
Abstract
Diet is one of the main factors that could affect quantitatively and qualitatively the stability of the gut microbiota. Polyphenols are abundantly present in the human diet and have an antimicrobial effect inducing selective changes in the microbiota composition, with potential beneficial effects for the human health. Our aim was to determine the human gut microbiota susceptibility toward wine polyphenols. Susceptibility to two commercial wine phenolic extracts (Vitaflavan® and Provinols™) was determined in isolates from fecal samples from 36 gastrointestinal healthy volunteers. To select the polyphenol-resistant isolates, feces were seeded in plates containing 1 mg/ml of phenolic extract. The minimal inhibitory concentration to polyphenols in the collected isolates was assessed by the agar dilution method. Overall results showed that Gram-negative isolates are most tolerant to the presence of both grape seed and red wine extracts. Furthermore, we purified to homogeneity the phenolic fractions by high-performance liquid chromatography (HPLC) to determine their antimicrobial effect and their influence on bacterial growth in four selected ATCC strains using the BioScreen apparatus. Results showed that the antimicrobial activity of the wine polyphenols is the result of the interaction of both the flavan-3-ol type and the bacteria. Bacterial Intraspecies differences in the phenolic susceptibility suggest the existence of polyphenol-resistant mechanisms that are uncharacterized as yet.
Introduction
T
Diet is one of the main factors that could affect quantitatively and qualitatively the stability of the intestinal microbiota. 27 Polyphenols are phytochemicals abundantly present in the human diet contributed by fruits and vegetables and related products such as tea, coffee, wine, and chocolate. Wine and grape derivatives contain a high proportion of polyphenols, namely, flavan-3-ols in the form of monomers (i.e., catechins) and oligo-/polymers (also known as condensed tannins or proanthocyanidins) that usually escape digestion in the small intestine and reach the colon where they are metabolized by the action of gut microbiota into different metabolites that may be more biologically active than their precursor polyphenols.9,21,22
Several in vitro studies have demonstrated that phenolic compounds could selectively inhibit the growth of different bacterial species inducing changes in the microbiota composition.1,6,14,23 Administration of dealcoholized red wine polyphenols to rats increased the growth of Lactobacillus and Bifidobacterium species, 8 whereas long-term administration of condensed tannins to rats resulted in a shift in the microbial population toward Gram-negative bacteria species such as Enterobacteriaceae and Bacteroides, with reduction of the Gram-positive Clostridium leptum group. 28 This suggests that food polyphenols may act by selecting polyphenol-tolerant bacteria able to metabolize it, originating new compounds with beneficial effects in human health. The aim of the present study was to determine the human gut culturable aerobic microbiota susceptibility and/or tolerance toward wine polyphenols.
Materials and Methods
Phenolic extracts
Two commercial extracts were used in this study: (1) a grape seed extract named Vitaflavan® kindly provided by Dr. Piriou (Les Dérives Resiniques & Terpéniques, S.A.) and (2) a red wine extract named Provinols™ supplied by Safic-Alcan Especialidades, S.A.U. The phenolic composition of both extracts was previously characterized, 24 and is shown in Table 1. Gallic acid, monomeric flavan-3-ols [(+)-catechin and (−)-epicatechin], procyanidin dimers (B1, B2, B3, and B4) and trimers (C1 and T2), (−)-epicatechin-3-O-gallate and procyanidin gallates (B1-3-O-gallate, B2-3-O-gallate, and B2-3′-O-gallate) were the main compounds in Vitaflavan giving a total phenolic content of 337 mg/g. Besides the mentioned compounds, Provinols also contained anthocyanins. Both extracts were dissolved in sterile distilled water and protected against light degradation.
SD, standard deviation.
Preparation of flavan-3-ol purified fractions
Flavan-3-ol fractions were purified to homogeneity, as described previously by González-Manzano et al. 10 The compounds in the different fractions were quantified from the areas of their chromatographic peaks recorded at 280 nm by comparison with calibration curves obtained with standards of catechin, epicatechin, dimers B1, B2, B4, and B2-3′-O-gallate, and trimer C1 isolated in the laboratory.
Fecal sample processing and bacterial isolation
Fecal samples from 36 gastrointestinal healthy volunteers (one per individual) were collected and frozen at −40°C. The complete project was submitted to the local Ethics Committee, which considered that an informed consent was not necessary because the identity of the volunteers remained secret. For processing, samples were defrosted at 4°C for 24 hr, and 1 g of feces was completely homogenized with 10 ml of saline solution and centrifuged at low speed (1000 rpm) for 10 min. An aliquot of 2.7 ml of the resulting supernatant was mixed with 300 μl of (A) grape seed extract (Vitaflavan) at 10 mg/ml, (B) red wine extract (Provinols) at 10 mg/ml), or (C) sterile water (control) and incubated for 3 hr at 37°C. After that, samples were serially diluted (10−2, 10−4, and 10−6) in saline solution and plated on unspecific (Blood Agar and Tryptone Soy Agar) and selective culture media (Mac Conkey Agar, Manitol Salt Agar, M-Enterococcus Agar, and Man Rogosa Sharpe Agar). For each sample, three sets of plates were used: control, medium supplemented with 1 mg/ml of grape seed extract, and medium supplemented with 1 mg/ml of red wine extract. All culture media were purchased from Difco, and plates were aerobically incubated for 48 hr at 37°C. All phenotypic colony variants were reseeded and identified by MALDI-TOF-MS analysis (Bruker Daltonik GmbH) and/or 16S rRNA nucleotide sequencing after PCR experiments, using universal primers (16S-F 5′-AGGATTAGATACCCTGGTAGTCCA-3′ and 16S-R 5′-AGGCCC GGGAACGTATTCAC-3′) that yielded a 500 bp amplicon. All isolates were cryopreserved at −80°C in semiskimmed milk for further experiments.
Bacterial susceptibility to phenolic extracts
The ability of grape seeds and red wine phenolic extracts to inhibit the growth of selected bacteria was determined by the agar dilution method using the Steer's replicator and following the Clinical and Laboratory Standards Institute (CLSI) guidelines for antibiotics. 5 Stock solutions of phenolic extracts were dissolved in sterile water, and Tryptone Soy Agar plates were prepared at final phenolic extract concentrations ranging from 1 to 10 mg/ml. Bacterial inoculum was prepared in saline solution and suspensions were adjusted to the 0.5 McFarland standard. Plates were incubated overnight at 37°C in aerobic conditions. The minimal inhibitory concentration (MIC) value was defined as the lowest concentration of phenolic extract that inhibited visible growth.
Bioscreen experiments
The ability of growth and the fitness cost of four selected strains, Enterococcus faecalis ATCC 29212, Enterococcus faecium ATCC 19434, Staphylococcus aureus ATCC 25923, and Escherichia coli ATCC 25922, in the presence of previously purified flavan-3-ol fractions, were evaluated using the Bioscreen methodology (Labsystems). The four ATCC strains had not been previously exposed to phenolic compounds. From an overnight broth culture, serial twofold dilutions were made using fresh media to obtain inoculum of 1×105 colony-forming units/ml and transferred (350 μl) to each well of a 100-well Bioscreen plate. The five flavan-3-ol purified fractions were added at a final concentration of 1 mg/ml. The final volume of the Bioscreen well was 0.5 ml. The optical density (600 nm) was measured at 20-min intervals over a 21-hr period (37°C) with a 10-sec shaking immediately before each reading. All experiments were carried out five times and results expressed the median value obtained.
Results
Isolation of phenolic-tolerant bacteria
To select the most resistant or at least the most tolerant isolates, fecal aliquots were preincubated with both phenolic extracts and seeded in agar plates supplemented with 1 mg/ml of polyphenol. Considerable reductions of a logarithmic order of the total bacterial counts after the polyphenol exposition were observed in all samples, confirming their antimicrobial activity. This reduction was especially significant in the Manitol Salt agar plates supplemented with grape seed extract (Vitaflavan) (Fig. 1).

Mannitol Salt agar from the same fecal sample.
The overall results allowed us to recover a total of 74, 80, and 76 morphologically different colonies from the grape seed extract, the red wine extract, and the control plates, respectively (Table 2). The polyphenolic-tolerant isolates were mostly identified as E. coli (n=42), E. faecalis (n=22), and E. faecium (n=102) (Table 2). Some species, such as Anaerococcus sp. (n=2), Klebsiella pneumoniae (n=2), and Staphylococcus equorum (n=2), were only detected in the control samples without phenol exposition (Table 2). Isolates that were not correctly identified by the MALDI-TOF MS were finally identified by 16S rRNA nucleotide sequence, detecting 15 uncultured microorganisms without any species or group assignation (Table 2).
Bacterial susceptibility to phenolic extracts
The MIC of both polyphenolic extracts was determined in the selected isolates, although to elucidate the influence of a previous incubation with phenolic extracts, we analyzed separately the isolates obtained from the control plates than those previously incubated with both phenolic extracts (Fig. 2). E. coli MIC values for both phenolic extracts were considerably higher when fecal samples had been previously incubated with subinhibitory concentrations of phenolic extracts. Differences in the MIC value distribution were also detected in all isolates that had been incubated with red wine extract.

Susceptibility distribution of the Escherichia coli (□), Enterococcus faecalis (), and Enterococcus faecium (■) isolates against the phenolic extracts. Columns showed the susceptibility to both commercial extracts, whereas the different settings of the study are represented in the rows.
Taking into account all susceptibility results, Gram-positive isolates were more susceptible to both compounds (Table 3 and Fig. 2). Considering the grape seed extract, higher differences in the MIC50/MIC90 values for E. coli, E. faecalis, and E. faecium were observed (10/10, 2/6, and 2/6 mg/ml; respectively) in comparison with the red wine extract (9/10, 4/6, and 4/5 mg/ml; respectively).
Purification of flavan-3-ol fractions
The grape seed extracts were purified to homogeneity, obtaining 5 flavan-3-ols fractions with increasing degree of polymerization. The distribution of the majority compounds present in the fractions can be summarized as follows: fraction 1 contained mostly monomers [52.4% (+)-catechin and 38.1% (−)-epicatechin] and minor amounts of procyanidin dimers; fraction 2 basically consisted of non-galloyled dimers (24.1% B1, 66.1% B2, and 6.7% B4); fraction 3 was composed of dimers (13.4% B1, 3.4% B2, and 38.7% B4), galloyled dimers (11.9% B2-3′-O-gallate), and trimers (19.9% C1); fraction 4 was rich in galloyled dimers (12.4% B2-3′-O-gallate, 10.0% B1-3-O-gallate, 8.0% B2-3-O-gallate, and 8.7% galloyled dimers) and trimers (19.9% C1 and 27.4% T2), and fraction 5 was a complex mixture of procyanidins above the trimer. In addition, the fractions also presented some minor peaks corresponding to minority or nondefined flavan-3-ols (baseline humps).
Bioscreen experiments
The influence of the five purified flavan-3-ol fractions on the growth of four representative facultative anaerobe intestinal strains was analyzed by OD600 measure of their growth rate in broth cultures (Fig. 3). All fractions showed an inhibitory effect on bacterial growth, with the greatest inhibition effect observed for the E. coli strain, which was unable to reach an OD600 of 0.4 in presence of the five flavan-3-ol purified fractions. The fractions with a greater inhibitory effect were fraction 3 (dimers, trimers, and galloyled dimers) and fraction 5 (mixture of higher procyanidin oligo/polymers). For Gram-positive strains, fraction 4 (galloyled dimers and trimers) exhibited the lowest inhibitory effect.
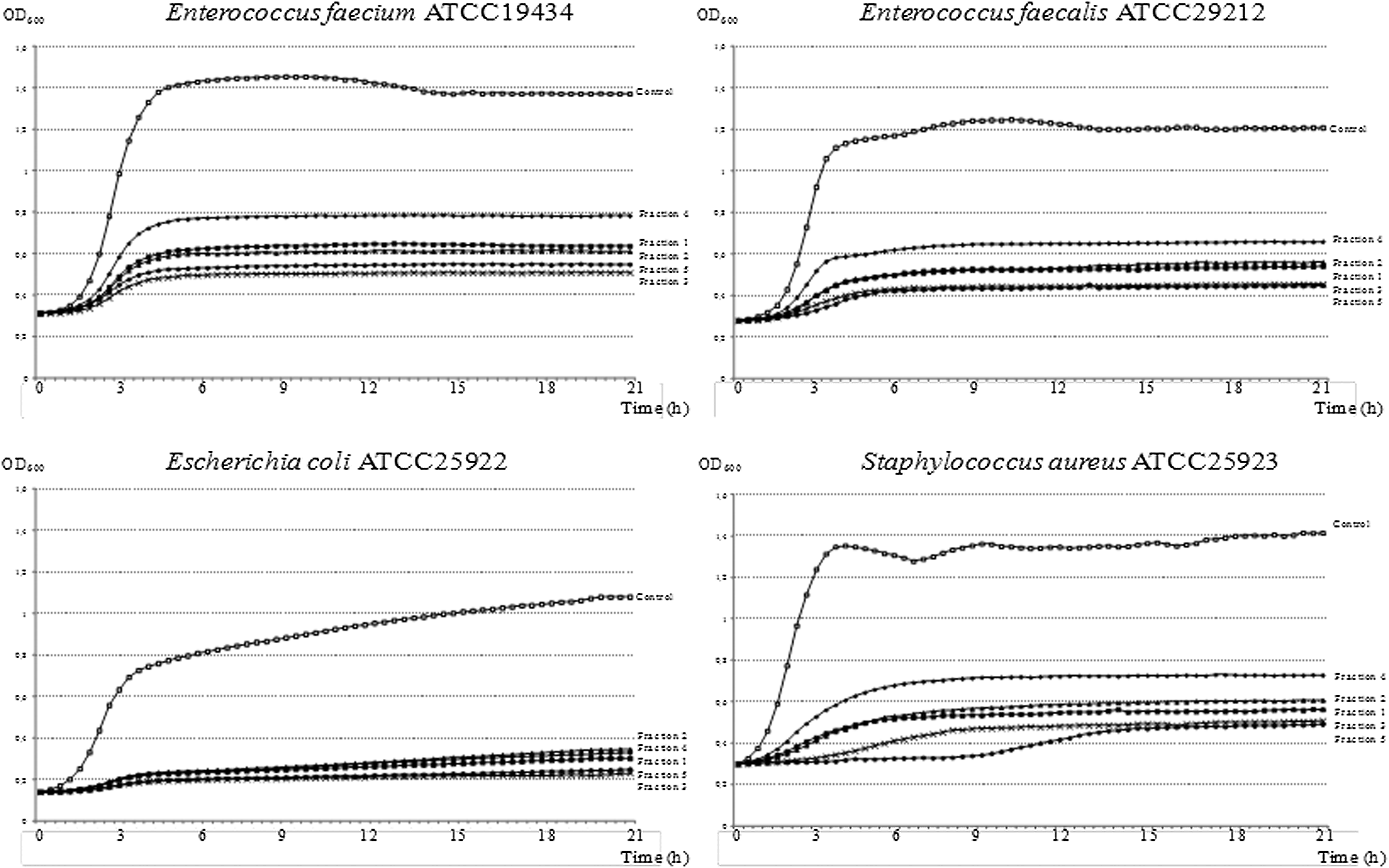
Growth curves obtained with the Bioscreen apparatus in the presence of the five purified flavans-3-ols fractions.
Discussion
In recent years, the interest to know the interactions between wine polyphenols and gut microbiota has increased due to their potential beneficial effects for human health.15,18,21,25,32,34 Several in vitro and in vivo studies with fecal microbiota have reported that polyphenols lead to changes in the human gut microbiota to their antimicrobial activity.31,33 However, knowledge concerning the interactions and the subsequent metabolism between bacteria and polyphenols is still limited.
In the first part of the present work, the in vitro tolerance of human gut microbiota to phenolic extracts was evaluated. As occurs with antibiotics, bacteria could present inducible mechanisms able to degrade phenols that must be activated by the polyphenol presence. For this purpose, fecal samples were incubated in the presence of subinhibitory concentrations of both extracts (1 mg/ml) to isolate phenolic-tolerant bacteria. Results indicated that the grape seed extract, Vitaflavan, exhibited a higher quantitative inhibitory effect in the human gut microbiota, especially Gram-positive bacteria such as the Staphylococcus genus. A combined effect of the phenol causing membrane destruction and the salt present in the Manitol Salt agar medium provoking osmotic stress might be the real cause of the bacterial count decrease, as shown in Fig. 1.
Overall results showed that tolerance to both phenolic extracts was higher in Gram-negative than Gram-positive bacteria (Table 3 and Fig. 2). All Gram-negative bacteria showed higher MIC values for the grape seed extract (≥10 mg/ml) than for the red wine extract (≥8 mg/ml), with the exception of the Klebsiella oxytoca isolates. Variations in antimicrobial activities might be attributable to differences in the composition of the extracts (Table 1) and also in cell surface structures between Gram-negative and Gram-positive species.6,30 It has been reported that the high procyanidin content in grape seed extract caused an inhibitory effect on the growth of S. aureus, Streptococcus pyogenes, Staphylococcus epidermidis, and Haemophilus influenzae. 16 A previous study also showed that the same grape seed extract used in this study was able to inhibit Lactobacillus spp. and Streptococcus salivarius strains. 30 In addition to procyanidins, the grape seed extract is rich in monomeric flavan-3-ols (catechin, epicatechin, and epicatechin-3-O-gallate) and gallic acid, for which antimicrobial activity has been also reported,6,12,14,20 and therefore, it should not be ruled out as a synergistic effect.
Most of the bacteria isolated after the fecal incubations with the grape seed extract belonged to the Enterobacteriaceae family. It has been reported that condensed tannins (proanthocyanidins) allowed the growth of Enterobacteriaceae and Bacteroides species. 28 In contrast, Yamakoshi et al., 36 observed that proanthocyanidin-rich extracts from grape seeds led to the inhibition of Enterobacteriaceae species.
Resistance to polyphenols in E. coli has been related to the expression of a complex set of transcriptional responses for maintaining the integrity of the cell membrane. 29 Different mechanisms have also been proposed, such as modification and degradation of polyphenols, interaction of polyphenols with membranes, cell walls and/or extracellular proteins, and metal ion chelation, which may overcome the inhibitory effects of dietary polyphenols. 29 In addition, it has been reported that the increasing oxidative stress response under aerobic conditions contributes to select proanthocyanidin-resistant E. coli strains. 28 Surprisingly, most of the assayed Gram-negative bacteria, including E. coli strains, were slightly more susceptible to the red wine extract in spite of its total phenolic content being lower compared with the grape seed extract. A possible explanation might be that the relatively low procyanidin content of the red wine extract was insufficient to induce an oxidative stress response.
On the contrary, E. faecalis and E. faecium showed a higher susceptibility to the grape seed extract, which could be related to their absence of protective outer membrane.12,26 In addition, it was observed that red wine-selected E. faecalis isolates showed more tolerance to both compounds than the isolates obtained after grape seed extract incubation. This result suggested that those anthocyanins and other phenolic compounds, which are only present in the red wine extract, might increase the E. faecalis tolerance.
To investigate the phenolic compound influence, five high-performance liquid chromatography-purified flavan-3-ols fractions were tested against four reference strains without previous contact with phenolic compounds. These strains were selected in representation of the anaerobe facultative intestinal microbiota. The growth of the four strains was affected by the flavan-3-ols fractions, although total inhibition was not observed, probably due to optimization of metabolic pathways involved in the degradation of the fractions. Growth curve analysis curiously demonstrated that E. coli ATCC 25922 was the most susceptible strain. This result contrasted with the high tolerance to phenolic extracts observed for clinical isolates of E. coli and highlights the importance of the diet in the development of phenolic-tolerant bacteria. 19 In this sense, some authors have proposed that a common exposure to dietary phenolics may be responsible for the selection of polyphenol-tolerant strains of intestinal microbiota,2,14 although a recent study concluded that a long-term exposure to polyphenols might be needed to induce polyphenol resistance and bioconversion genes on the microbiota. 13
On the other hand, the growth of Gram-positive bacteria was particularly affected by fraction 3 (dimers, trimers, and galloyled dimers) and 5 (procyanidins mixture), whereas the fraction 4 exhibited the lowest inhibitory effect. The highest content in galloyled compounds in fraction 4 relative to fractions 2 and 3 seems to suggest that galloylation reduces the inhibitory activity of procyanidin oligomers. Therefore, our results indicated that the inhibitory activity in the bacterial growth increased with flavan-3-ol polymerization and decreased with galloylation. 17
The experiments of this work have been performed in aerobic conditions and due to that, one of the main limitations is the absence of data about the anaerobic microbiota, which is the main group of the total gut bacteria. The overall susceptibility of these anaerobic bacteria might be also explored to better understand the global reaction to polyphenols in the gut. Another point that needs attention is the importance of phenolic structure (composition and the degree of polymerization); flavan-3-ol monomers are mainly absorbed in the small intestine, whereas oligomers and polymers reach the colon where they can be extensively metabolized by the microbiota to produce phenolic metabolites, including simple phenols, phenolic acids, and other metabolites. In our work, we tried to cover this aspect as it includes a detailed study using phenolic fractions of different compositions and degree of polymerization and using physiological concentrations that could be reached in the gut.
We hypothesized that similar to antibiotics might exist different molecular and phenotypic mechanisms that allow the bacteria to survive in the presence of phenols and potentially to metabolize it. In fact, two different points might be explored: the induction of tolerant bacteria after continuous polyphenol exposition by nonacquired mechanisms, and the existence of molecular mechanisms conferring phenolic resistance that could be transferred to other bacteria as occurs with antibiotic-resistant determinants. In conclusion, results of our study highlight the variability in the tolerance to grape and wine phenolic compounds among cultivable facultative anaerobe and aerobe bacteria from human gut microbiota, the Gram-negative isolates being the most tolerant to both grape seed and red wine extracts. Further assays were performed with fractions containing purified flavan-3-ols, the majority phenolic compounds in grape and wine extracts lead us to understand that the antimicrobial activity was mainly conditioned by both, flavan-3-ol-type and bacterial species. Nevertheless, further investigations are necessary to elucidate the mechanism by which some intestinal bacteria are able to resist the inhibitory effects of polyphenols.
Footnotes
Acknowledgments
Work in the laboratory of the authors was funded by the Spanish Ministry for Science and Innovation (AGL2009-13361-C02-00, AGL2010-17499, and CSD2007-00063 Consolider Ingenio 2010 FUN-C-FOOD Projects) and the Comunidad de Madrid (S-0505/AGR/0153 Project). C.C. is the recipient of fellowships from the FPI-MEC program.
Disclosure Statement
No competing financial interests exist.