
Abstract
Drug-resistance to ofloxacin (OFX) in Mycobacterium tuberculosis is due to missense mutations in gyrA and other factors, such as alterations in the activity of drug efflux pumps. In this study, we identified 8 extensively drug resistant tuberculosis (XDR-TB), 40 multidrug resistant TB (MDR-TB), 38 polydrug resistant TB (PDR-TB), and 16 single OFX-resistant TB from 102 clinical isolates. We tested the effect of three efflux inhibitors, reserpine, verapamil, and carbonyl cyanide m-chlorophenyl hydrazone (CCCP), on changes in the OFX minimum inhibitory concentration (MIC) using Resazurin microtitre assay. These three inhibitors changed the MICs from 2- to 32-fold, with CCCP having the strongest effect. A total of 55%, 74%, and 83% of the tested isolates had changes in MIC of more than two-fold by reserpine, verapamil, and CCCP, respectively. The inhibitors led to similar fold-changes of OFX MICs in the XDR, MDR, PDR, and single OFX-resistant isolates. For each inhibitor, a higher resistance to OFX was associated with the greater efflux pump activity. There were no significant differences in the effect of efflux pump inhibitors upon Beijing and non-Beijing M. tuberculosis genotypes. Taken together, these results indicate that the efflux pump activity was greater in the isolates higher resistant to OFX and had similar effects on isolates with different drug resistant pattern, and had similar effects on Beijing and non-Beijing genotypes.
Introduction
D
Resistance to FQs is believed to be due to mutations in the quinolone resistance-determining region (QRDR) of DNA gyrase (gyrA). The association of missense mutations in this region with FQ resistance is attributed to the important role of QRDR in the interaction of FQs and DNA gyrase.22,35 In clinical isolates, missense mutations within the QRDR of gyrA have been reported in the following six sites: Ala74, Gly88, Asp89, Ala90, Ser91 and Asp94.19,22,31
However, mutations in DNA gyrase alone do not account for resistance in a significant proportion of FQ-resistant M. tuberculosis isolates. This suggests other mechanisms that underlie the resistance, such as efflux pumps. 29 Accumulated evidence has implicated that efflux pumps may play a major role in intrinsic resistance. 25 In particular, increased expression of an efflux pump that exports FQs7,15 is involved in FQ resistance in laboratory-selected strains of M. tuberculosis. Several mycobacterial efflux pumps associated with FQ resistance have been described, including pumps of the Major Facilitator Super (MFS) family (lfrA, Rv1634 and Rv1258c) and ATP Binding Cassette (ABC) transporters (DrrAB, PstB and Rv2686c-2687c-2688c). 6 Previous research indicated that various efflux pump inhibitors, including MFS inhibitors (carbonyl cyanide m-chlorophenyl hydrazone [CCCP] and 2,4-dinitrophenol), an ABC transporter inhibitor (verapamil), and a Bmr efflux pump inhibitor (reserpine), increased the drug sensitivity of ofloxacin (OFX)-resistant M. tuberculosis isolates. 1
The interaction between the efflux pumps and point mutations in the QRDR was first reported in Escherichia coli, which demonstrated that early low-level levofloxacin resistance conferred by acrAB over-expression led to development of high-level resistance that was mediated by target site mutation(s). 28 Another study indicated that either mutation of efflux pumps or the point mutations in QRDR of the same M. tuberculosis clinical isolates resulted in large changes in the minimum inhibitory concentration (MIC). 27 In the present study, we investigated the effect of different point mutations of gyrA and the role of efflux pump inhibitors on resistance in M. tuberculosis. In particular, we analyzed the effects of certain efflux inhibitors on M. tuberculosis clinical isolates with different levels of resistance to OFX, including isolates characterized as XDR, MDR, polydrug resistant (PDR), and single OFX-resistant.
Materials and Methods
Strains and antibiotics
The control M. tuberculosis strain H37Rv (ATCC 27294) was obtained from the Chinese National Reference Laboratory. OFX-sensitive isolates were reported to have low or no response to efflux pump inhibitors,8,27 so the Lowenstein-Jensen (L-J) agar proportion method 31 was initially used to screen for OFX-resistant clinical isolates. A total of 102 OFX-resistant clinical isolates were obtained from 1,167 isolates from patients with TB in the Beijing Chest Hospital in 2009.
The following anti-TB drugs, purchased from Sigma-Aldrich (Beijing, China), were used for drug susceptibility test: isoniazid, rifampicin, streptomycin, ethambutol, OFX, AmK, capreomycin, and sodium para-aminosalicylate.
Drug susceptibility test
The drug spectrum was determined by the proportion method on L-J slants, with the M. tuberculosis H37Rv strain as the control.5,33 Briefly, 100 μL bacterial suspensions (∼10−2 mg/mL) were inoculated on drug-containing or drug-free L-J slants and incubated at 37°C for 4 weeks. Final readings were reported for drug-containing and drug-free slants.
To evaluate the efflux pump-mediated OFX resistance in M. tuberculosis isolates, MICs of OFX were determined using the Resazurin microtitre assay 20 in the presence or absence of efflux pump inhibitors (CCCP, reserpine, or verapamil; Sigma, St. Louis, MO).8,27,10 It was well known that both the resazurin and the efflux pump inhibitors might affect the proton motive force and thereby the metabolic/respiratory activity. To control this effect in this study we set up the positive control, negative control and the parallel MIC test with/without inhibitors for each tested isolate. The only difference in the parallel MIC test is the efflux pump inhibitor. Briefly, 100 μL of Middlebrook 7H9 broth with 0.05% Tween 80 and 10% OADC (Sigma) was dispensed into each well of a 96-well cell culture plate (Corning, Inc., Corning, NY) with various concentrations of OFX (0.5, 1, 2, 4, 8, 16, 32, and 64 mg/L). The recultured clinical isolates, without mutations in gyrA, were taken from L-J slopes and homogenized. The turbidity was adjusted to No. 1 McFarland standard (∼1×107 CFU/mL). The suspension was then serially diluted in Middlebrook 7H9 media and the final suspension (10 μL) was transferred to the prepared wells with 100 μL of each antimicrobial dilution. The final inoculum concentration was 5×104 CFU/mL per well. Plates were sealed and incubated at 37°C for 1 week. Twenty-five microlitres of 0.02% Resazurin (Sigma Chem. Co., St. Louis, MO) was added to each well and plates were reincubated for an additional 2 days. A change in color from blue to pink indicated the growth of bacteria. The MIC was defined as the minimum OFX concentration that prevented color change.
CCCP and reserpine are inhibitors of the proton motive force, 33 and verapamil blocks the calcium channels of ABC transporters. 2 Stock solutions of CCCP and reserpine were prepared in DMSO, and verapamil was dissolved in distilled water. The final concentrations of CCCP, verapamil, and reserpine used in Resazurin microtiter assays were 12, 60, and 8 mg/L, respectively.
Genomic DNA isolation, polymerase chain reaction and sequence analysis
The colonies were scraped, re-suspended in 500 μL of TE (10 mM Tris, 1 mM EDTA, pH 8.0), and killed by heating at 80°C for 30 min before extracting the DNA. The primers (CyberSyn Co., Beijing, China) and polymerase chain reaction (PCR) mixtures used for amplification were described previously. 19 A DNA fragment of 320 bp of the QRDR of gyrA was amplified using the following primers: 5′ CAG CTA CAT CGA CTA TGC GA 3′ and 5′ GGG CTT CGG TGT TAC CTC AT 3′, as described earlier. 19 The sequences of PCR products were confirmed using BLAST (www.ncbi.nih.gov/BLAST) and compared with that of the M. tuberculosis H37Rv strain using ClustalW multiple sequence alignment (www.ebi.ac.uk/Tools/clustalw2).
Molecular typing by spoligotyping
Spoligotyping was performed with a commercial kit (Isogen Bioscience BV, Maarssen, The Netherlands) according to the manufacturer's instructions. Amplification of the direct variant regions for spoligotyping was performed as described previously. 14 Interpretation of spoligotype patterns and assignment of octal codes were performed as previously described based on the SITVIT2 database (Pasteur Institute of Guadeloupe). This is an updated version of the SpolDB4 database (www.pasteur-guadeloupe.fr:8081/SITVITDemo). 13
Statistics
The Statistical Package for the Social Sciences (SPSS, Chicago, IL) for Windows was used for comparison of MICs by use of non-parametric statistical analysis (unpaired t-test, Gaussian Approximation). A p-value<0.05 was considered significant.
Results
Characteristics of OFX-resistant strains
We tested 102 OFX-resistant clinical isolates (MIC >2 mg/L on L-J agar) for resistance to nine anti-TB drugs by the agar proportion method. The results showed that 8, 40, 38, and 16 isolates were XDR, MDR, PDR, and single OFX-resistant, respectively (Table 1). The OFX-resistance levels were confirmed by the microtiter method with Middlebrook 7H9 plus 10% OADC. These results showed that 28 isolates had MICs of 16 mg/L or higher, 45 isolates had MICs of 8 or 4 mg/L, and 29 isolates had MICs of 2 mg/L or less (Table 1). We used this information as a basis to examine the association of OFX resistance with the response to efflux pump inhibitors.
Determined by agar proportion method.
Determined by microtiter method with Middlebrook 7H9 plus 10% OADC.
Single OFX, only resistant to ofloxacin; PDR, polydrug resistant, defined as resistant to two or more antibiotics but not MDR; MDR, multidrug resistant, defined as resistant to at least RIF and INH; XDR, extensively drug resistant, defined as resistant to at least RIF and INH among the first-line anti-TB drugs, resistant to a FQ, and resistant to at least one of the three injectable second-line drugs; MIC, minimum inhibitory concentration; RIF, rifampin; INH, isoniazid; FQ, fluoroquinolone; TB, tuberculosis.
Mutations in gyrA
Next, we identified the mutations in QRDR of gyrA in each isolate by DNA sequencing in triplicate. Mutations were present in 100% of the highly OFX-resistant isolates, 86.6% of isolates with MICs of 4–8 mg/L, and 82.8% of isolates with MICs of 2 mg/L or less (Table 2). The most frequent mutations were D94G and D94A (52.9% of tested isolates). One isolate had a G88C mutation. Double mutations were present in five isolates and the mutation patterns were A90V+S91P, A90V+D94G, and S91P+D94G. Comparison of the mutations of isolates that had MICs of 16 mg/L or greater with isolates that had MICs of 2 mg/L or less indicated only mutations of D94G and double mutations of A90V+S91P and A90V+D94G were related to OFX resistance. Eleven isolates had no mutations in the QRDR of gyrA, and most of these isolates had low MICs, with 5 of them having MICs of 2 mg/L or less. These results confirm that GyrA missense mutations play a role in the development of OFX resistance and that other factors might be involved in the resistance.
Nucleotide and residue numbers are based upon the H37Rv sequence (GenBank accession number L27294).
Mutation at Ser95 was not shown.
MICs of tested isolates without inhibition of the efflux pump activities (median, 25th to 75th centile).
CCCP, carbonyl cyanide m-chlorophenyl hydrazone.
Drug resistance spectra of isolates
We tested the inhibitory effects of the three efflux pump inhibitors on all 102 clinical isolates (Fig. 1). Efflux pump inhibitors altered the OFX MICs in 90.2% of isolates, and the changes of the MICs ranged from 2- to 32-fold. The change in MIC was more than two-fold in 55.9% of isolates in response to reserpine, in 74.5% of isolates in response to verapamil, and in 83.33% of isolates in response to CCCP. Based on the concentrations used in this study, CCCP had the greatest effect and reserpine had the weakest effect (Fig. 1A). Our data for those isolates with low-level resistance showed the evidence that they have low response to efflux pump inhibitor. Of all the 29 isolates with MICs not more than 2 mg/L by resazurin-based method only 5, 8 and 17 showed the fold changes with the inhibitors of reserpine, verapamil and CCCP, respectively.

Effect of reserpine, verapamil, and CCCP on the OFX MICs of Mycobacterium tuberculosis clinical isolates.
Five of the eight XDR-TB isolates had altered MICs in response to reserpine, verapamil, and CCCP (Fig. 1B–D). A total of 47.4%, 57.9%, and 86.8% of the MDR-TB isolates had a two-fold or greater change in the MIC due to reserpine, verapamil and CCCP, respectively. For PDR and single OFX-resistant isolates, the active efflux pumps were most affected by CCCP (85% and 68.8%) and least affected by reserpine (55% and 62.5%). The fold-change in the MICs induced by the three efflux pump inhibitors was similar in the XDR, MDR, PDR and single OFX-resistant isolates. In other words, the effect of these efflux pump inhibitors on OFX resistance was similar in isolates that had very different OFX resistant patterns.
Effect of efflux pump inhibitors on isolates with different levels of resistance to OFX
Analysis of the isolates by OFX-resistant levels indicated that for each inhibitor, isolates resistant to OFX with MICs of 16 mg/L or more were inhibited more than those with MICs not more than 2 mg/L and those with MICs of 4 and 8 mg/L. Only CCCP had a significant difference in the inhibition of isolates with MICs of 4 and 8 mg/L relative to those with MICs of 2 mg/L or less (p=0.006). There was no significant difference in reserpine or verapamil inhibition of isolates with MICs of 4 and 8 mg/L relative to those with MICs of 2 mg/L or less (Fig. 2A–C). The efflux pump inhibitors had less of an effect on the MICs in OFX-sensitive isolates (MIC ≤2 mg/L by microtiter method), with 14 of 29 isolates showing no change of MIC. Of the 11 isolates without mutation in QRDR, 5 with their efflux pump activity was inhibited by reserpine, 4 inhibited by verapamil and 8 inhibited by CCCP (Supplementary Data; Supplementary Data are available online at www.liebertpub.com/mdr).
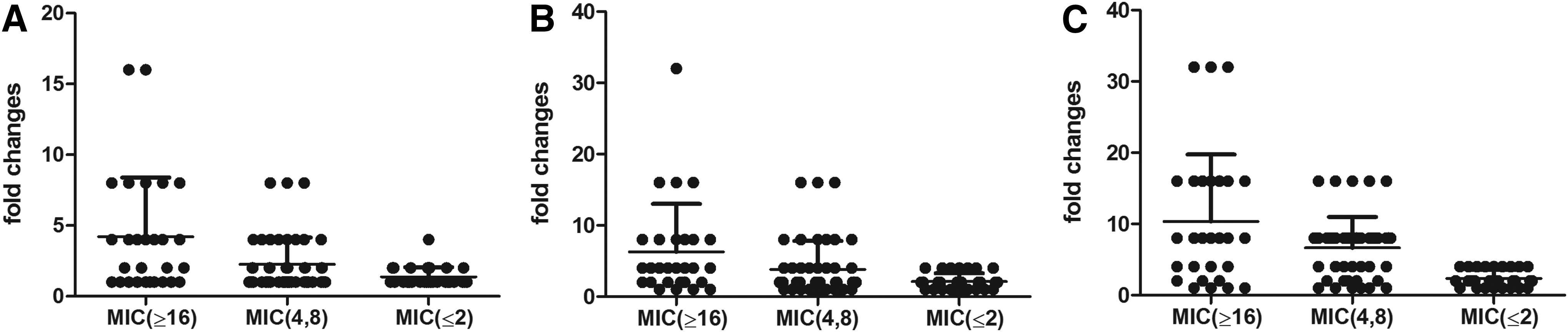
Effect of reserpine, verapamil, and CCCP on OFX MICs of M. tuberculosis clinical isolates.
Effects of efflux pump inhibitors on Beijing and non-Beijing M. tuberculosis genotypes
Finally, we analyzed the genotypes of the 102 M. tuberculosis clinical isolates by spoligotyping (Table 3). The results indicated that there were 11 spoligotypes, that 89 isolates (87.3%) were in the Beijing family, and that the spoligotype pattern of 000000000003771 was the most common (81 isolates). There were also 13 non-Beijing genotypes including those in the T1, T2, T5, H, and MANU2 families. Six of these isolates were in the T2 family. There was no significant difference in the MICs of isolates between Beijing and non-Beijing genotypes (p=0.5850) and also no special mutations correlated with one of the spoligotypes in this this study (Supplementary Data). In addition, the effect of the efflux pump inhibitors on OFX resistance was similar in isolates with Beijing and non-Beijing genotypes (Fig. 3).
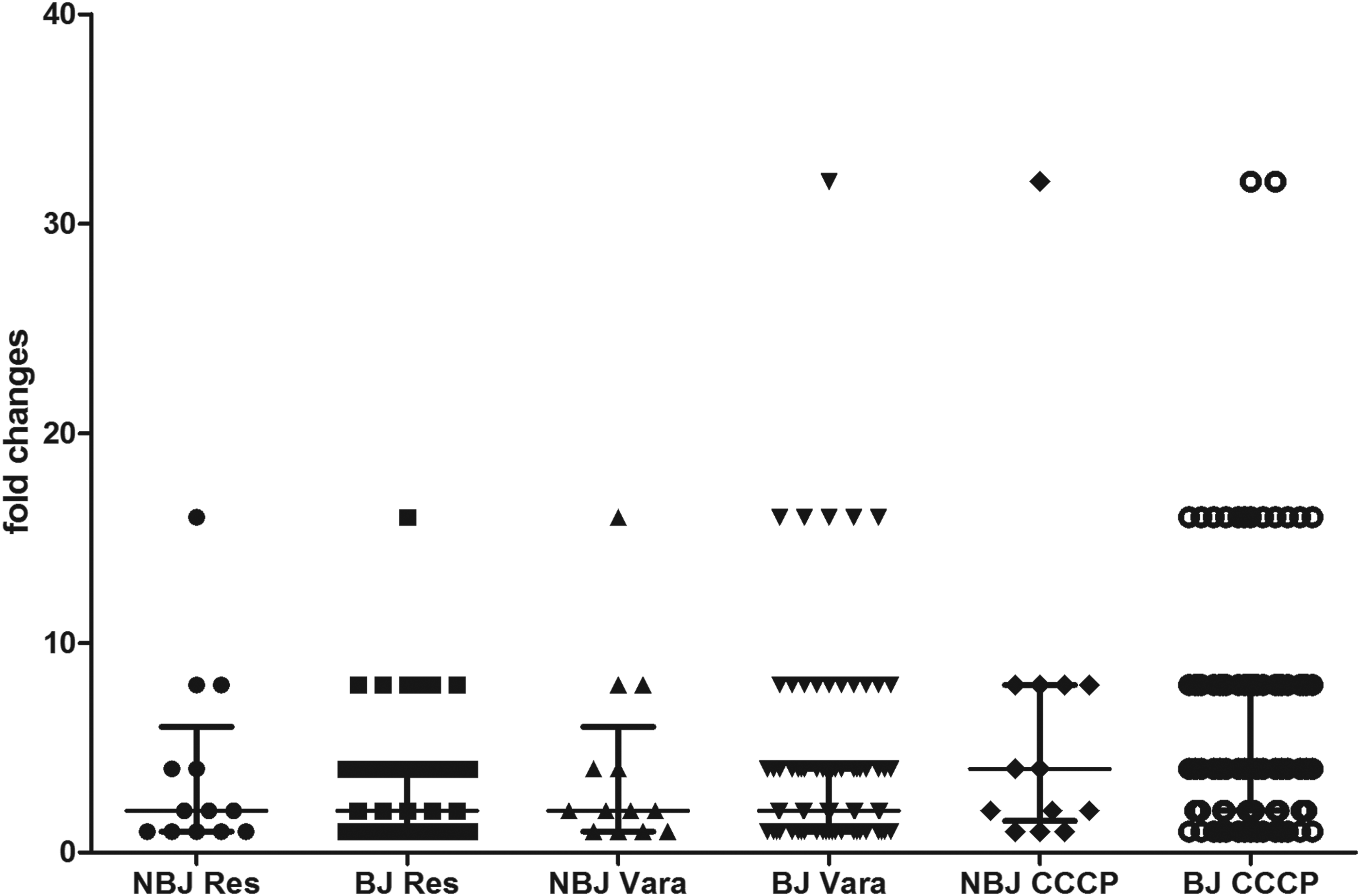
Comparison of changes in the OFX MICs of Mycobacterium tuberculosis clinical isolates of the Beijing genotype (BJ, n=89) and the non-Beijing genotype (NBJ, n=13) in the presence of CCCP, resperine (Res), and verapamil (Vera).
Discussion and Conclusion
A specific gene alteration (mutation, insertion, or deletion) can alter the structure of a drug's target protein and thereby influence the susceptibility of an organism to that drug. In particular, missense mutations in the QRDR of gyrA lead to resistance of M. tuberculosis to FQs.16,35 However, our previous data showed that resistance in some clinical isolates of M. tuberculosis cannot be explained by QRDR mutations. 22 Recent studies indicated that the MICs of most OFX-resistant M. tuberculosis isolates were affected by efflux pump inhibitors.23,27 In this study, we investigated the alteration of the OFX MICs by efflux pump inhibitors in isolates classified XDR, MDR, PDR and single drug resistant TB, in isolates with different OFX MICs, and in isolates with different genotypes.
Many research groups have reported the role of active efflux pumps in the development of OFX-resistance in M. tuberculosis clinical isolates.8,10,17,27 Our data confirms these previous results and also indicates that different efflux pump inhibitors had different effects on OFX sensitivity. In our study, efflux pump inhibitors affected the MICs in more than 90% of the 102 OFX-resistant clinical isolates (clinical isolates by the agar proportion method), compared to 66.6% of the isolates reported by Singh et al. 27 This difference may be due to the use of different concentrations of efflux pump inhibitors. In our study CCCP (12 mg/L) had a more potent effect than verapamil (60 mg/L) or reserpine (8 mg/L), while the inhibitory effect of CCCP (1 mg/L) reported by Singh et al. was not so strong as that of verapamil (5 mg/L). 27 It has also been reported that in the presence of reserpine, about 57%–100% M. tuberculosis isolates resistant to FQs (ciprofloxacin, moxifloxacin, levofloxacin, OFX, gatifloxacin) showed at least two-fold reduction in MIC. 8
The sensitivity to OFX can be affected by different efflux pump inhibitors, and by the intrinsic characteristics of the mycobacteria. Our results indicated no relationship between the drug resistance pattern (XDR, MDR, PDR, or single resistant TB) and the effect of efflux pump inhibitors on OFX sensitivity (Fig. 1). However, isolates with MICs of 16 mg/L or greater were more sensitive than those with MICs of 4 or 8 mg/L and those with MICs of 2 mg/L or less (Fig. 2), indicating that the inhibitory effects by the efflux pump inhibitors were related to the level of drug resistance. These results support the findings that efflux pump inhibitors have less of an effect in OFX-sensitive isolates of M. tuberculosis. 8
Evolution and accumulation of drug resistance drive drug sensitive individuals of M. tuberculosis to transform into MDR-TB and XDR-TB.18,24 Our previous data showed that the spectrum and level of drug resistance increased concomitantly in M. tuberculosis isolates. 30 In the present study, there were significant differences in the sensitivity of isolates with different levels of OFX resistance to efflux inhibitors, indicating that the activity of efflux pumps might be greater in clinical isolates with greater resistance to OFX (Fig. 2). In this study, mutations in gyrA were present in 89.2% of the tested OFX-resistant M. tuberculosis clinical isolates. But the percentages of mutations in gyrA were different in isolates with different levels of OFX resistance; the highest percentage was in isolates with MICs greater than 16 mg/L, and the lowest percentage was in isolates with MICs no more than 2 mg/L. In other words, M. tuberculosis clinical isolates with high levels of OFX resistance had mutations in gyrA and efflux pumps with high activity. One needs to be aware that those isolates without mutations in QRDR of gyrA do not mean that there is no mutation in other FQ-resistant determining genes, such as gyrB19,22 and fluoroquinolone resistance protein A (MfpA) and MfpB, 32 etc., which we did not tested. Singh et al. reported that there was a temporal interplay between efflux pumps and target mutations in development of antibiotic resistance in E. coli, and that early low-level resistance conferred by efflux pumps preceded and facilitated development of high-level resistance mediated by target site mutation(s). 28 Studies on the global transcriptional profiling of longitudinal clinical isolates of M. tuberculosis also showed that without specific mutations in drug target genes, activated drug efflux pumps (such as drrA) may promote the survival of M. tuberculosis prior to the acquisition of drug resistant mutations, which was thought to have the least fitness cost. 3 Schmalstieg et al. also reported that the first step in a general pathway to drug resistance is the introduction of efflux pumps, which leads to high-level chromosomal-mutation-related resistance in mycobacteria. 26
The effect of efflux pump inhibitors on different mycobacterial species, especially non-tuberculous mycobacteria (NTM), has also been tested. 10 Gupta et al. reported that addition of efflux pump inhibitors (CCCP and verapamil) to different mycobacterial species and isolates could reverse (to various degrees) the resistance to all major anti-tuberculous drugs (rifampicin, isoniazid, streptomycin, OFX).1,4,10,11 Our analysis on the inhibitory effects of three efflux inhibitors on isolates with the Beijing and non-Beijing genotypes indicated no significant difference, although only 13 isolates non-Beijing isolates were examined.
In conclusion, the results presented here indicate that OFX-resistance is due to missense mutations in the QRDR of gyrA and to changes in drug efflux pump activity. Our data confirmed that the inhibition to efflux pump activity by inhibitors was much greater for those OFX-resistant isolates with higher MICs. Although the 102 isolates had different susceptibility to OFX in the presence of efflux pump inhibitors, there was no significant difference in the effect of these inhibitors on Beijing and non-Beijing genotypes.
Footnotes
Acknowledgments
This work was supported by the National Special Key Project of China on Major Infectious Disease (2013ZX10003009-001), National Natural Science Foundation of China (30901283) and High-Level Technical Personnel Training Project in Health of Beijing (2011-3-069).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.