
Abstract
Introduction
The aim of this study was to evaluate the in vitro activity of DOR alone and in combination with a variety of commonly used anti-Acinetobacter chemotherapeutic agents against MDR A. baumannii strains, including carbapenem-resistant strains, isolated during some outbreaks in central Italy.7,8 Part of this work has been presented at the 51th ICAAC meeting, Chicago (USA), 2011 (Poster E-736).
Materials and Methods
Bacterial isolates and epidemiological typing
As part of the surveillance activity of the network “Gruppo Romano Acinetobacter baumannii,” 22 A. baumannii isolates were selected from a well-characterized collection of A. baumannii strains obtained from January 2004 to December 2008 from ICU patients cared in eight general hospitals from central Italy (designated A, B, C, D, I, L, M, and P)7,8 (Table 1). Twenty-one A. baumannii isolates were obtained from clinical specimens of ICU patients, and one was recovered from the ICU environment (Table 1). None of the patients underwent previous treatment with DOR. The collection comprises an index strain (ACICU) from an ICU outbreak, whose complete genome has recently been sequenced. 13 The two prototypic strains for the epidemic international clonal lineages (ICLs) I (RUH875) and II (RUH134) were included as reference. 20 To analyze if synergistic interactions were mostly related to particular features of the strains, A. baumannii isolates were selected based on the hospital of origin (representing 8 hospitals), DOR susceptibility (5 sensitive and 17 resistant), molecular pulsotype, carbapenemase gene carried, and antibiotic resistance profile (Tables 1 and 2). Since there is no widely accepted definition for MDR A. baumannii, hereafter, we shall refer to the MDR phenotype as diminished susceptibility to ≥3 classes of antimicrobial agents. 10 The MDR phenotype of A. baumannii isolates of this study was previously described.7,8 Seventeen isolates were genetically related to either the ICL I (n=2) or II (n=15), and indicated as PFGE type 2 or 1, respectively. Typing data have been published elsewhere.7,8 In addition, five A. baumannii isolates (study codes #105, #122, #133, #138, and #215) showing a variant molecular type and not related to any ICL, were included. Isolates belonging to PFGE type 1 (except RUH134), and the strains 105 and 138 (characterized by a variant molecular type) were resistant to carbapenems, while those belonging to PFGE type 2 were susceptible (Table 2).
Breakpoint criteria were as follows. DOR: susceptible, ≤1 mg/L; resistant >1 mg/L, according to FDA; TIG: susceptible, ≤2 mg/L; intermediate, 4 mg/L; resistant, ≥8 mg/L according to FDA; COL: susceptible, ≤2 mg/L; resistant ≥4 mg/L5; AMK: susceptible, ≤16 mg/L; intermediate, 32 mg/L; resistant, ≥64 mg/L5; SAM: susceptible, ≤8/4 mg/L; intermediate, 16/8 mg/L; resistant, ≥32/16 mg/L5; RIF: susceptible, ≤1 mg/L; resistant, ≥2 mg/L.12
A. baumannii index strain 13 .
International clonal lineage I (RUH 875) and II (RUH 134) 20 .
DOR, doripenem; TIG, tigecycline; COL, colistin; AMK, amikacin; SAM, ampicillin/sulbactam; RIF, rifampicin; FDA, Food and Drug Administration; MIC, minimum inhibitory concentration.
Isolates are ranked in decreasing order of resistance.
Isolates showing an intermediate level of susceptibility were classified as resistant.
The synergistic activities are shaded in gray.
R, resistant to; S, sensitive to; In, indifference.
Antimicrobial agents and MIC assays
Antimicrobial agents were the following: DOR, TIG, COL, AMK, ampicillin/sulbactam, and rifampicin. MIC determinations for all antibiotics were performed by the broth microdilution method, according to the Clinical and Laboratory Standards Institute (CLSI) protocol. 4 Powders were obtained from the Sigma-Aldrich (Milan, Italy), except DOR (Ortho-McNeil Pharmaceutical Corporation, Raritan, NJ). MICs were determined in 96-well microtiter plates (Costar, Cambridge, MA) containing a freshly prepared Mueller-Hinton broth (Oxoid, Milan, Italy). The inoculum was adjusted to ∼5×105 CFU/ml in a 100-μl final volume, and microtiter plates were visually read after incubation for 24 hr at 37°C. Escherichia coli ATCC 25922 and Staphylococcus aureus ATCC 29213 were used as internal quality control strains. No CLSI breakpoints were available for DOR, thus, a susceptibility breakpoint of 1 mg/L was adopted according to the US Food and Drug Administration (FDA). The US FDA breakpoints approved for Enterobacteriaceae were also applied to define TIG susceptibility (susceptibility, ≤2 mg/L; resistance, ≥8 mg/L). Breakpoints for rifampicin were interpreted according to Hogg et al. 12 Other MIC results were interpreted according to the CLSI breakpoint criteria. 5
Chequerboard assays
Antibiotic interactions were determined using the chequerboard assay as previously described. 23 The range of drug concentrations used in the chequerboard analysis was such that the dilution range encompassed the MIC for each drug used in the analysis. Broth microdilution plates were inoculated with each A. baumannii isolate to yield ∼5×105 CFU/ml in a 100-μl final volume, and incubated for 18 hr at 37°C.
Synergy was defined as requiring a fourfold reduction in the MIC of both antibiotics in combination, compared with each used alone, measuring the fractional inhibitory concentration index (FICI). The FICI was calculated for each combination using the following formula: FICI=FICA+FICB, where FICA=MIC of drug A in combination/MIC of drug A alone, and FICB=MIC of drug B in combination/MIC of drug B alone. The FICI was interpreted as follows: synergy, FICI≤0.5; indifference, 0.5<FICI≤4; antagonism, FICI>4. 23
Time-kill assays
Tubes containing the freshly prepared Mueller-Hinton broth supplemented with the drug were inoculated with A. baumannii isolates to a density of ∼5×105 CFU/ml in a final volume of 10 ml and incubated in a shaking bath at 37°C. Aliquots were removed at time 0, 3, 6, and 24 hr postinoculation, and serially diluted in saline for determination of viable counts. Diluted samples (100 μl) were plated on Mueller-Hinton agar plates and bacterial counts were determined after an 18-hr incubation at 37°C. The antibiotic concentrations used in time-kill assays corresponded to 0.5-, 1-, and 2-fold the MIC values in combination as determined by the chequerboard method, that is, 4- to 16-fold lower than the MIC of each antibiotic alone. The bactericidal activity was defined as ≥3 log10 CFU/ml reduction in the colony count relative to the initial inoculum. 27 Synergy was interpreted as ≥2 log10 decrease in CFU/ml by the drug combination when compared with its most active constituent, and ≥2 log10 decrease in the CFU/ml below the initial inoculum, at any time point. The drug combination was considered to be antagonistic for ≥2 log10 increase in CFU/ml and indifferent for <2 log10 change in CFU/ml. 23 All synergistic interactions were confirmed by triplicate assays. Only antibiotic combinations showing synergism or antagonism in both chequerboard and time-kill assays were accepted as authentic synergistic or antagonistic interactions, respectively.
Results
The antibiotic susceptibility levels, expressed as MIC of DOR, TIG, COL, AMK, ampicillin/sulbactam, and rifampicin were preliminarily determined for the whole panel of 24 A. baumannii isolates (Table 1). All isolates, except the isolate 50, were susceptible to COL (95.9%). A high number of isolates were resistant to rifampicin (n=20, 83.3%), DOR (n=17, 70.8%), ampicillin/sulbactam (n=17, 70.8%), and AMK (n=15, 62.5%), while only nine isolates (37.5%) were resistant to TIG. Reference strains RUH 875 and RUH 134, isolated in early 1980s, showed an overall susceptible profile (Tables 1 and 2).
Chequerboard analysis performed with all antimicrobials in combination with DOR showed synergy in 13 A. baumannii isolates (54.2%) (Table 2). Moreover, seven isolates (29.2%) showed ≥2 synergistic interactions (Table 2). DOR showed a synergistic activity in combination with TIG (eight strains; 33.3%), COL (eight strains; 33.3%), AMK (four strains; 16.7%), ampicillin/sulbactam (two strains; 8.3%), and rifampicin (one strain; 4.2%) (Table 2). No antagonistic interactions were observed. Considering only DOR-resistant strains, synergistic interactions were detected in 76.5% of isolates (Table 2). Remarkably, synergistic effects were detected only in DOR nonsusceptible strains (Table 2).
All synergistic interactions inferred from chequerboard analysis were reassessed by time-kill kinetic experiments. Time-kill diagrams for effective combinations are shown in Fig. 1. Time-kill assays confirmed synergistic interactions in eight isolates (giving 10 synergistic interactions) for DOR in combination with TIG (n=4), COL (n=5), and AMK (n=1). The concentration of individual drugs in synergistic combinations is shown in Table 3. Synergistic effects were observed at 6 hr for DOR/TIG, at 24 hr for DOR/COL and DOR/AMK combinations. Synergistic antibiotic concentrations in time-kill assays resulted in 4- to 16-fold lower than the MIC of each antibiotic alone (Table 3 and Fig. 1). No synergistic combination resulted in a bactericidal activity. Moreover, no antagonistic interactions were observed with time-kill assays.
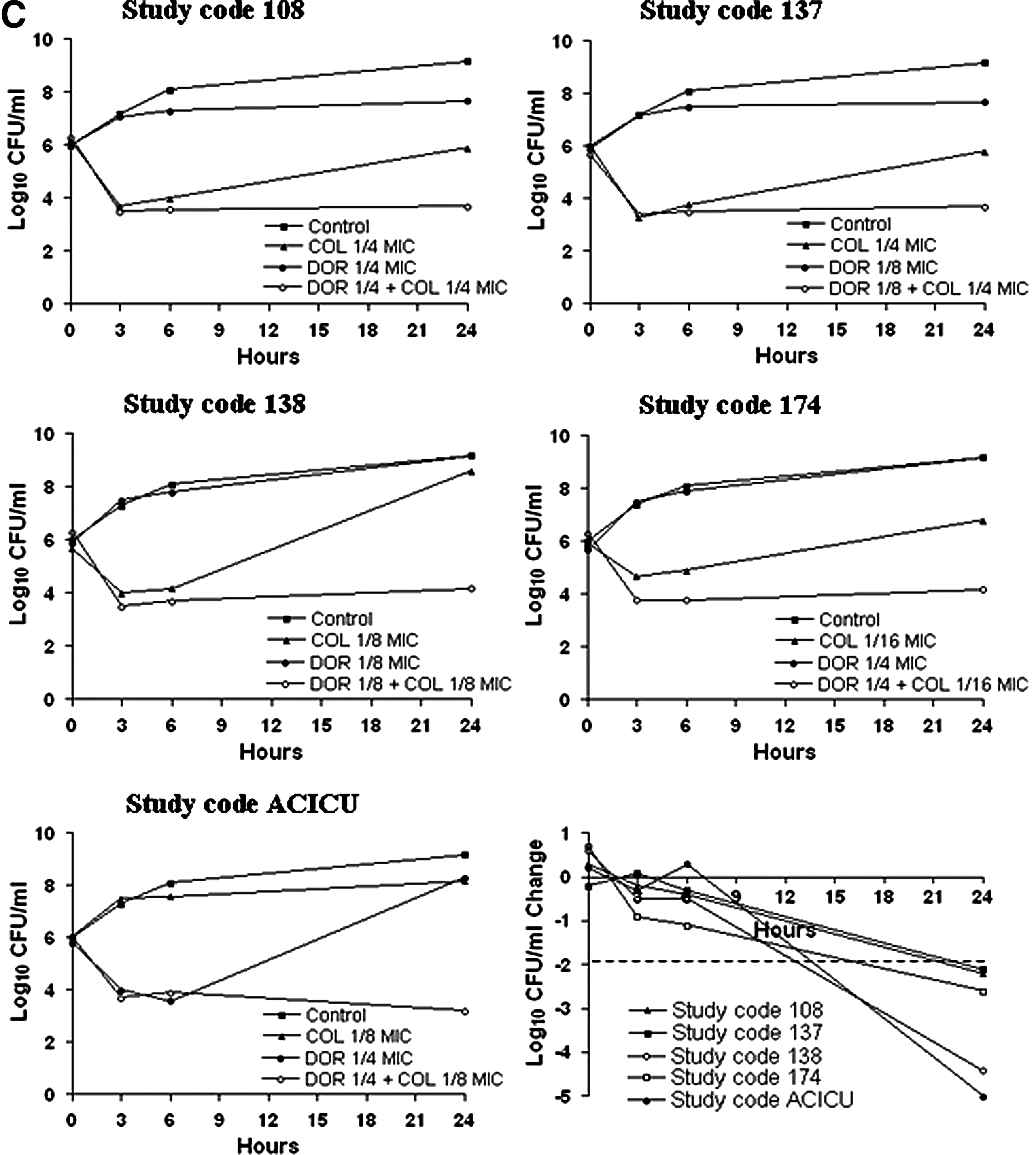
Time-kill kinetics and comparison of quantitative change in CFU/ml relative to the most active constituent, for the synergistic interactions:
Breakpoint criteria are provided in Table 1.
MDR, multidrug resistant; NS, not synergistic.
Discussion
A. baumannii is currently resistant to multiple antibacterial agents, including carbapenems, and occasionally COL and TIG. The ability to survive in the environment during prolonged periods of time, combined with its innate resistance to desiccation and disinfectants, makes A. baumannii almost difficult to eradicate from the clinical setting. For critically ill patients with MDR A baumannii infection, therapeutic options are limited and a combination treatment is advisable. In this work, we investigated DOR interactions with various antimicrobials by a two-step approach, involving preliminary chequerboard screening and subsequent time-kill assays. The chequerboard is an easy to perform, high-throughput method, which provides a single time point evidence of bacterial growth inhibition, and generally results in an overestimate of synergistic interactions. 23 For these reasons, all effective combinations inferred from chequerboard analysis were reassessed by time-kill assays. Although time-consuming and cumbersome, the time-kill assays provide a dynamic picture of antibiotic action over time. 9 Hence, only combinations showing synergy in both assays were interpreted as authentic synergistic interactions.
Chequerboard screening provided synergistic results for DOR in combination with TIG, COL, AMK, ampicillin/sulbactam, and rifampicin in 13 out of 24 isolates, with 7 isolates showing ≥2 synergistic interactions. Time-kill kinetics confirmed synergism in eight MDR isolates, giving a total of 10 synergistic interactions, only for the combinations DOR/TIG (n=4), DOR/COL (n=5), and DOR/AMK (n=1).
The DOR/TIG combinations showed synergistic interactions in time-kill assays at 6 hr of incubation period. All, but one, TIG synergistic concentrations exceed the maximum plasmatic concentration of TIG (0.38 mg/L) achievable with a standard dosage. 18 However, thanks to its pharmacodynamic properties, TIG is rapidly distributed into some tissues resulting in higher tissue concentrations as compared to plasma. 22 The concentrations of DOR used in our study were below the maximum plasmatic concentration achievable with a standard dosage. 1 Remarkably, three isolates showed synergistic interactions with TIG in combination with levofloxacin (study codes #5 and #11) and imipenem (study code #80), as previously described, 25 showing a particular trend to develop synergistic interactions based on TIG in these TIG nonsusceptible strains. These considerations suggest a potential clinical usefulness for the DOR/TIG combination. Notably, we described for the first time the in vitro synergistic interaction between these two antibiotics.
The DOR/COL combinations showed synergistic interactions in time-kill assays at 24 hr of incubation period. All concentrations of DOR and COL in our study were below the maximum serum concentrations achievable after a standard dosing regimen.1,6 In vitro synergistic interactions between DOR and COL in A. baumannii were previously described only by Pankuch et al., showing a maximum synergistic effect at 24 hr of incubation period in agreement with our findings. 21 COL causes permeabilization of the bacterial outer membrane, which would allow enhanced penetration by and activity of the other antibiotics in combination. The DOR/COL synergistic interaction could therefore have an impact in clinical practice by reducing the therapeutic dosage of COL, and hence, the risk of collateral effects, which currently represents a major limitation to its clinical use. 24
We identified one isolate, the epidemic index strain ACICU, showing the TIG/AMK synergistic interaction at 24 hr (4 and 16 mg/L for DOR and AMK, respectively). The concentrations of DOR and AMK were below the maximum serum concentrations achievable after a standard dosing regimen.1,6 In vitro synergistic interactions between DOR and AMK in A. baumannii were previously described by Pankuch et al., showing a maximum synergistic effect at 24 hr of incubation period in agreement with our findings. 21
In all, but four cases, strains became susceptible to DOR (MIC in combination ≤1 mg/L; see Table 3) in synergistic interactions. In the remaining four cases, synergistic combinations contributed only to reduce the MIC values, but strains remained resistant to DOR (MIC in combination >1 mg/L; see Table 3).
No antagonism or bactericidal activity was identified in time-kill assays. Notably, all synergistic interactions were identified in blaOXA-58-positive A. baumannii strains resistant to DOR. No synergistic interactions were found in blaOXA-23- or blaOXA-24-positive strains in time-kill assays. These findings demonstrated the major activity of DOR against blaOXA-58-positive A. baumannii strains, giving lower MIC values for DOR than blaOXA-23- or blaOXA-24-positive strains, although resulting in nonsusceptible profiles (see Tables 1 and 2).
The molecular mechanisms of synergy between DOR and the various antibiotics deserve further investigation. Synergistic interactions were observed only for PFGE type 1, demonstrating a largely lineage-specific trend, even though the number of isolates belonging to PFGE type 2 was very limited. The different synergistic activities observed in A. baumannii isolates sharing the same epidemiological type probably reflect the variable expression of different resistance determinants. This poses the need to test synergistic interactions even in the case of clonal isolates characterized by identical genetic fingerprints and resistance profiles. Synergy tests should be performed before starting any combined therapy. Unfortunately, the chequerboard method and time-kill assay are not applicable in a laboratory routine, being time-consuming and cumbersome, so synergy tests were applied only in selected cases.
Resistance to DOR in A. baumannii epidemic strains is a serious cause of concern, but in vitro synergistic solutions with other antibiotics could refresh its clinical usefulness against these microorganisms. However, as a limitation, in vitro synergy tests alone are not able to predict a successful in vivo response to treatment. Further studies are needed to elucidate the molecular mechanisms responsible for synergistic interactions with DOR and to explore their therapeutic potential. It will also be necessary to combine in vitro findings with additional pharmacokinetic and pharmacodynamic data to provide a more meaningful prediction of the in vivo efficacy of synergistic combinations in clinical practice. If confirmed, our findings could represent a possible option to be clinically evaluated in a combination therapy for treating serious infections caused by MDR A. baumannii.
Footnotes
Acknowledgments
We thank Francesco Luzzaro for critical reading of the manuscript. Ortho-McNeil Pharmaceutical Corporation (Raritan, NJ, USA) kindly provided laboratory-grade DOR. This study was partially supported by the funding of the Ministry of Health (RF2009-1526402).
Disclosure Statement
Nicola Petrosillo received honoraria as speaker for Pfizer, Astellas, Sanofi Aventis, Wyeth, Glaxo SmithKline, Merck Sharp & Dohme, Novartis, Carefusion, Johnson & Johnson, Janssen Cilag, and Bristol Myers Squibb.