistant CAZ, CFP, CTX, or FEP, which suggests that other determinants should be responsible for resistance to AMP. The ESBL gene blaPER was identified in association with the qnrA1 gene only in Proteus mirabilis AK2. A β-lactamase (PER) types are rare, and most blaPER-1 genes are associated with the Tn1213 transposon backbone and IS4 family. 36 We previously reported that blaPER-1 associated with ISCR1 was located downstream of aacA4-qnrVC4-aacA4-catB3 in Aeromonas punctata 159, which was isolated from the same site (THBB). 62
Antimicrobial susceptibility testing showed that >50% of the qnr-borne strains had resistance to quinolones, and all the minimal inhibitory concentrations of the quinolone antibiotics NAL and CIP in their transconjugants or transformants were ≤8 and 0.25 μg/ml, respectively; whereas none reached a resistant level. This might be due to the presence of qnr genes that promote mutations in the quinolone resistance-determining regions of gyrase or topoisomerase, which eventually produces high levels of quinolone resistance.4,42 Although the independent presence of qnr genes might not lead to clinically significant levels of quinolone resistance, the PMQR mechanism complements the traditional quinolone resistance pathway, which diversifies in drug-resistant forms of bacteria. In addition, qnr genes located in mobile genetic elements will promote the spread of quinolone resistance.
The frequency of class 1 integron in qnr-positive strains was high at 90.3%, and 46.4% (13/28) of them could co-transfer with qnr genes. A further study of the qnr gene-related ISCR1 element in complex class 1 integrons found five different structures in 38.7% (12/31) qnr-positive strains and three of the five type ISCR1 mediated downstream structures harboring qnrA1, qnrB2, or qnrB6 in seven strains (Fig. 3), which agreed with previous reports.14,56 This suggests that the qnr genes are in close contact with class 1 integrons or complex class 1 integrons containing ISCR1 elements. If these strains or resistance determinants come into clinical contexts via water circulation, the qnr genes present in complex integrons will integrate with other varieties of resistance genes, which will inevitably narrow the possible drug combinations available to clinical medicine for treating bacterial infections, thereby exacerbating the current challenging situation.
In total, 18 (58.1%) qnr genes could be transferred horizontally and most could be co-transferred with β-lactamase (13 of 18), which agrees with a previous report in which PMQR determinants were associated with ESBLs or other β-lactamases. 38 The qnr genes of 13 strains failed to be successfully transferred. Some factors that might affect this are low efficiency of conjugation or transformation for wild-type plasmids or perhaps those genes are located on chromosomes, although qnr genes were discovered first in plasmids.
The high frequency of PMQR genes associated with other resistance genes in environments and the high level of horizontal gene transferability probably aggravated the bacterial antimicrobial resistance crisis. In addition, we isolated 391 resistant strains from wastewater samples collected at the sampling sites, all of which were resistant to >3 antibiotics and most possessed integrons. In addition to the qnr-positive isolates, the remaining strains will be studied further to characterize their antibiotic resistance genes and integrons in order to analyze their antimicrobial resistance mechanisms.
In this study, we used a cultivation-based approach to study the qnr genes of Gram-negative strains from water samples, which extended the investigation scope, rather than focusing on specific clinical strains such as K. pneumoniae or E. coli.19,56 Moreover, our analysis of the qnr gene in the strains cultivated from water samples partially simulated a situation in which the nutrition in water is adequate. Inevitably, we may have missed some strains that were not suitable for the cultivation conditions; so, further work is required to make a comparison based on whole genome sequencing or fluorescent quantitative methods 10 to detect the presence of qnr genes in the aquatic environment. The results may also differ from the actual situation in an aquatic environment, although more detailed research could be conducted to identify qnr gene variant and to analyze their genetic environment of environmental strains, which may even find novel resistance mechanisms. In future studies, it will be better to combine cultivation-based approaches with whole genome methods, as well as analyze the residual antibiotics in water samples to assess the resistance situation in the aquatic environment. Thus, it is necessary to monitor the environment further to control the abuse of antibiotics and the spread of antibiotic resistance.
Footnotes
Acknowledgments
This work was supported by the grants from the National Natural Science Foundation of China (31170112), the Medical and Health Technology Foundation of Shandong (2011HZ050), the Life Sciences Student Training Program, and the State Key Laboratory of Microbial Technology, Shandong University.
The authors thank George A. Jacoby for providing E. coli J53 (Azide R), and Dan Wu, Liang Xu, Daxiao, Sun, and Xingrun Zhang for their technical assistance.
Disclosure Statement
The authors declare that there are no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.