
Abstract
The mechanisms of high-level carbapenem resistance in Klebsiella pneumoniae isolated in Japan were investigated. High-level carbapenem-resistant K. pneumoniae Mkp4437 and a less carbapenem-sensitive K. pneumoniae strain, Mkp4365, were recovered from the same patient. These two strains were found to be homologous by pulsed-field gel electrophoresis, and both strains contained blaIMP-1, blaDHA-1, blaCTXM-14, blaTEM-1, and blaSHV-1. Based on the sodium dodecyl sulfate–polyacrylamide gel electrophoresis analysis, the lack of Ompk36 was observed in Mkp4437. Direct sequencing of the ompK36 gene demonstrated that a new insertional sequence in the open reading frame of the ompK36 gene was found in Mkp4437. Three clinical isolates (minimum inhibitory concentration [MIC] 2–4 mg/L to imipenem) were identified upon screening the strains of K. pneumoniae isolated in the University hospital with MICs of ≥1 mg/L to imipenem. Interestingly, these three isolates all lacked OmpK36. Conjugation of the plasmid harboring IMP-1 to these three OmpK36-deficient strains led to the isolation of high-level carbapenem-resistant transconjugants. In conclusion, the mechanisms of high-level carbapenem resistance in K. pneumoniae entail not only the production of IMP-1 β-lactamase but also the lack of OmpK36. It is vital to monitor for the presence of less carbapenem-sensitive K. pneumoniae strains, which lack OmpK36, because blaIMP-1 transmission to these strains may result in isolates with a high-level carbapenem-resistant phenotype.
Introduction
Although the outbreak of carbapenem-resistant K. pneumoniae has been observed in some countries,8,13,18,21,32,38,40,41 KPC or NDM-1 β-lactamase is uncommon in Japan.3,43 The most clinically important carbapenemase in Japan is thought to be the MBL, IMP-1, which has already been disseminated into several members of the Enterobacteriaceae family.10,34,35 High-level carbapenem-resistant K. pneumoniae or K. oxytoca harboring IMP-1 β-lactamase have been reported in 1996, 33 2007, 7 and 2012 36 in Japan. However, some reports indicated that K. pneumoniae or K. oxytoca strains that produce IMP-1 β-lactamase are not always resistant to carbapenems.14,26 Therefore, the mechanisms of high-level carbapenem resistance in K. pneumoniae or K. oxytoca strains that produce IMP-1 β-lactamase remain poorly understood. In this study, we investigated the mechanism of high-level carbapenem resistance in K. pneumoniae by the analysis of the plasmid and outer membrane protein (OMP) of both high-level and low-level carbapenem-resistant K. pneumoniae strains.
Materials and Methods
Bacterial strains
Among the total of 3,151 isolates stocked between 2008 and 2011 in the University of Occupational and Environmental Health Hospital in Japan, three K. pneumoniae strains, Lkp8012, Lkp8026, and Lkp8027, with minimum inhibitory concentrations (MICs) of 4, 2, and 2 mg/L, to imipenem were found after screening for the isolates with MIC ≥1 mg/L to imipenem. In this study, the categories of less carbapenem-sensitive and high-level carbapenem-resistant were defined as MIC 1–4 mg/L and MIC ≥16 mg/L to imipenem, respectively. K. pneumoniae Mkp4437 (high-level carbapenem-resistant strain, MIC 32 mg/L to imipenem) and K. pneumoniae Mkp4365 (less carbapenem-sensitive, MIC 1 mg/L to imipenem) were obtained from a multicenter bacterial collection of the clinical isolates at the Hibiki Research Group for Clinical Microbiology. Mkp4437 and Mkp4365 were isolated from the urine of a patient who was hospitalized for acute pyelonephritis in the Western part of Japan in 2008. K. pneumoniae Lkp8001 was isolated from the sputum of a patient with pneumonia, and K. pneumoniae ENkp8003 was isolated from the urine of a patient with acute cystitis in our hospital.
Bacterial species were identified using the Vitek2 system (BioMerieux Japan Ltd.). Commercially available competent Escherichia coli DH5α (Toyobo Co., Ltd.) were used in the transformation experiments. Rifampin-resistant ML4901, E. coli ML4905 (kindly presented by Dr. Shizuko Iyobe in the Gunma University School of Medicine), was used for the recipient strain in conjugation experiments. The transformant TF-11 was obtained by a transformation experiment using DH5α and plasmids that had been extracted from Mkp4437. The transconjugant TS-11 was obtained by a conjugation experiment using ML4905 as the recipient strain and Mkp4437 as the donor strain. To obtain K. pneumoniae strains harboring blaIMP-1 of Mkp4437, a conjugation experiment was performed using TS-11 as the donor strain. Lkp8012/p4437, Lkp8026/p4437, and Lkp8027/p4437 were obtained by a conjugation experiment using Lkp8012, Lkp8026, and Lkp8027 as recipient strains and TS-11 as the donor strain (Table 1). The methods of transformation, conjugation, and curing are described below.
High-level carbapenem-resistant: MIC ≥16 mg/L to imipenem.
Isolates from the same hospitalized patient.
Less carbapenem-sensitive: MIC 1–4 mg/L to imipenem.
MIC, minimum inhibitory concentration.
Antimicrobial susceptibility testing
Antimicrobial MICs were determined using a twofold serial agar dilution on Muller-Hinton agar (Eiken Chemical Co.), as recommended by the Clinical and Laboratory Standards Institute. 4 Antimicrobial agents were purchased from commercial sources or provided by each manufacturer. 2-Mercaptopropionic acid (MPA; Nacalai Tesque, Inc.; 128 mg/L) was used with meropenem or ceftazidime to inhibit MBL in agar dilution. 3-Aminophenylboronic acid (APB; Tokyo Kasei Kogyo Co., Ltd.; 300 mg/L) was used with meropenem or ceftazidime to inhibit Class C β-lactamases. 42 Both MPA and APB were used with meropenem or ceftazidime to inhibit both MBL and Class C β-lactamases.
PCR and DNA sequencing of OMP and β-lactamase genes
Class A, Class B, and Class C β-lactamases were detected by PCR with primers specific for the blaKPC, blaCTXM, blaTEM, blaSHV, blaIMP, blaCMY, and blaDHA as described by Muratani et al. 26 and Bradford et al. 2 To detect the ompK35 and ompK36 genes, the following primer pair was designed: ompK35_52F (5′-GACACCAAACTCTCATCAATGG-3′), ompK35_1206R (5′-TTGGTAAACGATACCCACGG-3′), ompK36-327F (5′-AAA CCCAGATCAACGACCAG-3′), and ompK36-1207R (5′-ACC ACGTCGTCGGTAGAGAT-3′). These primers were used to amplify a 1,155-bp fragment from ompK35 gene of KT755 6 (GenBank accession no. AJ011501), and a 881-bp fragment from the ompK36 gene of the imipenem-sensitive strain VM522, (GenBank accession no. FJ577673).
The following amplification program was used for the PCR assays: 94°C for 2 minutes, and 32 cycles of 94°C for 30 seconds and 56°C for 30 seconds, and 72°C for 1 minutes. The PCR products were separated by electrophoresis on a 1.8% agarose gel, stained with ethidium bromide, and compared with amplified DNA from positive controls.
The DNA of structural genes was sequenced using the ABI PRISM Big Dye Terminator Cycle sequencing Ready Reaction kit with AmpliTaq DNA polymerase (Perkin-Elmer, Inc.) on an ABI PRISM 377 DNA sequencer (Perkin-Elmer, Inc.) using the sequencing primers described by Kaczmarek et al. 12 The following additional sequence primers were designed: ompK36-28F (5′-CAGCACAATGAATATAGCCG-3′), ompK36-851R (5′-CAGACCGCCGGTGTAGGTTT-3′), ompK36-841F (5′-CGACAACGCTGAAACCTACA-3′), and ompK36-1827R (5′-GACACCAGATAGCGGGAGAT-3′) from the ompK36 gene of the K. pneumoniae NTUH-K2044 strain (GenBank accession no. FJ577673). 11 Sequence analysis and alignments were performed using the NCBI Blast program and compared with the sequences deposited in the GenBank database (http://blast.ncbi.nlm.nih.gov/Blast.cgi).
Pulsed-field gel electrophoresis typing
XbaI-digested genomic DNA was prepared according to the manufacturer's instructions (Bio-Rad Laboratories, Inc.). The DNA fragments generated were then separated as described by Muratani et al. 26 The XbaI restriction patterns of the DNAs were analyzed and interpreted according to the criteria proposed by Tenover et al. 39
Plasmid
The plasmid DNA was extracted from each isolate with a HiSpeed Plasmid Midi Kit (QIAGEN K.K). The size of the plasmid DNA was determined using S1 nuclease digestion 1 by the CHEF mapper pulsed-field gel electrophoresis (PFGE) system on a 1% agarose gel using the following conditions: pulse time ramped linearly from 3 to 60 seconds, run time 22 hours, temperature 14°C, and 6 V/cm constant voltage.
Conjugation and transformation
Bacterial conjugation was performed using a transconjugant of Mkp4437 and ML4905 as the donor strain and Lkp8012, Lkp8026, and Lkp8027 as the recipient strains. These isolates were mixed, suspended in 0.9% saline, and plated on CHROMagar Orientation plates containing 8 mg/L meropenem. Blue-colored colonies that grew within 48 hours at 35°C were selected for further analysis. Bacterial conjugation was performed in a lysogeny broth using rifampin-resistant E. coli ML4905 as the recipient strain to analyze the effect of IMP-1 β-lactamase production. The donor and recipient strains were cultured separately to the early log phase in 10 ml of antimicrobial-free lysogeny broth, after which 0.9 ml of the recipient strain and 0.1 ml of the donor strain were mixed. The bacteria were suspended in 0.9% saline and plated on CHROMagar Orientation plates (Kanto Chemical Co., Inc.) containing 500 mg/L rifampin and 10 mg/L ceftazidime. Pink-colored colonies that grew within 48 hours at 35°C were selected for further analysis. The transfer experiment was carried out in mixed lysogeny broth cultures using plasmids extracted from Mkp4437 without electroporation, using E. coli DH5α as the recipient strain to analyze the effect of IMP-1 β-lactamase production. Transformants were selected on lysogeny broth agar plates containing ceftazidime at 10 mg/L.
OMP isolation
K. pneumoniae strains were grown overnight in a lysogeny broth at 37°C, and OMPs were isolated by sarkosyl extraction. 9 Protein samples were boiled for 5 minutes, separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) on 11.6% acrylamide–0.4% bisacrylamide–0.1% SDS mini gels, and visualized by Coomassie Blue staining.
Nucleotide sequence accession numbers
The aligned sequence of the ompK36 gene in Mkp4437 was submitted to the EMBL/GenBank/DDBJ sequence database (accession no. JX891627).
Results
Antimicrobial susceptibilities and molecular analysis for β-lactamase and OMP
The MICs of meropenem against Mkp4437 and Mkp4365, which were isolated from the same patient, were 32 and 0.5 mg/L, respectively. The MICs of ceftazidime against Mkp4437 and Mkp4365 were 128 and 256 mg/L. These strains contained the blaIMP-1, blaDHA-1, blaCTXM-14, blaTEM-1, and blaSHV-1 genes, which were confirmed by PCR and DNA sequencing. The addition of CLA, MPA, or APB individually did not affect the MICs of meropenem or ceftazidime, but the combination of both MPA and APB reduced the MICs of meropenem and ceftazidime against Mkp4437 by 16-fold and 64-fold, respectively (Table 2).
+, production; –, no production.
Klebsiella pneumoniae Mkp4437, high-level carbapenem-resistant clinical isolate; K. pneumoniae Mkp4365, clinical strain less sensitive to carbapenem than the wild-type strain ENkp8003; K. pneumoniae Lkp8012, Lkp8026, and Lkp8027, carbapenem-resistant clinical isolate; K. pneumoniae Lkp8012/p4437, Lkp8026/p4437, and Lkp8027/p4437, high-level carbapenem-resistant clinical isolate harboring blaIMP-1 from Mkp4437; Escherichia coli DH5α (supE44ΔlacU169[Φ80 lacZΔM15] hsdR17 recA1 gyrA96 thi-1 relA1)(maA-10); E. coli TF-11, ceftazidime-resistant DH5α transformant harboring blaIMP-1 from Mkp4437; E. coli TS-11, ceftazidime-resistant ML4905 conjugant harboring blaIMP-1 from Mkp4437; K. pneumoniae ENkp8003, susceptible clinical isolate.
CLA, 4 mg/
Mkp4437 and Mkp4365 were found to be genetically homologous by PFGE. SDS-PAGE was performed to determine the difference in OMPs. Outer membrane profiles of Mkp4437 revealed that Mkp4437 lacked an OMP with a molecular weight of 36 kDa that was present in Mkp4365 (Fig. 1).
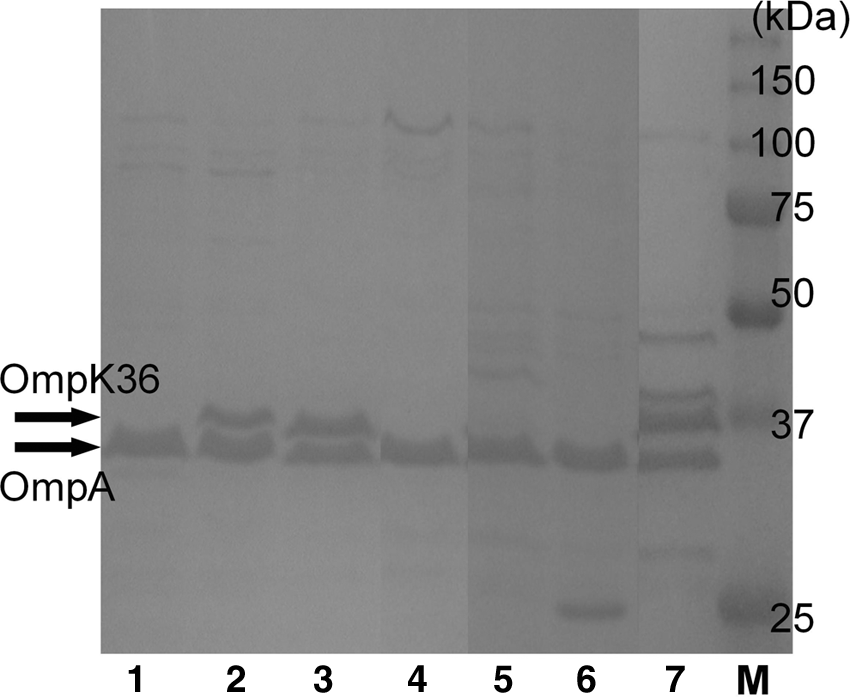
Outer membrane protein profiles of Klebsiella pneumoniae isolates. Lane 1, K. pneumoniae Mkp4437; Lane 2, K. pneumoniae Mkp4365; Lane 3, K. pneumoniae Lkp8001; Lane 4, K. pneumoniae Lkp8012; Lane 5, K. pneumoniae Lkp8026; Lane 6, K. pneumoniae Lkp8027; Lane 7, K. pneumoniae Ekp8003; Lane M, protein molecular mass standard (Bio-lad).
Class A, Class B, and Class C β-lactamases were detected by PCR, and Lkp8026 and Lkp8027 contained blaDHA-1 and blaSHV-1, and Lkp8001 contained blaIMP-1 and blaSHV-1. No broad-spectrum β-lactamase was found in Lkp8012 and ENkp8003. Only blaSHV-1 was found in these two isolates. The addition of CLA or MPA did not affect the MICs of meropenem and ceftazidime against Lkp8012, Lkp8026, and Lkp8027. The addition of APB reduced the meropenem and ceftazidime MICs by 16-fold to 32-fold in Lkp8026 and Lkp8027, respectively (Table 2). These three isolates also lacked an OMP with a molecular weight of 36 kDa that was present in Lkp4365, Lkp8001, and ENkp8003 (Fig. 1).
Antimicrobial susceptibility and analysis of plasmid
Ceftazidime-resistance was transferred successfully from K. pneumoniae Mkp4437 to E. coli DH5α by transformation (E. coli TF-11) and to E. coli ML4905 by conjugation (E. coli TS-11) to create a transformant and a transconjugant, respectively. The mobilization frequency from K. pneumoniae Mkp4437 to E. coli ML4905 was 4.32×10−5. A plasmid analysis of Mkp4437, Mkp4365, TF-11, and TS-11 demonstrated that they all carried a plasmid of ∼80 kbp, indicating acquisition of this plasmid by both the transformant and transconjugant. No plasmid was observed in E. coli DH5α and ML4905. The MICs of ceftazidime in TF-11 and TS-11 were elevated 256-fold and 512-fold compared with DH5α and ML4905, respectively. TF-11 and TS-11 were sensitive to meropenem, but the MICs against meropenem were elevated 4-fold to 8-fold compared with DH5α and ML4905, respectively. Meropenem resistance was transferred successfully from E. coli TS-11 to K. pneumoniae Lkp8012, Lkp8026, and Lkp8027 by conjugation (Lkp8012/pMkp4437, Lkp8026/p4437, and Lkp8027/p4437). PCR analysis confirmed that blaIMP-1 was transferred from Mkp4437 to TF-11, TS-11, Lkp8012/pMkp4437, Lkp8026/p4437, and Lkp8027/p4437, suggesting that blaIMP-1 was located on the 80-kbp plasmid. TF-11 and TS-11 were susceptible to aztreonam, which is stable in the presence of IMP-1 β-lactamase (Table 2).
Analysis of OMPs
The sequences of the ompK36 gene in Mkp4365 and Lkp8012 were 100% identical compared with that of the K. pneumoniae strain VM522. The ompK36 gene in Mkp4437 demonstrated that the insertion sequence of 618 bp was placed into the ompK36 gene at position 555 (Fig. 2). This insertion sequence was bound by 9-bp direct repeat sequences. Comparison of this insertional sequence with the sequences deposited in the GenBank database revealed 99.5% homology to 46.5% of insertion sequence IS10 (GenBank accession no. FJ829469). Using the Basic Local Alignment Search Tool, the insertion sequence was 618 bp of IS10, which had a 712-bp deletion between imperfect direct-repeat sequences (ACACGACTC and ACACGGACTC) of IS10. Direct sequencing of the PCR products demonstrated a single-amino-acid mutation in Lkp8026 (valine to proline) and Lkp8027 (valine to proline). PCR experiments also detected the ompK35 gene in Mkp4437 and Mkp4365. Direct sequencing of the PCR products demonstrated a premature stop codon at position 133 (tryptophan to stop codon) in the ompK35 gene in both isolates.
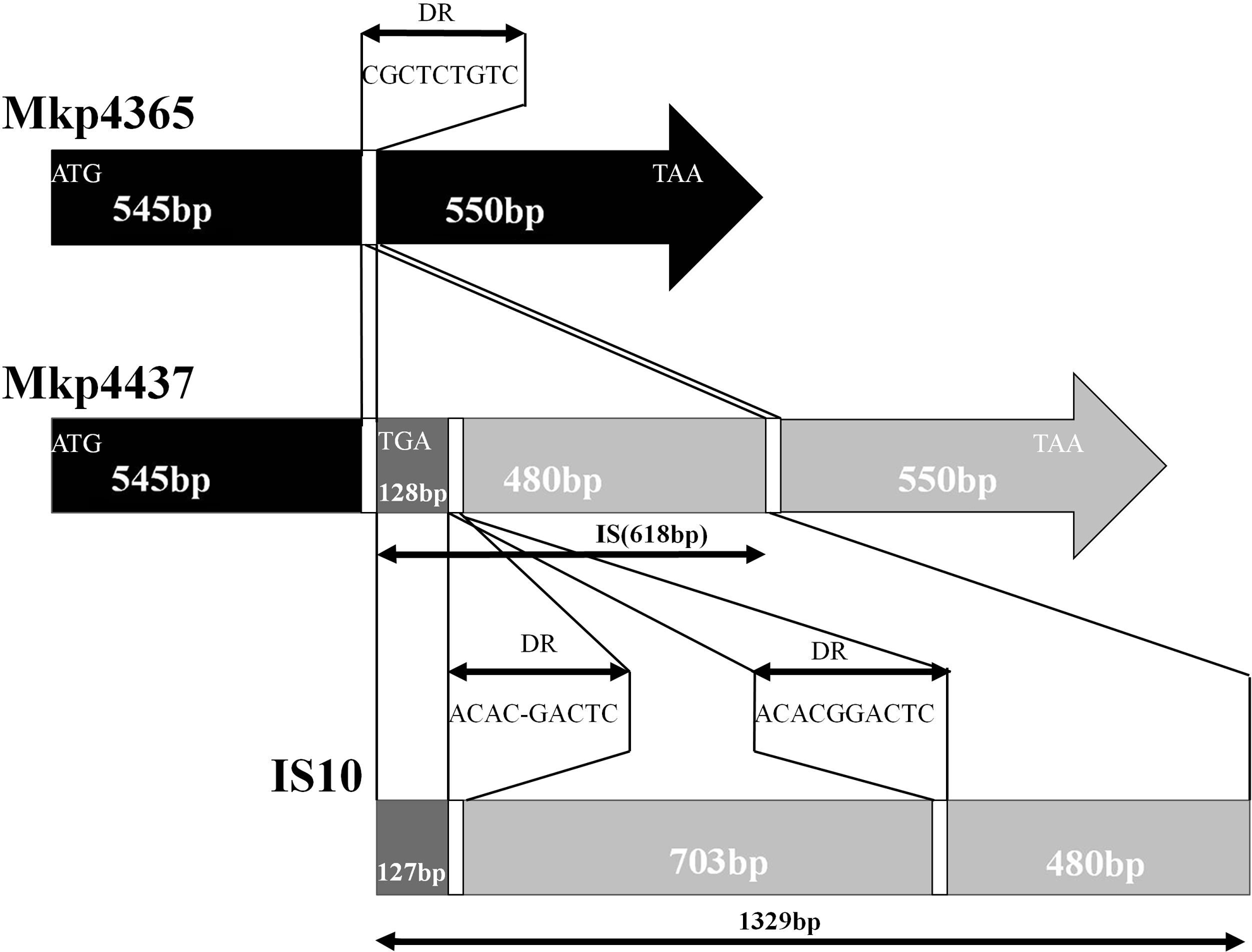
Genetic organization of the insertion element (819 bp) disrupting the ompK36 gene in the K. pneumoniae isolate, Mkp4437. The transcription directions of the ompK36 gene are depicted as black arrows. IS, insertion sequences; DR, direct repeat sequences.
Discussion
In this study, we discovered a high-level carbapenem-resistant K. pneumoniae strain, Mkp4437, and a less carbapenem-sensitive strain, Mkp4365. These strains were isolated from urine specimens at different time points in the clinical course of the same patient. Mkp4365 was isolated first, and Mkp4437 was isolated after treatment with doripenem for acute pyelonephritis. Despite the difference in carbapenem resistance, identical PFGE patterns were observed in both isolates. Both Mkp4437 and Mkp4365 contained blaIMP-1, blaDHA-1, blaCTXM-14, blaTEM-1, and blaSHV-1. The observed difference between the two strains was a lack of the OmpK36 porin protein in Mkp4437 that was present in Mkp4365. This difference alone resulted in a 32-fold to 64-fold increase in the MICs for the carbapenems. The OmpK35 porin protein was not expressed in either isolate by SDS-PAGE and sequence analysis. This result was consistent with the observations of Hernandez-Alles et al. that most ESBL-producing K. pneumoniae strains produce only the OmpK36 porin protein and not the OmpK35 porin protein. 9
IMP-1 β-lactamase is now reported to be the most epidemic carbapenemase of gram-negative bacilli in Japan, 43 but it is difficult to explain the mechanism of high-level carbapenem resistance by the production of IMP-1 alone. The MICs of meropenem against Mkp4365, Lkp8001, TS-11, or TF-11, which carry blaIMP-1, were 0.06–1 mg/L, demonstrating that IMP-1 β-lactamase production is insufficient for high-level carbapenem resistance in K. pneumoniae or E. coli. K. pneumoniae isolates carrying blaIMP-1 and displaying high-level carbapenem resistance have been identified in two Japanese reports,7,33 but OMPs were not investigated.
After screening for the isolates with MIC ≥1 mg/L to imipenem stocked between 2008 and 2011 in our University, we found three clinical K. pneumoniae strains, Lkp8012, Lkp8026, and Lkp8027, which had MICs for imipenem of 2–4 mg/L. Interestingly, these three strains lacked OmpK36, but they did not contain blaIMP-1. The clinical strain Lkp8012 encoded only SHV-1 β-lactamase and did not encode broad-spectrum β-lactamase. The clinical strains Lkp8026 and 8027 encoded the DHA-1 and SHV-1 β-lactamase, but did not express OmpK36, and were less susceptible to imipenem (MIC 2 μg/ml). Only when the IMP-1 β-lactamase is present in combination with the loss of Ompk36, where the MICs to imipenem ≥16 μg/ml. Ten other carbapenem-sensitive clinical strains (MIC for imipenem of ≤0.5 mg/L) were analyzed as controls, and all of these sensitive strains contained OmpK36. A lack of OmpK36 was related to carbapenem resistance, but it was insufficient for high-level carbapenem resistance.
To obtain a new experimental OMP-deficient strain that harbored blaIMP-1 as Mkp4437 does, a conjugation experiment was performed using the transconjugant E. coli TS-11 harboring blaIMP-1 from Mkp4437 as the donor strain, and Lkp8012, Lkp8026, and Lkp8027 as the recipient strains. By utilizing TS-11 instead of Mkp4437, we could select the transconjugants cultured on CHROMagar Orientation plates by the colors of the colonies. The plasmid was transferred successfully to OmpK36-deficient K. pneumoniae recipient strains, Lkp8012, Lkp8026, and Lkp8027, by this conjugation experiment. These transconjugants, Lkp8012/p4437, Lkp8026/p4437, and Lkp8027/p4437, displayed a high-level carbapenem resistance. A high MIC for meropenem (MIC 32 mg/L) with addition of APB was also observed in Mkp4437, Lkp8026/p4437, and Lkp8027/p4437, which had the production of DHA-1 β-lactamase in addition to IMP-1 β-lactamase. It became clear that both the production of IMP-1 β-lactamase and the lack of OmpK36 were the mechanisms of high-level carbapenem resistance. DHA-1 had some activity to elevate the MIC to carbapenem. However, additional IMP-1 induced high-level carbapenem resistance by a conjugation study. We believe that this article is the first report of the transmission of a plasmid harboring blaIMP-1 by conjugation into clinical OmpK36-deficient K. pneumoniae strains. Higher resistance to carbapenem was observed in Lkp8012/p4437 compared with the donor strain Mkp4437. This result suggests the another mechanism, such as duplicated IMP-1 genes or unknown resistant mechanisms, and further analysis is needed to confirm this hypothesis. The frequency of OmpK36-deficient K. pneumoniae strains in our university was rare (0.1%; 3 out of 3,151 isolates), but it is vital to pay attention to the presence of these isolates because of their potential to acquire the capacity to produce IMP-1 β-lactamase, which would lead to a high-level carbapenem-resistant phenotype. OmpK36-deficient K. pneumoniae strains were investigated for ESBLs or AmpC β-lactamases, and a higher resistance to cephalosporin or carbapenem was observed,5,23 and high-level carbapenem resistance with KPC and OXA carbapenemase, and OmpK35/36 inactivation have been reported extensively worldwide.18,19,24,27–29,41 However, few reports have previously investigated whether OmpK36-deficient K. pneumoniae strains with the MBL, IMP-1, show high-level carbapenem resistance. In 1999, a high-level carbapenem-resistant K. pneumoniae strain harboring IMP-1 was reported in Singapore. 16 It was later reported that the lack of an OMP was the other factor in carbapenem resistance. 17 However, the MICs against carbapenems of OMP-deficient strains that lack the production of IMP-1 β-lactamase were not investigated, nor were the possible reason for the lack of an OMP.
A unique genomic change was observed in Mkp4437, Lkp8026, and Lkp8027 by direct sequencing of the PCR products of the ompK36 gene. An additional insertion sequence resulted in a premature stop codon in the ompK36 gene of Mkp4437 that lacked the OmpK36 porin protein. The insertional inactivation of the ompK36 gene has been described previously.20,37 Mkp4437, however, showed differences in the additional DNA sequence and in the insertion positions relative to these strains. The insertion sequence was a part of IS10, and a 712-bp deletion was found in the transposase gene. This deletion, which was found in IS10, may deactivate or lower the activity of the IS10 transposase, 15 and increase the stability of the inactivated ompK36 gene of Mkp4437. Sequence analysis of the ompK36 gene of Lkp8026 and Lkp8027 showed a single-amino-acid mutation (valine to proline). Lkp8012 did not show any mutation in the amino acids of the ompK36 gene, suggesting that there might be a mutation in the promoter region. Further analysis is needed to confirm this hypothesis.
In conclusion, a high-level carbapenem-resistant K. pneumoniae strain coproducing IMP-1, DHA-1, CTXM-14, TEM-1, and SHV-1 β-lactamase was identified, and the mechanism of high-level carbapenem resistance in K. pneumoniae was clarified by an analysis of the plasmid and outer membrane. It is important to monitor for the presence of less carbapenem-sensitive K. pneumoniae strains that lack OmpK36, because blaIMP-1 transmission to these strains may lead to isolates with a high-level carbapenem-resistant phenotype. In future studies, it will be necessary to examine other unknown β-lactamase and OMP losses to identify other possible resistance mechanisms.
Footnotes
Acknowledgments
We thank H. Aono for excellent technical support. We are also very thankful to the members of the Hibiki Research Group for Clinical Microbiology who contributed to collection of the clinical isolates. Funding: None.
Disclosure Statement
None to declare.