
Abstract
Drug-resistant derivatives of serovar-specific virulence plasmids, such as pSLT, in clinically-relevant Salmonella enterica serovar Typhimurium strains, represent a threat for human health. We have analysed 14 S. Typhimurium isolates recovered in Italy and the United Kingdom from swine and from cases of human infection for the presence of virulence-resistance (VR) plasmids. They were negative for the multidrug resistance (MDR) region of the Salmonella genomic island 1 (SGI1), but expressed resistance to ampicillin, chloramphenicol, streptomycin/spectinomycin, sulfamethoxazole, and tetracyclines. The isolates were characterised by XbaI pulsed-field gel electrophoresis, multilocus sequence typing, and detection of resistance and virulence determinants (PCR/sequencing). Identification of VR plasmids was accomplished by PCR detection of bla genes (encoding ampicillin resistance), class 1 integrons and the pSLT virulence gene spvC. Plasmid analyses were performed by alkaline lysis, S1-nuclease digestion, replicon typing, conjugation, restriction analyses, and Southern blot/hybridization. Two blaOXA-1 positive isolates contained pSLT-derived plasmids related to pUO-StVR2. In nine isolates, eight from swine and one from a patient, MDR-conferring-IncFII-VR plasmids were detected. They contained the blaTEM-1 gene as well as a nonconventional class 1 integron with dfrA12-aadA2 gene cassettes in its variable region, and a sul3 gene in the 3′ conserved segment. Restriction analysis suggested a novel pSLT variant. The results obtained underline the role of swine as a potential reservoir for the blaTEM-1-IncFII-plasmids. The occurrence and spread of virulence- and MDR-conferring plasmids should be considered as a potential public health problem.
Introduction
In a previous study on the epidemiology of SGI1 in a European Salmonella strain collection, conducted in 2006/2007 by Amar et al. 1 within the European Union (EU) Network of Excellence Med-Vet-Net, several isolates showed the typical SGI1 resistance phenotype AMP-CHL-STR-SMX-TET, but were negative for SGI1. To elucidate if this resistance phenotype could be based on the presence of virulence-resistance (VR) plasmids, 14 S. Typhimurium isolates were selected for further characterization.
Materials and Methods
Bacterial strains
In the investigation performed by Amar et al., 1 66 S. Typhimurium isolates showed a SGI1-like resistance phenotype, although SGI1 was not present. From this series 14 isolates which shared a core resistance pattern to ampicillin, tetracyclines, sulfamethoxazole, and full or intermediate resistance to streptomycin (MIC≥32 mg/L, resistant; MIC=16 mg/L, intermediate) were selected for the present study. Ten of these isolates displayed the SGI1 characteristic pentaresistance (AMP-CHL-STR-SMX-TET) phenotype. Additional resistance to gentamicin, kanamycin, nitrofurantoin and/or trimethoprim was also present in some of the isolates. They were collected between 2004 and 2006 and were provided by following project partner institutes: Istituto Superiore di Sanità (ISS), Rome, Italy (five human isolates) and Animal Health and Veterinary Laboratories Agency (AHVLA), Weybridge, United Kingdom (nine porcine isolates). The strain designations used in this manuscript correspond to BfR lab codes, which are made up by the year of strain receipt followed by a serial processing number. S. Typhimurium SUO5 (containing pUO-StVR2) 22 was included as a control in the experiments involving analysis of ‘blaOXA-1-In8-like’-positive isolates.
Phenotypic analysis
To confirm the features of the isolates received by BfR, they were subjected to the following analyses. Serotyping was performed according to the White-Kauffmann-Le Minor scheme, 18 and phage typing following Anderson et al. 2 Isolates were tested by broth microdilution, according to the guidelines of the CLSI (M07-A8), 12 using custom-defined microtitre plates (TREK Diagnostic Sytems, East Grinstead, UK) with 14 antimicrobial substances: ampicillin (AMP), cefotaxime (FOT), ceftazidime (TAZ), chloramphenicol (CHL), ciprofloxacin (CIP), colistin (COL), florfenicol (FLO), gentamicin (GEN), kanamycin (KAN), nalidixic acid (NAL), streptomycin (STR), sulfamethoxazole (SMX), tetracycline (TET), and trimethoprim (TMP). 8 The evaluation of the results was performed according to the European Community Decision 2007/407/EC using the epidemiological cut-off values (ECOFFs) set by the European Committee on Antimicrobial Susceptibility Testing 14 (www.eucast.org).
All isolates were tested by disk diffusion method (M07-A8) 12 to determine their susceptibility to amoxicillin-clavulanic acid (AMC), nitrofurantoin (NIT), spectinomycin (SPE), and sulfamethoxazole/trimethoprim (SXT) (Oxoid Ltd., London, England). The results were interpreted using CLSI clinical breakpoints (M100-S21; for these antimicrobials no ECOFFs for disc diffusion were available for Salmonella spp.). 13
Detection of resistance and virulence determinants
Depending on their respective resistance phenotype all isolates were tested by PCR amplifications to assess the presence/absence of 30 resistance genes, the class 1 integrase gene, class 1, and class 2 integrons and 15 pSLT genes (spvC, rck, pefA, traT, repA2, traX-finO, oriT, samA, pefB, pefD, parA, parB, ccdAB, pefI/ORF7, and srgA). Primers and amplification conditions were described previously.6,10,20,25,44 Mutations in the gyrA gene in the nalidixic acid-resistant isolates and in integrons inserted gene cassettes were identified by DNA sequencing of the amplicons (Qiagen, Hilden, Germany). 21 The sequences obtained were analyzed using the BLAST program (http://blast.ncbi.nlm.nih.gov/Blast.cgi).
Plasmid analysis and Southern blot/hybridization
All isolates were screened for the presence of plasmids by alkaline lysis using the Kado and Liu method. 30 Bacterial plasmid DNA was separated on 0.8% vertical agarose gels. Escherichia coli reference plasmids R27 (169 kb), R1 (94 kb), RP4 (55 kb), and ColE1 (6 kb) were used as size standards. Plasmid sizes were determined by the S1-nuclease (Takara, Saint-Germain-en-Laye, France) digestion method followed by pulsed field gel electrophoresis (PFGE) as previously described.5,22 For size determination, the Lambda Ladder PFG Marker (New England Biolabs, Schwalbach, Germany) was used. The incompatibility (Inc) groups of plasmids were determined by PCR-based replicon typing.9,16 DNA obtained either from PFGE- or from alkaline lysis-generated plasmid gels was transferred by Southern blot onto nylon membranes and hybridized with different DNA probes specific for blaTEM-1, blaOXA-1, aadA1, strA, catA, cmlA, aphA1, tet(A), tet(B) and dfrA12 and also pSLT genes (spvC, and IncFII), as previously described. 19 The transfer potential of plasmid pMVN-VR42 (Table 2) was investigated by conjugation experiments using S. Typhimurium strain 08-02874 as donor and a sodium azide-resistant E. coli J53 derivative as recipient. Two parallel test series were conducted both at room temperature and 37°C, one in Luria-Bertani (LB) broth and the other one on solid LB (Oxoid) and M9 38 media on Millipore Membrane Filter (25 mm, 0,45 μm; Neolab, Heidelberg, Germany). Transconjugants were selected on eosin–methylene blue agar containing sodium azide (100 mg/L) and ampicillin (100 mg/L) or chloramphenicol (30 mg/L). Triparental mating was used to investigate the mobility of the plasmid pMVN-VR42. Overnight cultures of donor strain 08-02874, recipient E. coli J53 and a KAN resistant E. coli “helper” carrying plasmid pRK2013 15 were grown in LB broth. From each culture, 100 μl were pooled to a final volume of 300 μl, pelleted by centrifugation and resuspended in 100 μl bi-distilled water. The resuspension was streaked onto a LB agar plate and incubated for 4–6 hours at 37°C. Growth surface was washed in 2 ml LB broth, which was then plated on eosin–methylene blue agar containing the selective antimicrobials indicated above. For restriction fragment length polymorphisms (RFLP) analyses pMVN-VR42 and pSLT were extracted from S. Typhimurium 08-02874 and S. Typhimurium LT2, respectively, using the Qiagen Plasmid Maxi Kit (Qiagen). The 100 kb CTX-M-1 IncI1 plasmid pSal60 from S. Paratyphi B (dT+) 09-01801 (unrelated to pSLT; characterised within the “ESBLs” EU-SafefoodERA project 08176) was used as negative control in RFLP-hybridization experiments (data not shown). Each plasmid was independently digested with the restriction endonucleases PstI, SalI, SphI, HindIII, ClaI, and EcoRI under the conditions recommended by the manufacturer (Fermentas, Leon-Rot, Germany). Restriction endonuclease-generated fragments were separated on 0.6% horizontal agarose gels by conventional electrophoresis, transferred by Southern blot onto a nylon membrane and hybridized with the entire pSLT plasmid. 27 To prepare the probe, the pSLT plasmid of S. Typhimurium LT2 was extracted from a 0.45% agarose gel, purified by electroelution and labelled with the DIG DNA labelling kit (Roche, Mannheim, Germany) according to Herrero et al. 27
Macrorestriction analysis and multilocus sequence typing
All isolates were subjected to macrorestriction analysis of genomic DNA using the restriction endonuclease XbaI (Roche Diagnostics, Mannheim, Germany). A CHEF-DRIII SYS220/240 system (Bio-Rad Laboratories, Munich, Germany) was used to separate the fragments by PFGE. Agarose gel plugs were prepared and PFGE was performed using the PulseNet standardized protocol (www.pulsenet-europe.org). The resulting profiles were analyzed by recording the presence or absence of fragments larger than 33 kb. Profiles in which there were differences in two or more bands were designated as XT1, XT2, etc.; patterns with only one band difference were sub-designated using letters (e.g., XT1a, XT1b, XT1c).
For further typing, all isolates were analysed by multilocus sequence typing (MLST) according to Kidgell et al. 31 and to the recommendations of the MLST database hosted by the University College Cork, Ireland (http://mlst.ucc.ie/mlst/dbs/Senterica). For each isolate seven housekeeping genes aroC, dnaN, hemD, hisD, purE, sucA, and thrA were amplified by PCR and sequenced (Qiagen). The DNA sequences obtained were processed by the software SeqMan Pro (DNAStar, Inc., Madison, WI) and the alleles and sequence types subsequently determined using the MLST database website.
Results
Identification of bla-positive isolates and integron detection
All 14 S. Typhimurium isolates were analysed by PCR for the presence of bla genes and class 1 integrons. Twelve isolates generated amplicons for blaTEM-1 (503 bp), while the other two tested positive for blaOXA-1 (708 bp). The gene blaPSE-1 was not identified in any of the isolates. The results of the PCR-screening for further resistance genes are summarized in Table 1.
Country abbreviations: DK, Denmark; E, Spain; I, Italy; UK, United Kingdom.
VR plasmids are marked in thick numbers.
For abbreviations, see Methods. Resistance phenotypes were assessed by the broth microdilution method following the guidelines of the CLSI (M07-A8). 12
Resistance phenotypes for AMC, NIT, SPE, and SXT were assessed by disk diffusion method. 12 The breakpoints used were the CLSI breakpoints (M100-S21). 13
Variable regions of class 1 integrons; indicated are their approximate lengths of the PCR product using the 5′CS and 3′CS primers 32 and the inserted gene cassettes.
The 14 isolates were screened for class 1 integrons by the classical PCR analysis using the 5′CS/3′CS primers (Table 1). 32 The two blaOXA-1-positive isolates generated a PCR product of 2,000 bp containing the blaOXA-1 and aadA1 genes. From the 12 isolates positive for the blaTEM-1 gene, only one generated a class 1 integron PCR-product (1,600 bp), which carried dfrA1-aadA1 gene cassettes. Ten blaTEM-1 isolates, negative for this PCR-amplification, were positive for the class 1 integrase-coding gene intI1 and the antimicrobial resistance-conferring genes dfrA12, aadA2, cmlA1 and sul3. The presence of a nonconventional (no amplification with the cited primers) class 1 integron described in S. enterica isolates from Portugal was therefore, suspected. 3 This integron contains an unusual 3′ conserved segment with the sulfamethoxazole resistance conferring gene sul3. The presence of this integron was confirmed by PCR using primer sets for intI1/dfrA12 (1,021 bp), dfrA12/aadA2 (1,697 bp), and cmlA1/aadA1 (1,577 bp). The size of the expected amplicons was determined by means of the nucleotide sequence of the sul3-carrying integron deposited by Antunes et al. in the EMBL database (accession number EF051037). 3 The remaining blaTEM-1 positive isolate did not possess any class 1 integron.
Characterization of blaOXA-1-In8-like positive isolates
The two S. Typhimurium DT120 isolates from cases of human infection in Italy (08-02878 and 08-02879), and which were ‘blaOXA-1-In8-like’-positive, showed resistance phenotype [AMP-AMC]-CHL-[STR(r/i)-SPE(r/i)]-TET-SMX-NIT, as displayed by the control strain SUO5, which carries the pUO-StVR2 plasmid. The isolates were positive for the spvC virulence gene typical for pSLT. Subsequent plasmid analysis revealed that both isolates from Italy lacked pSLT but contained an IncFII plasmid of ∼140 kb (the same Inc group as pSLT) (Table 1). These IncFII plasmids also hybridized with the spvC and the blaOXA-1probes. These hybrid plasmids also carried the ∼140 kb pUO-StVR2 genes catA1, aadA1, tet(B), and sul1, and were therefore, assigned to the S. Typhimurium pUO-StVR2 group. The screening for other selected typical pSLT virulence genes showed that both isolates were, like control strain SUO5, positive for rck, pefI, traT, traX, oriT, samA, parA, parB, ccdAB, and srgA genes. No amplification of the pefA, pefB, pefD genes of the pef (plasmid-encoded fimbriae) operon, the repA2 of the IncFIB replication region, and the rsk locus (regulation of serum killing), were observed, suggesting the absence of these genes as reported in pUO-StVR2 (accession no. AM991977) (Table 2).
Country abbreviations: DK, Denmark; E, Spain; I, Italy; UK, United Kingdom.
VR plasmids are marked in bold numbers.
nd, not determined.
Both blaOXA-1-In8-like-positive isolates were subjected to XbaI-based PFGE analysis. By this method, they were assigned to two closely-related XbaI PFGE profiles designated XT1 and XT1a (XT1a possessed an additional band of ∼230 kb, data not shown). The pUO-StVR2 control strain SUO5 also showed the XT1 PFGE profile. The results are summarized in Table 2.
Characterization of blaTEM-1-dfrA12 positive isolates
Two of the 12 blaTEM-1-positive isolates, ascribed to MLST type ST34, only harboured plasmids <30 kb and were negative for the spvC gene. The remaining 10 isolates, all of them ascribed to ST19, carried the spvC gene and an integron associated to sul3. They also contained one IncFII plasmid ≥110 kb (Fig. 1). The ‘classical’ 94 kb pSLT plasmid was not detected in any of these isolates. All IncFII plasmids hybridized with probes for the genes spvC, blaTEM-1, and dfrA12. These 10 isolates that contained VR plasmids showed the common core-resistance pheno-/genotype AMP-CHL-[STR-SPE]-SMX-TMP-SXT/blaTEM-1-cmlA1-[aadA1-aadA2]-sul3-dfrA12. In some isolates additional resistances to KAN/aphA1 (in two isolates), TET/tet(A) (eight), and NIT (three) were detected. In these isolates, five different virulence genotypes were identified. All isolates were positive for the genes spvC, rck, pefI, traT and traX. With the exception of one isolate, all the pef genes were present.
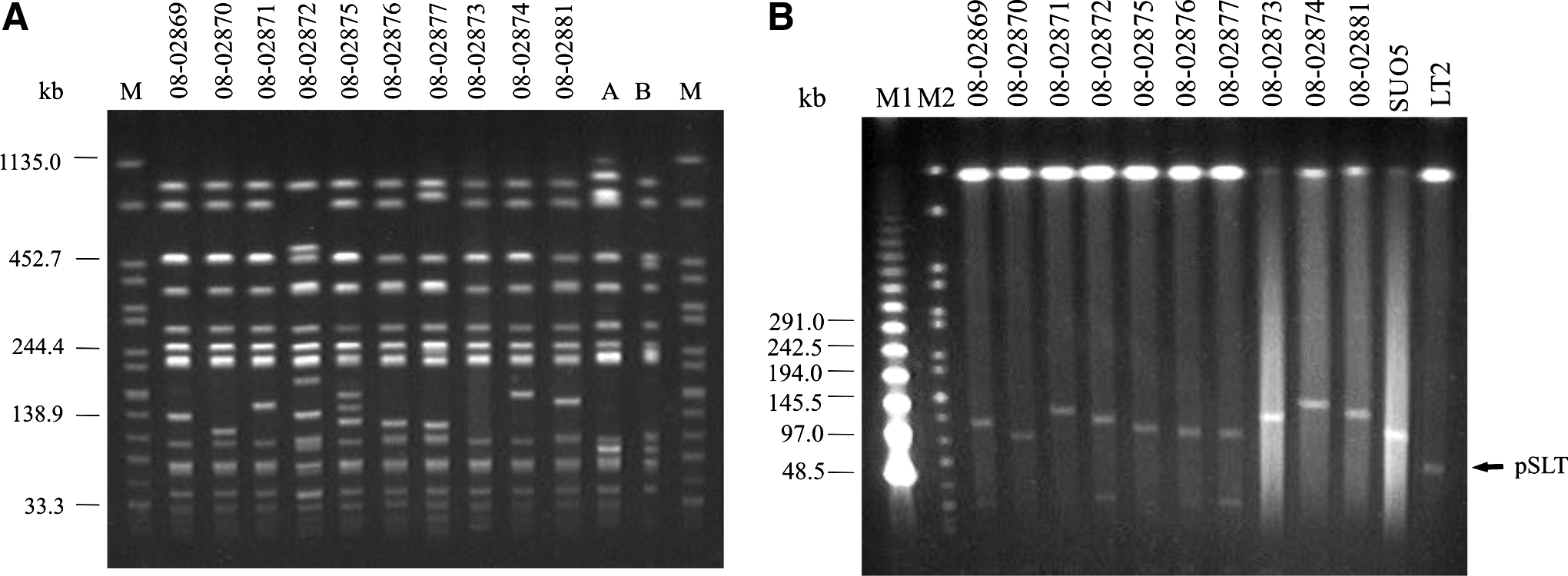
PFGE analysis revealed related but slightly different profiles for each isolate. One isolate was assigned to the XT1 profile (also detected in the blaOXA-1-In8-like-positive isolates) and two exhibited the closely-related patterns XT1b and XT1c. The results are shown in Table 2 and Fig. 1.
Characterization of selected VR plasmids in blaTEM-1-dfrA12 positive isolates
The three isolates (08-02873, 08-02874, and 08-02881) that showed an identical or highly similar XbaI-PFGE profiles (XT1, XT1b, XT1c) to those of the pUO-StVR2-carrying isolates (XT1 and XT1a) were subjected to further characterization. By plasmid DNA hybridization, the genes aadA1, cmlA, and strA were found to be located on the VR-plasmids. The tet(A) gene, present in the three isolates was located on smaller plasmids. The aphA1 gene, harboured only by the isolate 08-02873, was also located on a small plasmid. The IncFII plasmids carried by these strains were designated pMVN-VR41, pMVN-VR42, and pMVN-VR49, respectively (Tables 1 and 2).
The ∼160 kb plasmid pMVN-VR42 was selected for further characterization. The plasmid pMVN-VR42 could not be transferred by conjugation nor mobilized. Restriction analysis and hybridization of pMVN-VR42 and pSLT revealed individual profiles (Fig. 2). Nevertheless, many restriction enzyme-generated fragments of pMVN-VR42 had counterparts in pSLT, thereby suggesting a close relationship between the two plasmids. The CTX-M-1 IncI pSal60 plasmid, used as a negative control, generated restriction patterns which neither corresponded to those of pSLT nor hybridized with the pST90 probe (Supplementary Fig. S1, Supplementary data; Supplementary Data are available online at www.liebertpub.com/mdr).
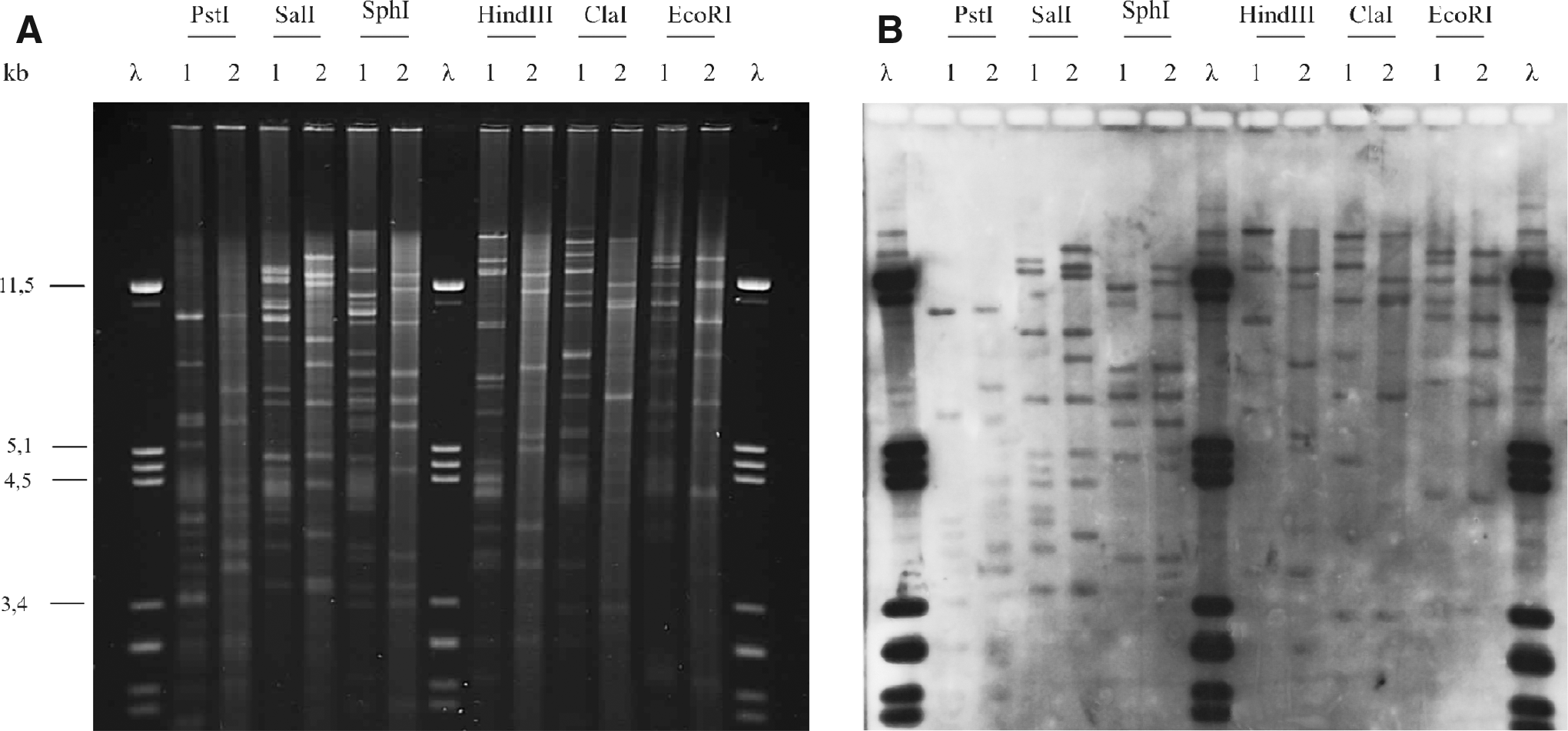
Relation between pMVN-VR42 and pSLT.
Discussion
The evolution from avirulence to pathogenicity often entails the acquisition of external DNA virulence-associated fragments. 33 Different combinations of such fragments may result in the appearance of novel antimicrobial resistance and virulence phenotypes, which may then be subjected to natural selection according to changing environmental conditions. 4 The first references for an association of the spv virulence gene with additional antimicrobial resistance determinants date back to studies in the 1990s.23,40 In one of these studies, plasmids of ∼120 kb, present in isolates of S. Typhimurium DT193, were found to exhibit endonuclease-generated restriction patterns with considerable congruence with that of pSLT, the serovar-specific S. Typhimurium virulence plasmid. 40 Resistance derivates of serovar-specific virulence plasmids were also detected in in S. Enteritidis and highly invasive S. Choleraesuis. 34 In 2002, MDR S. Typhimurium isolates that contained the pSLT-derived VR plasmid pUO-StVR2 were first described in cases of human infection in Spain. Subsequently strains possessing such virulence/resistance plasmids have played an important role in human salmonellosis in that country, being implicated in sporadic cases, outbreaks,20,25,26 and in extraintestinal disease, thereby confirming their invasive potential. 34 Similar strains of S. Typhimurium have also been detected in the UK, and are suspected to be present also in Portugal and Norway. 28 To date, S. Typhimurium isolates positive for pUO-StVR2 have been mainly isolated from clinical samples, but have also been detected in foods and food-producing animals. 34 One of the characteristics of these strains is their resistance to nitrofurantoin.28,34 In the present study, pUO-StVR2-like plasmids were found in two DT120 isolates from cases of human infection in Italy. Identical or highly similar PFGE profiles (XT1, XT1a) suggest a relationship to the Spanish pUO-StVR2-harbouring isolates. An In8- blaOXA-1 (also designated as blaOXA-30) containing VR plasmid different from those characterized in this study, was described previously in an Italian S. Typhimurium isolated from a patient. Apart from In8, this plasmid carried a second integron (In7), which contained the gentamicin and kanamycin resistance-conferring gene aadB. 45
In 10 other S. Typhimurium isolates, MDR-conferring IncFII-plasmids containing the blaTEM-1 gene were identified. These isolates originated from swine in the UK (nine isolates) and from a patient in Italy; three of these isolates also showed resistance to nitrofurantoin. Some of these isolates revealed the same or very similar PFGE profiles (XT1, XT1a and XT1b) like the isolates of the pUO-StVR2 group (XT1 and XT1a), but the genetic resistance background of both VR plasmid groups was different. These plasmids contained a nonconventional class 1 integron that was associated with the sulfamethoxazole conferring gene sul3 downstream of the variable region dfrA12-aadA2-cmlA1-aadA1. In Salmonella, sul3-integrons located on plasmids of various sizes (≥100 kb) were also found in isolates collected in Portugal. 3 The association of these integrons with epidemic plasmids and MDR Salmonella clones is assumed to contribute to the maintenance of antimicrobial resistance and to its spread from food producing animals to humans. 3 The restriction analysis carried out for pMVN-VR42 suggested this plasmid to be a novel pSLT-derived variant. Large, nonconjugative plasmids (∼150 to 200 kb) also carrying resistance and virulence genes were detected in Spanish isolates belonging to the monophasic S. Typhimurium variant 4,5,12:i:-. These isolates showed resistance up to seven antimicrobials AMP-CHL-GEN-STR-SMX-TET-TMP encoded by the genes [blaTEM-1-cmlA1-aacC4-(aadA1-aadA2-strA)-(sul1-sul2-sul3)-tet(A)-dfrA12]. Unlike the plasmids described in this study, they belonged to IncA/C±IncN, and were considered as resistance-virulence plasmids.17,19 Other blaTEM-1 associated VR plasmids were recently identified in monophasic S. Typhimurium 4,5,12:i:- isolates also collected from swine and patients in the UK (Patricia García, unpublished results), in S. Typhimurium isolates from cattle in Japan, 39 and in Spanish S. Enteritidis isolates from patients. 35
The present study has shown that within S. Typhimurium several variants of VR plasmids can be found. The results obtained underline the role of swine as a potential reservoir for the blaTEM-1-IncFII-plasmids. The occurrence and spread of virulence- and MDR-conferring plasmids should be considered as a potential public health problem, because on one hand the use of antimicrobial agents promotes the coselection of both resistance and virulence traits and might complicate the clinical treatment of invasive infections. On the other hand horizontal transfer facilitates the spread of these plasmids. 34
Footnotes
Acknowledgments
We thank the technical staff of the NRL-Salm at the Federal Institute for Risk Assessment for excellent technical assistance and the Med-Vet-Net WP21 Project Group, which provided the isolates used in this study. We also thank A. Herrero for invaluable advice and L. Martínez Martínez (Hospital Universitario de Valdecilla, Santander, Spain), A. Carattoli (ISS, Rome, Italy), and the ESBLs-SafefoodEra Consortium (EU-SAFEFOODERA project 08176) for kindly providing reference strains. This work was supported by the Federal Institute for Risk Assessment (BfR: 46-001 and 45-004) and the EU Network of Excellence Med-Vet-Net (WP21).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.