
Abstract
To study the characteristics of plasmids harboring oqxAB among blaCTX-M-negative Escherichia coli isolates and search for oqxAB-harboring plasmids similar to plasmids carrying oqxAB-blaCTX-M reported previously, conjugation experiment was performed for 115 randomly selected oqxAB-positive but blaCTX-M-negative E. coli isolates from diseased animals in Guangdong, China. S1 nuclease pulsed-field gel electrophoresis (PFGE) and southern blotting experiments were performed to investigate the location of oqxAB and other resistance genes. The EcoRI digestion profiles of the plasmids with oqxAB were also analyzed. The clonal relatedness of donor isolates was investigated by PFGE. In this study, 32 oqxAB transconjugants were successfully obtained and most transconjugants showed multidrug resistances. Eleven replicon combination types were found in these transconjugants. floR and oqxAB were found on the same plasmids in all nine transconjugants resistant to florfenicol. The sequences between floR and oqxAB were identical in most transconjugants and the two genes were both linked with tnp in insertion sequences. Nine F18:A-:B1 plasmids with only oqxAB shared identical EcoRI digestion profiles and the profiles were also identical with that of a plasmid carrying oqxAB-blaCTX-M found previously. Co-transfer of plasmids carrying oqxAB and fosA3, respectively, was also observed in one isolate. This study demonstrates the dissemination of oqxAB among blaCTX-M-negative E. coli isolates was mainly mediated by identical F18:A-:B1 plasmids. A novel arrangement of regions between floR and oqxAB might play an important role in the dissemination of floR-oqxAB. This is the first description of the genetic environment of the relationship between oqxAB and floR in E. coli.
Introduction
F
oqxAB has been found among Enterobacteriaceae from humans, animals, food, and environment,5,15,20,26 and the number of the epidemiology reports of this gene has increased recently. Most of the previous reports just focused on its prevalence until the study of the dissemination and characterization of plasmids carrying both oqxAB and blaCTX-M was reported in our previous work. 17 In that study, we found a relative low prevalence of transferable plasmids carrying both oqxAB and blaCTX-M, however, whether there are oqxAB-plasmids with the same backbone structure to the oqxAB-blaCTX-M-plasmids found previously remains unknown. With the application of antibiotics, resistance plasmid might acquire additional corresponding resistant gene to become a multidrug resistance plasmid with similar backbone structure to the original plasmid. So, the investigation of oqxAB-plasmids with the same backbone structure as oqxAB-blaCTX-M-plasmids found previously becomes very important. In China, fluoroquinolones have been widely used in veterinary medicine: ciprofloxacin has often been used for prophylaxis and the treatment of bacterial infections in pigs and poultry, while enrofloxacin has been used widely in pigs and poultry for therapeutic purposes. Olaquindox, the main substrate of OqxAB, is also commonly used as a therapeutic and preventive antibiotic in swine in China. The usage the these drugs might be one reason for the high prevalence of oqxAB in China. 5 Aside from fluoroquinolones and olaquindox, ceftiofur and florfenicol, are also used very commonly in both swine and poultry production in China. So the investigation of the characteristics of oqxAB-plasmids in blaCTX-M-negative isolates in China is very urgent. The study of the dissemination and characteristics of oqxAB-carrying plasmids among blaCTX-M-negative E. coli will provide insights into the understanding of the widespread of oqxAB and the potential of oqxAB-harboring plasmids to acquire blaCTX-M and become oqxAB-blaCTX-M plasmids as previously reported. 17
This study was conducted to characterize plasmids harboring oqxAB among blaCTX-M-negative E. coli isolates from diseased animals in Guangdong, China, and to evaluate the potential of oqxAB-harboring plasmids to acquire blaCTX-M.
Materials and Methods
Bacterial isolates
Two hundred one oqxAB-positive E. coli isolates without CTX-M were obtained according to the data published in our previous study. 17 All the E. coli isolates were recovered from diseased animals (124 from pigs and 77 from avian) in Guangdong, China, during 2002 and 2012, and one isolate per animal sampling strategy was used. The liver or heart samples selected to obtain isolates were from animals infected with E. coli, and the other samples were from animals with diarrhea. Further information about these animals, the underlying disease and possible antimicrobial pretreatment were unfortunately not available. All isolates were stored at −80°C in Luria–Bertani broth containing 30% glycerol.
Conjugation experiment
One hundred fifteen oqxAB-harboring isolates were randomly selected as donors for conjugation experiments by the broth-mating method as previously described. 4 E. coli C600, highly resistant to streptomycin, was used as the recipient in the conjugation experiments. OqxAB, conferring resistance to many drugs, just confers low-level resistance to fluoroquinolones, however, it confers high-level resistance to olaquindox and it is also the first identified genetic resistance mechanism toward olaquindox. 8 Therefore, olaquindox was used to limit the growth of the recipient in this experiment as previously described. 9 After incubation, transconjugants were selected on MacConkey agar plates containing streptomycin (1,000 mg/L) and olaquindox (32 mg/L). Transconjugants harboring oqxAB were confirmed by PCR as previously described 16 and antimicrobial susceptibility testing for the transconjugants, recipient, and donors was done according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI).
Pulsed-field gel electrophoresis analysis
The 32 oqxAB-harboring donors, from which oqxAB-positive transconjugants were obtained, were used to determine the genetic relatedness of oqxAB-positive E. coli isolates. The XbaI-digested genomic DNA of bacterial cells embedded in low-melting-point agarose were subjected to pulsed-field gel electrophoresis (PFGE) according to a protocol described previously. 7 The gels were run at 6.0 V/cm with an initial and final switch time of 0.5 and 63.8 sec, respectively, and an angle of 120° at 14°C for 22 hr. The DNA banding patterns were analyzed using BioNumerics software version 6.0 (Applied Maths, Kortrijk, Belgium), and a cutoff value of 95% of the similarity values was chosen to indicate identical PFGE types. Salmonella enterica serotype Braenderup H9812 standards served as size markers.
Plasmids analysis of transconjugants
PCR-based plasmid replicon typing was performed to characterize the conjugative plasmids carrying oqxAB. 2 Briefly, 18 primer pairs, targeting the FIA, FIB, FIC, F, HI1, HI2, I1/Ic, L/M, N, P, W, T, A/C, K, B/O, X, Y, and FIIA replicons, were used for the PCRs. Then, a replicon sequence typing scheme was used to discriminate IncF plasmid variants according to the method reported previously. 24 Alleles were assigned by submitting the amplicon sequence to the plasmid multilocus sequence typing database (www.pubmlst.org/plasmid). To investigate the number of plasmids in transconjugants and analyze the location of the oqxAB gene and other resistance genes, especially floR, in transconjugants, S1 nuclease-PFGE and southern blot analysis were performed. Briefly, whole-cell DNA of the transconjugants co-harboring oqxAB and floR genes embedded in agarose gel plugs was treated with S1 nuclease (TaKaRa Biotechnology, Dalian, China) and separated by PFGE alongside the chromosomal DNA of S. enterica serotype Braenderup H9812 digested with XbaI as marker. Subsequently, southern blot hybridization was performed with DNA probes specific for oqxB, and other resistance genes including floR and fosA3, which were nonradioactively labeled with a DIG High Prime DNA labeling and detection kit (Roche Diagnostics, Mannheim, Germany).
Plasmid DNA extraction was performed using a Qiagen plasmid midi kit (Qiagen, Hilden, Germany). To clarify whether the dissemination of a specific plasmid carrying oqxAB had occurred among the isolates, plasmids with the same replicon type were subjected to restriction fragment length polymorphism (RFLP) analysis. The RFLP analysis herein was also used to investigate whether the specific plasmid was similar to the plasmids harboring oqxAB-blaCTX-M reported previously. 17 Briefly, plasmids from these transconjugants containing a single plasmid were digested with the endonuclease EcoRI (TaKaRa Biotechnology), and then digested plasmid DNA was separated on a 0.8% agarose gel in 0.5× Tris-borate-EDTA for 3 hr at 80 V. λ DNA digested with HindIII (TaKaRa Biotechnology) was used as a size marker. Patterns differing by only a few bands (n=1–3) were assigned to the same profile.
Results
Conjugation experiment
Thirty-two transconjugants were successfully obtained from the randomly selected 115 oqxAB-carrying isolates. The minimum inhibitory concentrations (MICs) of olaquindox for all oqxAB-harboring transconjugants were similar to those for the donor isolates but were about 16- to 64-fold increases, compared with the recipient strain (Table 1). As shown in Table 1, all the donor isolates were resistant to at least one quinolone drugs, and their transconjugants showed 2- to 16-, 2- to 32-, and 2- to 32-fold increases in the MICs of nalidixic acid, enrofloxacin, and levofloxacin, respectively, when compared with the recipient strain. With regard to the MICs of ciprofloxacin, most of the transconjugants (except 18-2-T and Z7-T) were about 2- to 64-fold higher than that for the receipt. Eighteen donor isolates were high-level resistant to florfenicol (MICs ≥256 mg/L), a veterinary antibiotic commonly used in veterinary medicine and aquaculture, and almost all transconjugants (except Z7-T) showed different-level increases in the MICs of this drug. Notably, nine transconjugants also showed high-level resistance to florfenicol (MICs ≥256 mg/L). Twenty-six of the transconjugants showed additional multidrug resistance phenotypes including resistance to ampicillin, olaquindox, kanamycin, and/or tetracycline. In addition, the co-transfer of resistance to amikacin, was also observed in two transconjugants (Table 1).
A to T, city A to city T; the number 1 to 3 following the same English letter, represents farm 1 to farm 3 in the same city.
RFLP patterns differing by only a few bands (n=1–3) were assigned to the same RFLP profile.
FS3Z3CT, this strain was reported in reference by Liu et al. 17
AMI, amikacin; AMP, ampicillin; CIP, ciprofloxacin; ENR, enrofloxacin; FFL, florfenicol; KAN, Kanamycin; LEV, Levofloxacin; MIC, minimum inhibitory concentration; NAL, nalidixic acid; OLA, olaquindox; RFLP, restriction fragment length polymorphism; TET, tetracycline; UT, untypeable; ND, not determined. NA, not analyzed in these transconjugants with more than one plasmid.
PFGE analysis
Of the 32 E. coli isolates underwent PFGE analysis, PFGE patterns were successfully available for 23 isolates and no PFGE fragment pattern was obtained from the other 9 isolates. The PFGE patterns for the 23 isolates were presented in Figure 1. As shown in Figure 1, all the 23 isolates showed different XbaI-PFGE patterns.

Dendrogram of pulsed-field gel electrophoresis (PFGE) patterns of 24 donor isolates from which oqxAB-positive transconjugants were successfully obtained in this study.
Plasmids analysis of transconjugants
As listed in Table 1, the IncFII (7 different alleles) replicon types were detected in 28 transconjugants, 18 of them carrying 2 replicons (FII in combination with FIB). The most common replicon combination type in this study was F18:A-:B1. N replicon type was found in three transconjugants. In transconjugant Z31-T, three replicons were found. The floR gene, a gene that confers resistance to florfenicol, was detected in nine transconjugants with higher MIC (≥256 mg/L) to florfenicol. The donor isolates of the nine transconjugants were isolated from eight different cities (Table 1). The gene floR was also detected in 18 donor isolates highly resistant to florfenicol. To analyze whether the gene oqxAB was co-located on the same plasmid with floR, S1-PFGE and southern blot analysis were performed among the nine transconjugants. As shown in Figure 2, the floR and oqxAB genes were co-located on the same plasmid in all the nine transconjugants. The plasmids with floR and oqxAB were in the sizes varying from about 54 kb (7ZUER-T) to 170 kb (85CHANG YI-T) (Fig. 2). Three of the F46:A-:B24 plasmids (including S12-T, S48-T, and S1-T) were of the same size (about 130 kb) and their donors showed very different PFGE patterns (Figs. 1 and 2). The two F16:A-:B- plasmids in S53 and S68 with different PFGE patterns also showed similar size (about 70 kb). Interestingly, there were additional bigger bands in four transconjugants with floR in the S1-PFGE and these bands could also be hybridized with the corresponding probes (Fig. 2). To study whether a specific plasmid with the same backbone, occurred in the transconjugants with the same replicon combination type, the F18:A-:B1 plasmids of the nine transconjugants with only one band in the S1-PFGE experiment (data not shown) were successfully extracted and subjected to RFLP. As shown in Figure 3, the nine F18:A-:B1 type plasmids with only oqxAB gene shared the same EcoRI digestion profiles, however, the donors of these nine plasmids were from different farms and showed diverse PFGE patterns (Table 1 and Fig. 1). Notably, the RFLP profiles of the nine F18:A-:B1 plasmids were also identical with a F18:A-:B1 plasmid carrying oqxAB-blaCTX-M-55 from isolate FS3Z3C in our previous report 17 (Fig. 3). The four F46:A-:B24 plasmids with floR were also successfully extracted and subjected to RFLP, and three F46:A-:B24 plasmids shared the same RFLP profile. Although some of the plasmids carrying oqxAB-floR had different replicon types and RFLP profiles, we wanted to know whether the regions surrounding/between the two genes had similarity among these plasmids. We selected only one transconjugant from each group of transconjugants with the same RFLP or replicon type (including transconjugants S1-T, 101FEI-T, S53-T, S106-T, 7ZUER-T, and 85CHANGYI-T) for studying the regions between oqxAB and floR. Sequences between oqxAB and floR regions were amplified by PCRs using the nondigested plasmids as templates and then were determined by primer walking sequencing. Forward primer targeting oqxA (5′-CTTGCAC TTAGTTA AGCGCC) and reverse primer targeting floR (5′-TCACCGCCAATGTCCCGACGAT) were used to amplify the regions between oqxAB and floR. The sequences of the four PCR fragments from S1-T, 101FEI-T, S53-T, and S106-T were identical, and the sequence of a 10,054-bp segment of plasmid pS53T has been deposited in the GenBank database under the accession number KF731829. As shown in Figure 4, the sequence consisted of six different open reading frames in the following order; oqxA, oqxB, tnp in IS26, tnp in ISCR2, a hypothetical protein and floR. The initial 5,770-bp including oqxA, oqxB, and tnp in IS26, exhibited 100% nucleotide sequence identity to the plasmids pOLA52 (accession no. NC_010378), and pACN001-A (KC853434). The adjacent 4,284-bp showed 99% identity to plasmid IncA/C-LS6 from Klebsiella pneumoniae of human origin (JX442976). Notably, this region adjacent to oqxAB including the tnpA in ISCR2, a hypothetical protein and the floR resistance gene also exhibited 100% nucleotide sequence identity to the plasmid pACN001-A (KC853434), although oqxAB was 21,590-bp far away from the floR on the plasmid pACN001-A. For 7ZUER-T and 85CHANGYI-T, no PCR amplicon was obtained.
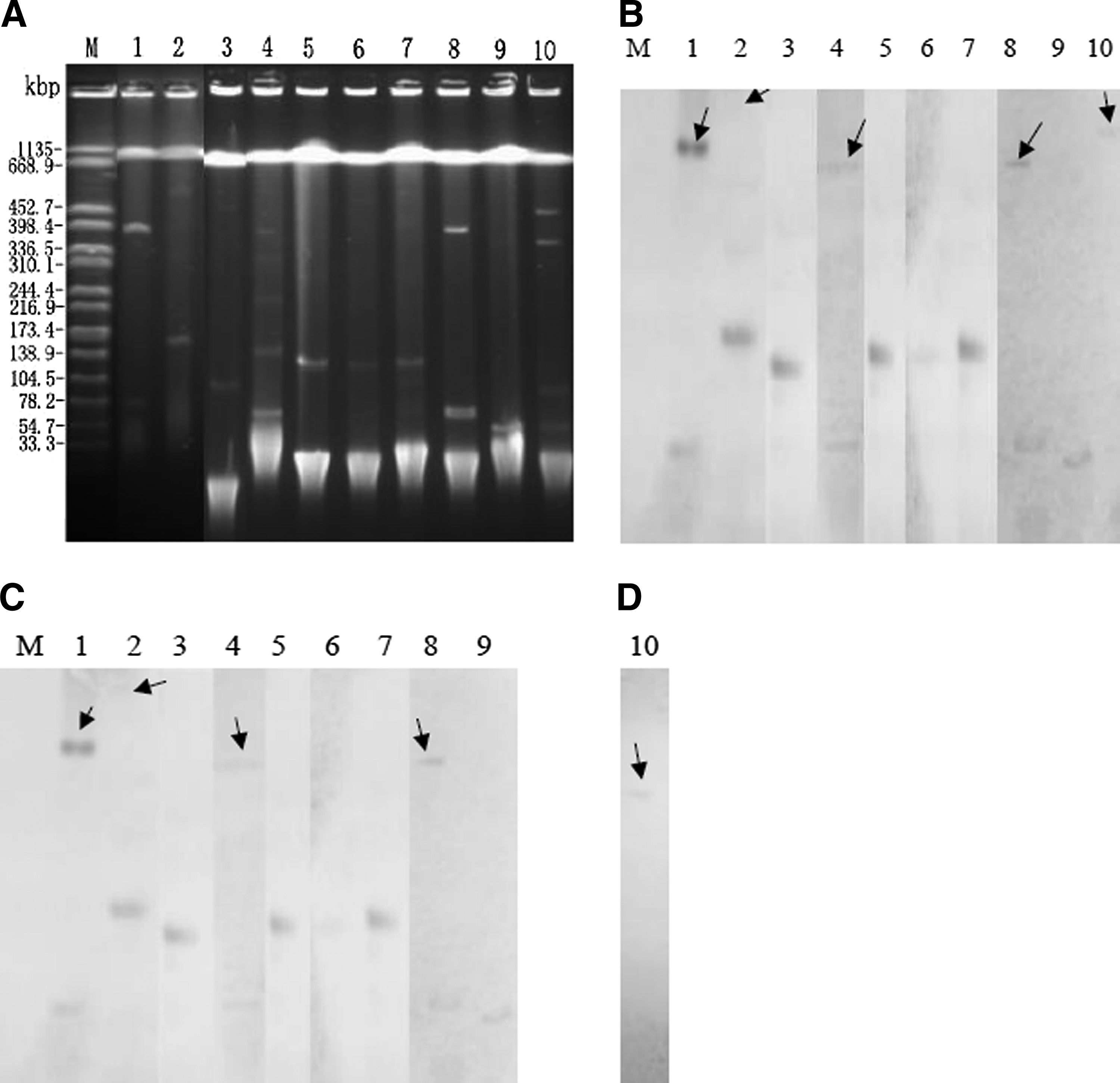
Plasmid analysis of transconjugants carrying oqxAB and floR/fosA3.

EcoRI restriction digestion profiles of F18:A-:B1 type plasmids harboring oqxAB from transconjugants containing only one plasmid. Lanes 1–10: A26-T, A58-T, Z1-T, A9-T, A44-T, 6YX4D-T, 4GAN-T, FS3Z3CT, 1GAN-T, and S45-T; Lane M: λ-HindIII marker.

Genetic environment relationship between the genes oqxAB and floR on plasmid pS53T of Escherichia coli strain S53 (accession number KF731829) and structural comparison with plasmids pOLA52 from E. coli of swine origin (NC_010378), IncA/C-LS6 from Klebsiella pneumoniae of human origin (JX442976), and pACN001-A from E. coli of avian origin (KC853434). The arrows indicate the positions and directions of open reading frames. Different genes are displayed using different shadings. Regions of >99% homology are marked by gray shading or two longer parallel lines. The arrows marked as HP and RP in the map of IncA/C-LS6 indicate reading frames for a hypothetical protein and a putative transcriptional regulator protein, respectively.
Interestingly, as shown in Table 1, co-transfer of oqxAB and fosA3, a fosfomycin resistance gene that can encode fosfomycin-modifying enzyme, was also found in one isolate recovered from the heart tissue of one duck. The S1-PFGE and southern blot analysis results indicated that the transconjugant 3YX1-T contained two plasmids and the oqxAB was located on a larger plasmid while fosA3 located on the smaller one.
Disscussion
In the present study, the dissemination and characteristics of plasmids carrying oqxAB among blaCTX-M-negative E. coli isolates from diseased animals in South China were studied, and the potential of oqxAB-harboring plasmids to aquire blaCTX-M was also evaluated. Surveys on the dissemination mechanisms of oqxAB among E. coli have been previously reported,5,28 and the characteristics of plasmids carrying oqxAB was also reported in our previous study. 17 However, the number of oqxAB transconjugants was very small in those previous studies. Furthermore, the characteristics of oqxAB plasmids among blaCTX-M-negative E. coli and whether there are oqxAB-plasmids with the same backbone structure to the oqxAB-blaCTX-M-plasmids found previously 17 were not analyzed in those previous studies. In this study, 32 oqxAB-positive transconjugants were obtained from the 115 randomly selected oqxAB-positive but blaCTX-M-negative E. coli isolates, and the conjugation rate was similar to that reported in a previous study. 28 This result indicates that oqxAB can be disseminated by horizontal transfer of plasmids, and this might be an important reason for the widespread dissemination of this gene. The diverse PFGE patterns also demonstrated that the dissemination of oqxAB was not due to the clonal dissemination of oqxAB-positive E. coli. The reason for the failure of PFGE analysis for some isolates might be the DNA degradation during electrophoresis. 14 Similar result also occurred in some Enterobacteriaceae isolates in previous reports.18,29 Quinolones MICs were increased through the acquisition of oqxAB-harboring plasmid in all the transconjugants, however, the MICs had not reached the resistant-level, suggesting the oqxAB gene contributes only to low-level decreases in susceptibility to quinolones while oqxAB can facilitate the selection of high-level resistant mutant in the future. The co-transfer of resistance to ampicillin, kanamycin, and/or tetracycline in most transconjugants carrying a single plasmid suggests that related resistance genes conferring resistances to these drugs are also located on the same plasmid with oqxAB and that the usage of these drugs will accelerate the dissemination of oqxAB.
Twenty-eight of the 32 transconjugants carried a IncF replicon type plasmid, similar to that in previous studies.17,28 These results indicate that the dissemination of oqxAB was mainly mediated by IncF-type plasmid in E. coli. In this study, floR was co-located on the same plasmid with oqxAB in all the nine transconjugants highly resistant to florfenicol, with their donor isolates from eight different cities, and this provided further proof for previous findings that floR was often found to be associated with oqxAB in blaCTX-M-positive E. coli. 17 These results also suggest that the application of florfenicol, commonly used in veterinary medicine and aquaculture, will also accelerate the dissemination of oqxAB among E. coli isolates, and then result in the increase of the prevalence of high-level fluoroquinolones resistant mutant E. coli in the future, due to the findings that increased expression of efflux pump genes (including oqxAB) is the first step in the bacterium for subsequent target mutations for resistance to higher-level fluoroquinolones.21,28 The bigger bands in the four transconjugants (Fig. 2) might be the portion of the oqxAB-carrying plasmids not exposed to S1 nuclease in the S1-PFGE experiment, according to the findings of a previous study. 1 So in this study, eight of the nine transconjugants with floR (except S106-T) carried only one plasmid. The sequences of the regions between oqxAB and floR in four tested transconjugants with different RFLP profiles or replicon types were identical, and the tnp in insertion sequences was very close to the two genes. These results indicated that oqxAB and floR could be linked and shifted together among different plasmids in E. coli strains from different isolation cities via insertion sequences. Notably, no amplicons were obtained from 7ZUER-T and 85CHANGYI-T, indicating different genetic environments between oqxAB and floR might exist. F18:A-:B1 plasmids from different animal origins in this study were found to share the same RFLP profiles, although the donors were from different farms and showed very diverse PFGE patterns. This indicated that this identical F18:A-:B1 plasmid could be horizontally transferred between isolates from different animal farms and was the main type plasmid that mediates the dissemination of oqxAB among blaCTX-M-negative E. coli isolates. Notably, the same RFLP profiles were found between the F18:A-:B1 plasmids carrying only oqxAB and a plasmid carrying both oqxAB and blaCTX-M in our previous study. This finding suggests that with the use of cephalosporins in veterinary clinics, the same F18:A-:B1 plasmid carrying only oqxAB could capture the gene blaCTX-M and the prevalence of multidrug-resistant isolates with plasmids harboring oqxAB-blaCTX-M will increase in the future. Whether it represents the trend of these plasmids in the future is unclear and further work is required to clarify this. With the increasing prevalence of isolates resistant to both cephalosporins and fluoroquinolones, there is rekindled interest in using fosfomycin for treatment of multidrug-resistant infections in human and veterinary medicine. 6 A plasmid carrying fosA3, a gene encoding fosfomycin-modifying enzyme, was found to be co-transferred with plasmid carrying oqxAB in only one isolate in this study, however, the association of fosA3 and oqxAB should be monitored in the future because fosA3 or oqxAB was often found to be co-located on the same plasmid with blaCTX-M.12,17
In conclusion, the dissemination of oqxAB among blaCTX-M-negative E. coli isolates from food-producing animals is mainly mediated by the identical F18:A-:B1 plasmids. These plasmids may acquire blaCTX-M and become oqxAB-blaCTX-M plasmids, and further study is needed to monitor and clarify this trend in the future. oqxAB and floR could be linked and shifted together among different plasmids in E. coli strains via insertion sequences. Co-transfer of two distinct plasmids carrying oqxAB and fosA3 respectively, was also found in this study. To our knowledge, this is the first description of the genetic environment of the relationship between oqxAB and floR in E. coli.
Footnotes
Acknowledgments
This work was supported by the National Science Fund for Distinguished Young Scholars (Grant No. 31125026), the Program for Changjiang Scholars and Innovative Research Team in the University of Ministry of Education of China (Grant No. IRT13063), the Natural Science Foundation of Guangdong Province (Grant No. S2012030006590), and the Science and Technology Program of Guangzhou, China (Grant No. 2011J2200054).
Disclosure Statement
The authors have no conflicts of interest to declare.