
Abstract
A panel of 29 multidrug-resistant (MDR) Pseudomonas aeruginosa isolates recovered from seven hospitals as part of a country-wide surveillance of antimicrobial resistance in Bulgarian hospitals was studied. Molecular typing through multiple-locus variable number tandem-repeat analysis (MLVA6) yielded 23 different profiles. Phenotypic and genotypic tests for the detection of acquired carbapenemases yielded negative results in all cases. In contrast, 76% of the isolates produced other acquired β-lactamases, including extended-spectrum β-lactamases (ESBLs). Namely, 6 of the isolates (21%) produced a VEB-1 ESBL; 14 (48%) produced an OXA-10-type enzyme (7 OXA-10 and 7 OXA-10 ESBL variants, including 2 OXA-17 [A218G], 2 OXA-74 [C197T, A218G], and 3 OXA-142 [A218G, G470A]); 8 (28%) an OXA-2-type enzyme (all OXA-2); and 1 (3%) a PSE-1 carbenicillinase. Further analysis through multilocus sequence typing (MLST) revealed that the six VEB-1-producing strains, recovered from four hospitals, belonged to ST111 or ST244 international high-risk clones. Additionally, nearly all of the isolates (97%) lacked OprD production, explaining carbapenem resistance. Overexpression of AmpC was documented in 5 (17%) of the isolates, including most of the MDR isolates not producing any acquired β-lactamase. Particularly noteworthy was the very high prevalence of MexXY-OprM overexpression, documented in 72% of the isolates, whereas the prevalence of MexAB-OprM overexpression was lower (21%). In summary, while the production of metallo-β-lactamases is uncommon among P. aeruginosa isolates from Bulgarian hospitals, MDR profiles frequently result from the production of ESBLs combined with the lack of production of the carbapenem porin OprD and the overexpression of the MexXY-OprM efflux pump.
Introduction
Among the mutation-mediated resistance mechanisms are particularly noteworthy those leading to an alteration of the carbapenem porin OprD, the hyperproduction of the chromosomal cephalosporinase AmpC or the upregulation of one of the several efflux pumps.3,13,29 Further, the accumulation of these chromosomal mutations can lead to the emergence of MDR strains that may eventually be responsible for notable outbreaks in the hospital setting. 7 Likewise, multiple reports over the last decade have warned on the epidemic dissemination of the MDR strains producing transferable resistance mechanisms in multiple hospitals. Moreover, recent reports have provided evidence of the existence of MDR clones disseminated in several hospitals worldwide, denominated high-risk clones. 36 Among them, ST235, ST111, and ST175 are likely those more widespread.2,4,8–10,20,31 In this work, we describe the first multicenter study aimed to deciphering the molecular epidemiology and resistance mechanisms of MDR P. aeruginosa from Bulgarian hospitals.
Materials and Methods
Bacterial strains and susceptibility testing
A panel of 29 MDR P. aeruginosa isolates recovered from seven hospitals in the period 2004–2010 as a part of a continuous country-wide surveillance of antimicrobial resistance in Bulgarian hospitals was studied. All strains selected for the study were carbapenem nonsusceptible and met the recently established MDR consensus criteria: nonsusceptible to at least one agent in at least three antimicrobial classes, including antipseudomonal penicillins (ticarcillin or piperacillin), penicillin–β-lactamase inhibitor combinations (piperacillin–tazobactam), monobactams (aztreonam), carbapenems (imipenem or meropenem), fluoroquinolones (ciprofloxacin or levofloxacin), aminoglycosides (gentamicin, tobramycin, or amikacin), or polymyxins (colistin). 21 Identification and antimicrobial susceptibility testing were performed by conventional methods, including disk diffusion and VITEK2 (Bioéerieux). Minimal inhibitory concentrations to selected antimicrobials were confirmed by Etest (BioMérieux) and interpretated according to the EUCAST clinical breakpoints v 3.1 (www.eucast.org/clinical_breakpoints). Phenotypic assessment of potential carbapenemases was evaluated by Modified Hodge Test, 16 and MBL Etest (AB Biodisk), while the production of ESBLs was screened by double-disk synergy test using ceftazidime and amoxicillin–clavulanate in Muller-Hinton agar with and without 500 mg/L of cloxacillin.
Detection and sequencing of acquired antimicrobial resistance genes
Total bacterial DNA was isolated by the PrepMan
Characterization of mutation-driven antimicrobial resistance mechanisms
The levels of expression of ampC, mexB, and mexY were determined by real-time reverse transcription (RT)-PCR according to the previously described protocols.3,25 Briefly, strains were grown in 10 ml of LB broth at 37°C and 180 rpm to the late-log phase (optical density at 600 nm [OD600] of 1) and collected by centrifugation. Total RNA was isolated by using the RNeasy minikit (Qiagen), dissolved in water, and treated with 2 U of Turbo DNase (Ambion) for 30 min at 37°C to remove the contaminating DNA. The reaction was stopped by the addition of 5 μl of DNase inactivation reagent to the mixture. A 50 ng sample of purified RNA was then used for one-step RT and real-time PCR amplification using the QuantiTect SYBR green RT-PCR kit (Qiagen) with a SmartCycler II instrument (Cepheid). Previously described primers3,25 were used for the amplification of ampC, mexB, mexY, and rpsL (used as a reference to normalize the relative amount of mRNA). Strains were considered positive for ampC or mexY overexpression when the corresponding mRNA level was at least 10-fold higher than that of PAO1, negative if lower than 5-fold, and borderline if between 5-fold and 10-fold. Strains were considered positive for mexB overexpression when the corresponding mRNA level was at least 3-fold higher than that of PAO1, negative if lower than 2-fold, and borderline if between 2-fold and 3-fold. All PCRs were performed in duplicate. The mean values (±standard deviations) of mRNA levels obtained in three independent duplicate experiments were considered. Previously characterized strains overexpressing these mechanisms were used as controls. 3 Outer membrane protein (OMP) profiles were analyzed by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and stained with Coomassie blue following the previously described protocols. 24 Obtained OprD profiles were compared with those of PAO1 and its OprD-deficient mutant.
Molecular typing
Multiple-locus variable number tandem-repeat analysis (MLVA6) comprising four moderately variable minisatellites and two highly variable microsatellites was performed for molecular typing of P. aeruginosa with some modifications. 26 The primers were separated into two multiplex PCRs, mPCR1 (0.4 μM of ms010, 0.14 μM of ms077, and 0.15 μM of ms172 primers) and mPCR2 (0.25 μM of ms061, 0.08 μM of ms127, and 0.26 μM of ms142 primers). The PCR mixture (25 μl) contained ∼25 ng DNA, 1× Gold PCR buffer, 1 U AmpliTaq Gold, 0.2 mM dNTP, 1.5 mM MgCl2, and 6% DMSO. Thermocycling was performed with initial denaturation 94°C; for 6 min, followed by 37 cycles of 96°C for 15 sec, annealing at 60°C for 40 sec, elongation at 69°C for 90 sec, and final extension at 69°C for 10 min. PCR products were separated on a QiAxcel automated capillary electrophoresis system (Qiagen) applying the m500 method. The electrophoresis patterns were analyzed with BioCalculator software v3.2 (Qiagen), and the sizes converted to tandem-repeat numbers were imported into BioNumerics v4.5 (Applied Maths) as a character dataset. The cluster analysis was performed using the UPGMA (unweighed pair group method with arithmetic mean) algorithm and the categorical (or Hamming's) distance. 18 Prevalent MLVA6 clones were also analyzed by MLST, through the amplification and sequencing of seven housekeeping genes (acsA, aroE, guaA, mutL, nuoD, ppsA, and trpE) following the previously established schemes. 6 The isolates were assigned a sequence type (ST) number according to the allelic profiles available in the MLST database (http://pubmlst.org/paeruginosa).
Results and Discussion
Overall susceptibility data and resistance profiles for the panel of 29 carbapenem-resistant MDR isolates studied are shown in Tables 1 and 2, respectively. All isolates were susceptible to colistin. Among β-lactams, susceptibility was highest for piperacillin–tazobactam (51.7%), followed by cefepime (21.1%) and ceftazidime (17.2%). Among aminoglycosides, susceptibility was highest for tobramycin (27.6%), while nearly all isolates were nonsusceptible to gentamicin, amikacin, and fluoroquinolones (Table 1). The most frequent resistance profiles included all tested antibiotics, except colistin (n=7) or colistin and piperacillin–tazobactam (n=7) (Table 2).
NA, not applicable; S, susceptible; I, intermediate; R, resistant.
TIC, ticarcillin; PTZ, piperacillin-tazobactam; ATM, aztreonam; CAZ, ceftazidime; FEP, cefepime; IMP, imipenem; MER, meropenem; CIP, ciprofloxacin; LEV, levofloxacin; GEN, gentamicin; TOB, tobramycin; AMK, amikacin.
Results for MLVA6 typing and the acquired and mutational resistance mechanisms detected in the 29 MDR isolates analyzed from seven Bulgarian hospitals are summarized in Fig. 1. A total of 23 different MLVA6 patterns were detected. Significant correlation was found between the MLVA6 genotype and the resistance gene content, suggesting clonal spread of the MDR isolates.
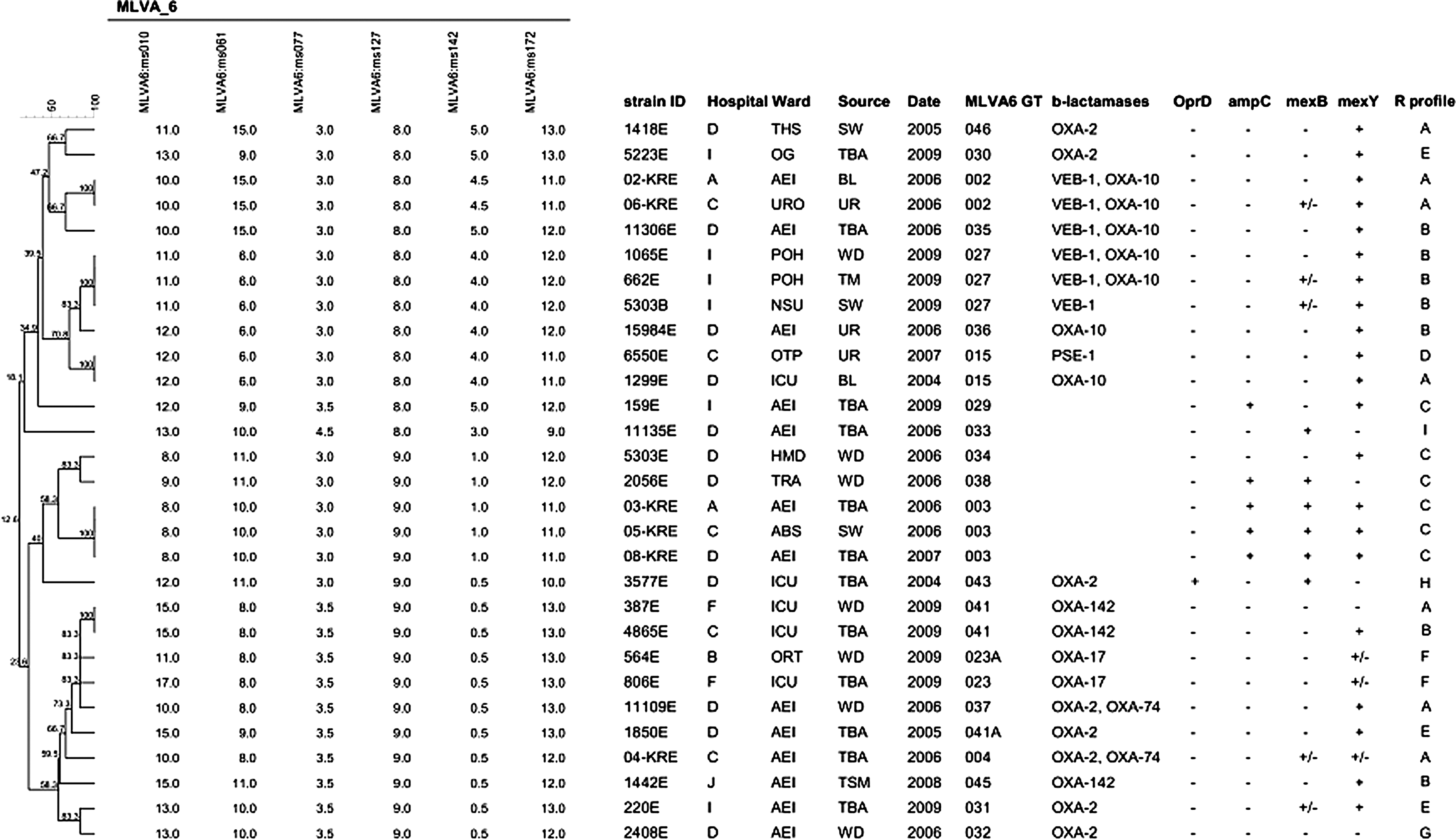
Dendrogram based on MLVA6 typing generated by applying the categorical coefficient and UPGMA clustering. The VNTR alleles are shown as repeat copy numbers. Hospital codes are as follows: A, plevem; B, Varna; C, Yambol; D, Sofia; F, Veliko tarnovo; I, Plovdiv; and J, Troqn. Hospital ward names were abbreviated as follows: ABS, Abdominal Surgery; AEI, Anesthesiology, Emergency and Intensive care; HMD, Hemodialysis; ICU, Intensive Care; NSU, Neurosurgery; OG, Obstetrics and Gynecology; ORT, Orthopedics and Traumatology; OTP, Outpatient; POH, Pediatric Oncohematology; THS, Thoracic Surgery; TRA, Traumatology; URO, Urology. The sources of the isolates were abbreviated as follows: BL, blood culture; SW, surgical wound; TBA, tracheobronchial aspirate; TM, Tap mixer; TSM, Tracheostoma; UR, Urine; WD, Wound. The codes used for resistance mechanisms are as follows: OprD, “+” presence and “−”absence in outer membrane protein (OMP) analysis through sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE); for ampC, mexB, and mexY, “+,” “+/−”, and “−” indicate overexpression, borderline overexpression, and no overexpression, respectively, according to the previously established criteria. 3 R profile stands for resistance profile according to the definitions provided in Table 2.
Phenotypic and genotypic tests for the detection of acquired carbapenemases yielded negative results in all cases. Thus, the growing incidence of MBL-producing P. aeruginosa observed in many European countries28–31,33 appears not to be yet a major threat in Bulgarian hospitals as opposed to neighboring Greece, where it is endemic. 28 In sharp contrast, up to 76% of the isolates were found to produce other acquired β-lactamases, mainly ESBLs. Namely, 6 of the isolates (21%) produced a VEB-1 ESBL; 14 (48%) produced an OXA-10-type enzyme (7 OXA-10 and 7 OXA-10 ESBL variants, including 2 OXA-17 [A218G], 2 OXA-74 [C197T, A218G], and 3 OXA-142 [A218G, G470A]); 8 (28%) an OXA-2-type enzyme (all OXA-2); and 1 (3%) a PSE-1 carbenicillinase. Figure 2 shows a map of the Bulgarian national territory indicating the acquired β-lactamases detected in each of the participating hospitals. Although several large outbreaks have been reported,9,35 the overall prevalence of ESBL-producing P. aeruginosa is found to be much lower in recent surveillance studies from other European countries.11,14
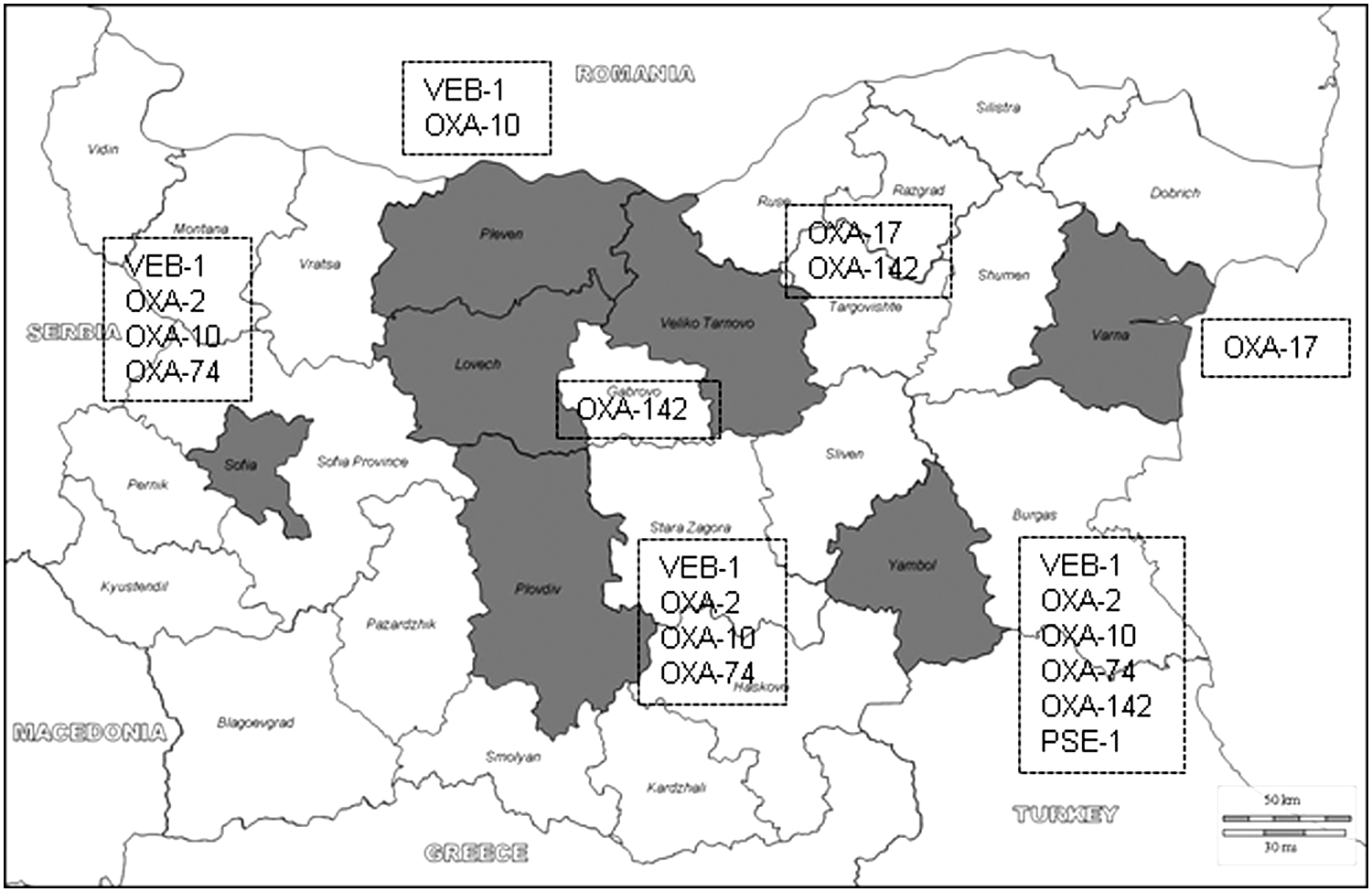
Map of the Bulgarian national territory (Daniel Dalet/d-maps.com) indicating the acquired β-lactamases detected in the hospitals from the seven districts participating in the study (shown in gray).
The detection of VEB-1-producing P. aeruginosa in four of the seven hospitals is particularly concerning. In this sense, our data extend the findings of a previous study showing a wide dissemination of VEB-1-producing P. aeruginosa in hospitals from Sofia not participating in the present multicenter study. 32 Among the six isolates producing this ESBL in the present work, three different MLVA6 clones were observed: 027 detected in three isolates from one of the hospitals (I), 002 detected in two isolates from two additional hospitals, and the closely related 035 found in 1 isolate from a fourth hospital (Fig. 1). Further typing through MLST, identified the MLVA6 027 clone as the international high-risk clone ST111, widely disseminated in Europe, and associated to the production of the MBL VIM-2.2,8,31,33 The analysis of an extended strain collection identified the VEB-1-producing MLVA6 027 (ST111) clone in 16 out of 21 MDR isolates from hospital I, indicating the epidemic dissemination of this strain. Thus, this work adds VEB-1, to the concerning transferable β-lactamases detected in the internationally spread high-risk clone ST111. Moreover, the closely related 002 and 035 MLVA6 clones were both found to belong to the ST244, previously detected in a PER-1 ESBL-producing strain causing an outbreak in a hospital from Poland, 9 and among colistin-resistant MDR P. aeruginosa isolates in Korea. 17 Therefore, ST244 should likely be added to the list of concerning high-risk clones. Moreover, our results are thus consistent with recent studies arguing for the usefulness of MLVA as a cost-effective alternative to MLST and PFGE for P. aeruginosa typing, due to the congruence between these methods. 34
The contribution of mutational resistance mechanisms to the MDR profiles of P. aeruginosa isolates from Bulgarian hospitals was also investigated (Fig. 1). All but one of the isolates (97%) lacked OprD production as documented by SDS-PAGE of OMPs (Fig. 3), explaining carbapenem resistance in these isolates. Our results are therefore consistent with previous data showing that in the absence of acquired carbapenemases, the inactivation of OprD is a nearly universal signature of imipenem resistance. 29 The single isolate showing wild-type OprD expression was an imipenem-susceptible meropenem intermediate isolate showing MexAB-OprM overexpression, known to affect meropenem, but not imipenem. 29 Additionally, overexpression of the chromosomal cephalosporinase AmpC was documented in 5 (17%) of the isolates, including most of the MDR isolates not producing any acquired β-lactamase. Thus, nearly all the studied MDR isolates either produced acquired β-lactamases or overexpressed the chromosomal cephalosporinase AmpC (Fig. 1).

OMP profiles of wild-type PAO1, its OprD-deficient mutant (PAOD), and six representative clinical isolates lacking OprD expression.
Regarding efflux pumps is particularly noteworthy the very high prevalence of MexXY-OprM overexpression, documented in up to 72% of the isolates (plus an additional 10% yielding borderline results), independently of the production or not of acquired β-lactamases (Fig. 1). As could be expected, all isolates showing MexXY-OprM overexpression were nonsusceptible to fluoroquinolones, gentamicin, and amikacin, and all except two to cefepime. Although MexXY-OprM has been previously shown to be the efflux pump most frequently overexpressed,3,13 such a high prevalence has not been previously reported outside of the cystic fibrosis chronic lung infection setting. In contrast, the prevalence of MexAB-OprM overexpression was considerably lower (21%) and was more frequent among those isolates not producing acquired enzymes (Fig. 1). As expected,3,13 the overexpression of this efflux pump was particularly frequent (45%) among meropenem-resistant isolates.
In summary, while the production of MBLs is uncommon among P. aeruginosa isolates from Bulgarian hospitals, the MDR profiles frequently result from the production of ESBLs combined with the lack of production of the carbapenem porin OprD and the overexpression of the MexXY-OprM efflux pump.
Footnotes
Acknowledgments
H. Hitkova, MD, Prof. I. Haydouchka, MD, B. Zaharieva, MD, A. Dimitrova, MD, V. Stefanova, MD, M. Grigorova, MD, and V. Donevska, MD are gratefully acknowledged for providing P. aeruginosa strains. This study was supported by the Ministerio de Economía y Competitividad of Spain, Instituto de Salud Carlos III, through the Spanish Network for the Research in Infectious Diseases (REIPI RD06/0008 and RD12/0015) and a Rio Hortega contract to XM and by the Direcció General d‘Universitats, Reserca i Transferència del Coneixement del Govern de les Illes Balears.
Disclosure Statement
None of the authors has conflicts of interest to declare.