
Abstract
Carbapenems are last-resort antibiotics to handle serious infections caused by multiresistant bacteria. The incidence of resistance to these antibiotics has been increasing and new resistance mechanisms have emerged. The dissemination of carbapenem resistance in the environment has been overlooked. The main goal of this research was to assess the prevalence and diversity of carbapenem-resistant bacteria in riverine ecosystems. The presence of frequently reported carbapenemase-encoding genes was inspected. The proportion of imipenem-resistant bacteria was on average 2.24 CFU/ml. Imipenem-resistant strains (n=110) were identified as Pseudomonas spp., Stenotrophomonas maltophilia, Aeromonas spp., Chromobacterium haemolyticum, Shewanella xiamenensis, and members of Enterobacteriaceae. Carbapenem-resistant bacteria were highly resistant to other beta-lactams such as quinolones, aminoglycosides, chloramphenicol, tetracyclines, and sulfamethoxazole/trimethoprim. Carbapenem resistance was mostly associated with intrinsically resistant bacteria. As intrinsic resistance mechanisms, we have identified the blaCphA gene in 77.3% of Aeromonas spp., blaL1 in all S. maltophilia, and blaOXA-48-like in all S. xiamenensis. As acquired resistance mechanisms, we have detected the blaVIM-2 gene in six Pseudomonas spp. (5.45%). Integrons with gene cassettes encoding resistance to aminoglycosides (aacA and aacC genes), trimethoprim (dfrB1b), and carbapenems (blaVIM-2) were found in Pseudomonas spp. Results suggest that carbapenem resistance dissemination in riverine ecosystems is still at an early stage. Nevertheless, monitoring these aquatic compartments for the presence of resistance genes and its host organisms is essential to outline strategies to minimize resistance dissemination.
Introduction
T
Carbapenems, such as imipenem, meropenem, and ertapenem, are considered last-resort antibiotics commonly applied to treat severe infections when all other therapeutic options fail. 8 In some countries, including Portugal, the use of carbapenems is still limited to hospital settings. 18 However, there has been a continuous increase in the prevalence of bacterial resistance to carbapenems in clinical settings.33,35 Some resistance mechanisms are still geographically constrained, but others have spread worldwide. 35 The most common mechanism of resistance to carbapenems in gram-negative bacteria is the production of carbapenemases. Carbapenem-hydrolyzing beta-lactamases may belong to one of the following Ambler classes: class A (e.g., KPC), class B (e.g., VIM, NDM), and class D (e.g., OXA-48). 8 Clinical isolates producing carbapenemases are mostly members of Enterobacteriaceae and nonfermenters such as Pseudomonas spp. and Acinetobacter spp.8,33,35,42
Antibiotic resistance dissemination in natural environments is nowadays recognized as a major public health problem. Despite that, the prevalence and diversity of carbapenem-resistant strains and carbapenemases-encoding genes in bacterial isolates of environmental origin are still overlooked. Even so, novel carbapenemases have been sporadically described in some environmental strains. That is the case of BIC-1 in Pseudomonas fluorescens 17 and Sfh-I and SFC-1 in Serratia fonticola.20,45 Carbapenemases that are largely disseminated in clinical settings have also been detected in strains isolated from different environmental sources. Examples of clinically relevant carbapenemases present in strains isolated from rivers or wastewater include KPC,11,37,38 VIM,12,43,44 IMP, 12 OXA-48,15,41 and NDM.22,55,58 Moreover, the genes encoding several carbapenem-hydrolyzing enzymes have their putative origin in the genomes of species commonly found in natural settings. Two examples are the class D carbapenemases OXA-23 in Acinetobacter radioresistens 39 and OXA-48 in Shewanella spp.40,50
Most information reported above occurred as sporadic observations. Few studies have addressed specifically bacterial resistance to carbapenems in natural environments. Data on the diversity of carbapenem-resistant bacteria and the resistance mechanisms they express in nature are still scarce.
The main goal of this research was to assess the prevalence and diversity of carbapenem-resistant bacteria in rivers. Carbapenem-resistant isolates were inspected for the presence of frequently reported carbapenemase-encoding genes.
Materials and Methods
Sample collection and river water quality status
Twelve water samples were collected in 12 sampling sites located in 11 rivers from the Vouga River basin, central Portugal (Table 1). Most sampling sites are impacted by different pollution sources of agricultural, industrial, and domestic origins. Previously analyzed physical, chemical, and microbiological parameters showed that these rivers displayed different levels of superficial water quality, from unpolluted to polluted characteristics, according to the national legislation for water quality categorization (for details, see Tacão et al. 49 ). Water was collected in sterile bottles (7 L) from 50 cm below the water surface and kept on ice for transportation.
IMIR, imipenem resistant.
Enumeration and selection of imipenem-resistant bacteria
River water was filtered in sterile 0.45-μm-pore-size cellulose ester filters, and the membranes placed on MacConkey agar plates (Merck) supplemented with 8 μg/ml of imipenem, the first carbapenem available for treating infectious diseases. 34 The MacConkey medium was used to select for gram-negative phylogenetic groups that are currently the greatest threats in terms of carbapenem resistance (e.g., Enterobacteriaceae, Aeromonas, Pseudomonas).8,33 The total filtered volumes varied from 1 to 500 ml, according to preliminary studies conducted in each sampling site. MacConkey agar plates without an antibiotic supplement were used to determine the proportion of imipenem-resistant bacteria. Plates were incubated at 37°C for 16 hr. Colony counting was done in triplicate. Mean bacterial counts were calculated for each site. Individual imipenem-resistant colonies were purified and stored in 20% glycerol at −80°C.
Molecular typing and identification of imipenem-resistant isolates
BOX element polymerase chain reaction (BOX-PCR) was used to type all isolates as previously described. 49 PCR products were loaded in 1.5% agarose gels for electrophoresis and band patterns were analyzed with software GelCompar II version 6.1 (Applied Maths; available from www.applied-maths.com/). Similarity, matrices were calculated with Dice coefficient, and cluster analysis of similarity matrices was performed by the unweighted pair group method using arithmetic averages (UPGMA). 47 Isolates displaying different BOX profiles were identified by 16S rRNA gene sequencing analysis with primers and PCR conditions as previously described. 49 Aeromonas isolates were further analyzed by gyrB gene sequencing, as previously described. 57 PCR products were purified with DNA Clean & Concentrator (Zymo Research) following the manufacturer's instructions, and used as template in the sequencing reactions. Similarity searches were performed with the BLAST software 1 at the National Center for Biotechnology Information website against the GenBank database. Identification was confirmed with the EZTaxon tool available at www.ezbiocloud.net/eztaxon, 24 using on average 1,200 bp.
Antibiotic susceptibility tests
Antibiotic susceptibility patterns were determined by the agar disc diffusion method on Mueller–Hinton agar. Fourteen antibiotics from six classes were tested: beta-lactams, quinolones, aminoglycosides, phenicols, tetracyclines, and the combination sulfamethoxazole/trimethoprim. Discs containing the following antibacterial agents were used according to the European Committee on Antimicrobial Susceptibility Testing (EUCAST) guidelines 14 : amoxicillin (10 μg), amoxicillin/clavulanic acid (20 μg/10 μg), cefotaxime (5 μg), cefepime (30 μg), imipenem (10 μg), ertapenem (10 μg), aztreonam (30 μg), ciprofloxacin (5 μg), gentamicin (10 μg), chloramphenicol (30 μg), and sulfamethoxazole/trimethoprim (25 μg) (Oxoid). Due to lack of EUCAST information for kanamycin (30 μg), nalidixic acid, (30 μg) and tetracycline (30 μg), the Clinical Laboratory Standards Institute criteria (CLSI) were used. 13 Also, CLSI guidelines were used for Pseudomonas spp. regarding chloramphenicol and sulfamethoxazole/trimethoprim. For Aeromonas spp. and Chromobacterium haemolyticum, we followed breakpoint tables for Enterobacteriaceae. After 24 hr of incubation at 37°C, results were analyzed following the corresponding guidelines.13,14 The Escherichia coli ATCC 25922 was used as quality control. Detection of extended-spectrum beta-lactamase (ESBL) production was carried out by a clavulanic acid combination disc method based on comparing the inhibition zones of cefpodoxime (10 μg) and cefpodoxime-plus-clavulanate (10/1 μg) discs (Oxoid).13,49
PCR amplification of antibiotic resistance determinants
Genes conferring resistance to beta-lactams (blaSHV, blaTEM, blaSPM, blaAIM, blaGIM, blaDIM, blaIMP, blaVIM, blaKPC, blaGES, blaNDM, blaCphA-like, blaL1, blaCTX-M, blaPER, blaVEB, and blaOXA-48), to tetracycline [tet(A), tet(B), tet(C), tet(D), tet(E), tet(G), tet(O), and tet(M)], and to fluoroquinolones (qnrA, qnrB, qnrVC, and aacA4-cr) were inspected by PCR using previously described primers and conditions (Supplementary Table S1; Supplementary materials are available online at http://www.liebertpub.com/mdr). Genomic DNA of positive control strains was used.2,19,49–51 Each experiment included also a PCR mixture containing water instead of DNA as a negative control. Results were confirmed by sequencing.
Integron screening and characterization
Integrase screening was performed for intI1 and intI2 genes. 21 The variable regions of integrase-positive isolates were further amplified by PCR as described before 51 and sequenced.
Nucleotide sequence accession numbers
All the nucleotide sequences stated in this work have been deposited in the GenBank database under the accession numbers KJ396795–KJ396890 (16S rRNA gene sequences), KR232493–KR232507 (gyrB genes from bacterial isolates), KJ620481–KJ620486 (gene cassette arrays), KM495226–KM495239 (blacphA genes from bacterial isolates), and KM495240–KM495266 (blaL1 genes from bacterial isolates).
Results
Prevalence and phylogenetic diversity of imipenem-resistant bacteria
Colony counts on MacConkey agar were on average 9.8×102 CFU/ml of riverine water. In the same media supplemented with imipenem, colony counts of 0.19×102 CFU/ml were obtained. This corresponds to 0.16% of the bacteria able to grow in the presence of imipenem. Results are shown in Table 1.
Clonal relationships between 184 imipenem-resistant isolates were evaluated by BOX-PCR. A total of 110 isolates displayed unique BOX profiles and were selected for further analysis (Table 2). Phylogenetic affiliation resulting from the analysis of 16S rRNA gene sequences is shown in Table 2. Detailed information is available in Supplementary Table S2.
Overall, the most frequent genus was Pseudomonas, representing 41.8% of the total number of strains, including several species Pseudomonas geniculata, Pseudomonas beteli, Pseudomonas hibiscicola, Pseudomonas aeruginosa, Pseudomonas protegens, Pseudomonas otitidis, Pseudomonas putida, Pseudomonas taiwanensis, and Pseudomonas sp. Strains identified as Stenotrophomonas maltophilia represented 24.5% of the total. Bacteria belonging to genus Aeromonas were 20.0% and included the following three species: Aeromonas veronii, Aeromonas hydrophila, and Aeromonas jandaei. The species C. haemolyticum (8.2%) and Shewanella xiamenensis (2.7%) were also found. Members of the Enterobacteriaceae (2.7%) included the species Enterobacter ludwigii, Enterobacter asburiae, and Providencia alcaligenes.
Antimicrobial susceptibility testing
Figure 1 presents the prevalence of resistance to each antibiotic tested. As expected, imipenem-resistant isolates showed resistance to both carbapenems tested (imipenem and ertapenem) and to ampicillin. Resistance to the third-generation cephalosporin cefotaxime was found on 88.2% of the isolates, and 62.7% were resistant to the fourth-generation cephalosporin cefepime. In what concerns non-beta-lactam antibiotics, the higher percentages of resistance were observed against aminoglycosides (particularly against kanamycin with 68.2% resistant isolates), tetracycline (50.9%), nalidixic acid (43.6%), chloramphenicol (43.6%), and sulfamethoxazole/trimethoprim (33.6%). The presence of ESBLs was evaluated using the double disc diffusion test. The results were negative for all isolates. All S. maltophilia and P. aeruginosa isolates presented resistance profiles in agreement to what has been already described as intrinsically resistance mechanisms. 25 Also, aeromonas presented susceptibility profiles in agreement to what has been described for the majority of Aeromonas spp., particularly in what concerns resistance to non-beta-lactams. 23 Besides intrinsic resistance, 44.4% of S. maltophilia isolates were multiresistant, that is, resistant to three or more classes of antibiotics, including tetracyclines, quinolones, chloramphenicol, and sulfamethoxazole/trimethoprim.
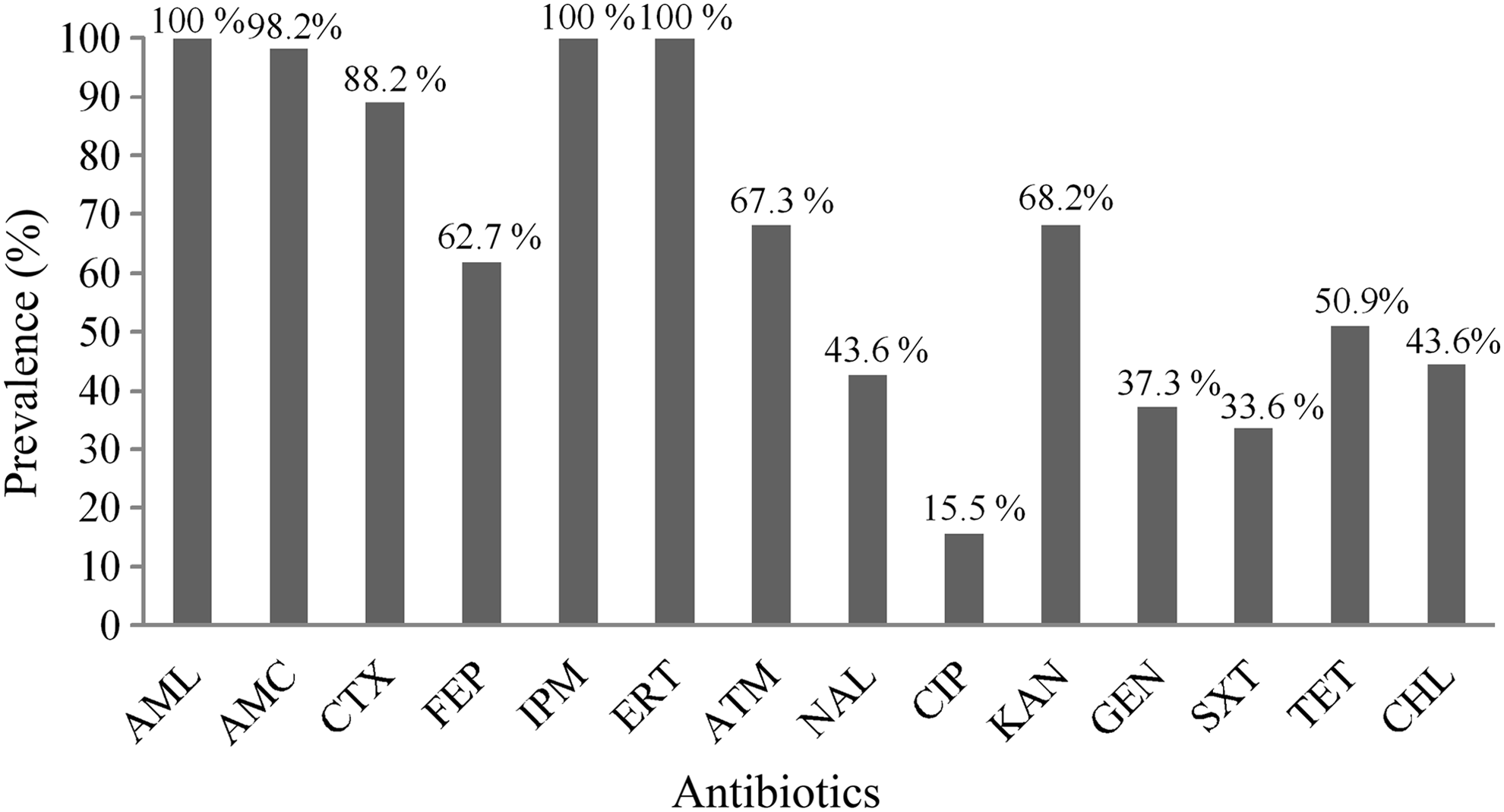
Prevalence of isolates (%) resistant to amoxicillin (AML), amoxicillin/clavulanic acid (AMC), cefotaxime (CTX), cefepime (FEP), imipenem (IPM), ertapenem (ERT), aztreonam (ATM), nalidixic acid (NAL), ciprofloxacin (CIP), kanamycin (KAN), gentamicin (GEN), combination trimethoprim/sulfamethoxazole (SXT), tetracycline (TET), and chloramphenicol (CHL).
Cluster analysis of the antibiotic susceptibility profiles of all pseudomonads (Fig. 2) showed that the isolates group preferentially according to their phylogenetic affiliation. There is a clear separation in two clusters: one that includes members of genus Pseudomonas sensu stricto (mostly P. aeruginosa, P. protegens, and P. otitidis)31,36 and the other composed of members of the genus Pseudomonas more closely related to the genus Stenotrophomonas but still listed as pseudomonads (P. geniculata, P. beteli, and P. hibiscicola).31,36 Whereas members of this second cluster showed resistance to all beta-lactams tested, most isolates in the Pseudomonas sensu stricto cluster showed resistance to penicillins and third-generation cephalosporins, but susceptibility to fourth-generation cephalosporins and monobactams. Analyzing resistance toward non-beta-lactams, differences are also clear between these groups; while the majority of Pseudomonas sensu stricto isolates are resistant to nalidixic acid and the combination sulfamethoxazole/trimethoprim, members of the second cluster are mostly resistant to kanamycin and tetracycline.
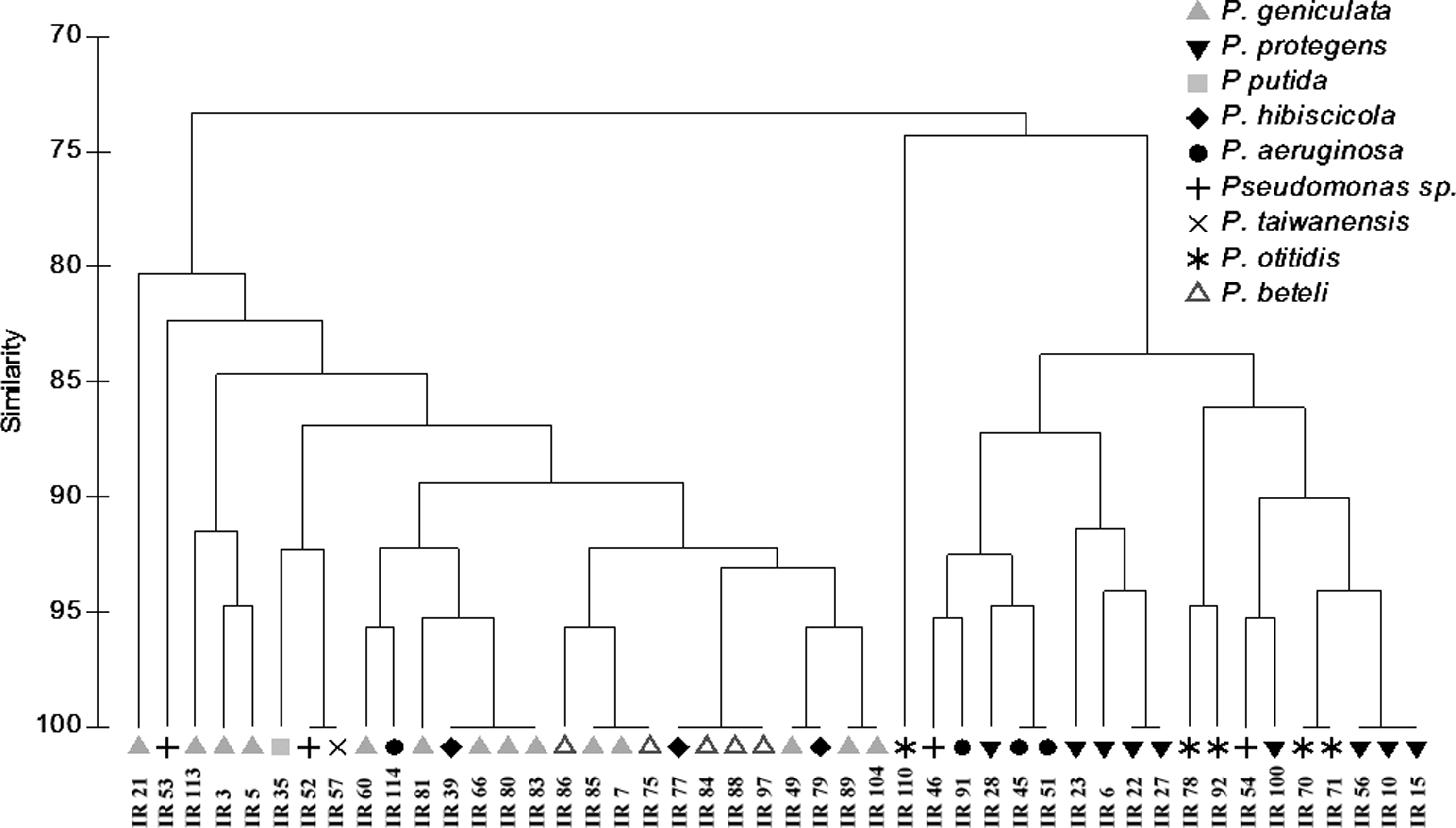
Cluster analysis of the antibiotic susceptibility profiles of Pseudomonas isolates from river water, using Bray–Curtis similarity coefficient and unweighted pair group method using arithmetic averages cluster methods.
Occurrence and diversity of antibiotic resistance genes
The imipenem-resistant isolates were further analyzed by PCR with the primer sets specific to several antibiotic resistance genes, as detailed in the Materials and Methods section. The carbapenemase genes blaCphA-like were detected in 77.3% of Aeromonas spp. (17 of 22) (Table 3). Sequencing results of the blaCphA amplicons displayed six genotypes and the deduced amino acid sequences resulted in six CphA variants. CphA-like sequences varied from 95.2% to 99.5% similarity between them and from 99% (one amino acid difference) to 100% identities with those previously described. The blaL1 gene was detected in all S. maltophilia (n=27) (Table 3), showing seven different blaL1 genotypes. The deduced amino acid sequences comprised four L1 variants, with 89.3–99.4% similarity between them and 99% (two different amino acids) to 100% identities with previously described L1 carbapenemases. blaVIM was detected in 13% of Pseudomonas isolates (6 of 46) (Table 3). Sequencing results showed that the six blaVIM-positive isolates (two Pseudomonas sp. and one representative of each of the species P. putida, Pseudomonas monteilii, and P. geniculata) carried a blaVIM-2 gene. Also, blaOXA-48-like genes were detected in all S. xiamenensis isolates, as previously described. 50
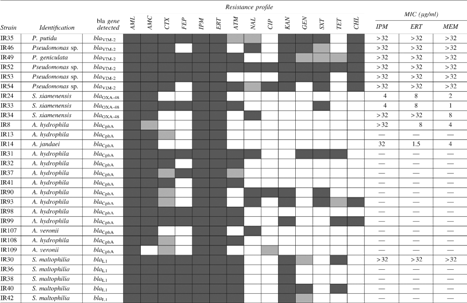

Identification, isolation site, bla genes detected, antibiotic susceptibility profiles (dark gray, resistant; light gray, intermediate; white, susceptible) and MIC determined for IPM, ERT, and MEM.
—, not determined; AMC, amoxicillin/clavulanic acid; AML, amoxicillin; ATM, aztreonam; CHL, chloramphenicol; CIP, ciprofloxacin; CTX, cefotaxime; ERT, ertapenem; FEP, cefepime; GEN, gentamicin; IPM, imipenem; KAN, kanamycin; MEM, meropenem; MIC, minimal inhibitory concentrations; NAL, nalidixic acid; SXT, combination trimethoprim/sulfamethoxazole; TET, tetracycline.
Genes encoding resistance to tetracyclines, aminoglycosides, or quinolones were not detected with the primers used in this study.
Integron characterization
The gene intI1 was detected in two Pseudomonas isolates (IR35 and IR49), both carrying the carbapenemase gene blaVIM-2. The integron variable regions were analyzed. The gene cassette arrays identified conferred resistance to aminoglycosides (aacA and aacC type genes), trimethoprim (dfrB1b), and beta-lactams (blaVIM-2). In P. putida IR35, the gene cassette array was aacA7-blaVIM-2-aacC1-aacA4 and in P. geniculata IR49 was dfrB1b-aacA4-blaVIM-2. The gene cassette array blaVIM-2–aacA4 was identified in four Pseudomonas spp. isolates (IR46, IR52, IR53, and IR54) using primers targeting both genes, but no integrase gene was detected.
Discussion
The increasing prevalence of antibiotic resistance in bacterial strains is problematic for public health. Particularly worrying is the growing numbers of clinical isolates resistant to last-resort antibiotics such as carbapenems.
In this study, we reported the prevalence and diversity of carbapenem-resistant bacteria in river water in Portugal. Isolates growing in the presence of imipenem were investigated with respect to specific resistance genes and genetic elements responsible for resistance dissemination. The use of imipenem for the selective cultivation may have underestimated the diversity of carbapenem-resistant bacteria and resistance mechanisms. For example, Pseudomonas strains susceptible to imipenem and resistant to other carbapenems have been frequently reported. 10 We have chosen imipenem as the selective agent since it was the first carbapenem available for the treatment of serious infections 34 and is still widely prescribed. In Portugal, its use is restricted to hospital settings to prevent resistance dissemination. 18
When compared with the proportion of cefotaxime-resistant bacteria (4.64%) found in the same rivers and sampling period, 41 the percentage of imipenem-resistant isolates (0.19%) was clearly lower. A low prevalence of imipenem-resistant bacteria was found in a previous study on bacteria from untreated drinking water in Portugal. 18 These numbers might be related to the restrictive rules of administration of carbapenems in Portuguese clinics. 18 The lack of similar studies in geographic regions with different carbapenems prescription policies prevents a comparison that would be of major interest.
Noteworthy, in this study, the highest prevalence of imipenem-resistant bacteria was detected in the river presenting the poorest water quality status (R2 in Table 2). This river water showed high concentrations of phosphates and ammonia (2.7 and 5.5 mg/L, respectively), as well as high loads of fecal streptococci, fecal coliforms, and total coliforms (all above 500 CFUs/100 ml). 49
This indicates that, as observed previously for cefotaxime resistance, 49 human activities may influence the prevalence of carbapenem resistance in aquatic systems. The river water here studied is impacted by pollution from different sources, which include domestic, agricultural, and industrial effluents. 49 The high levels of imipenem resistance detected in this polluted river might be related to coselection events driven by the presence of other contaminants rather than antibiotics. These types of effects were reported in other studies.5,46 Another explanation may be the presence of carbapenem-resistant fecal-derived bacteria, selected by antibiotic therapy in humans or other animals. However, the most common groups of fecal bacteria were not represented in our culture collection. Nevertheless, it is possible that carbapenem-resistant bacteria and the large fraction of fecal bacteria detected 49 at the most polluted site have common sources (e.g., a sewage water treatment plant).
As expected, since bacteria isolated in imipenem-containing culture media, high resistance rates were observed toward beta-lactams. The majority of the bacterial isolates selected are intrinsically resistant not only to carbapenems but also to other beta-lactams.
The isolation of carbapenem-resistant Aeromonas and S. maltophilia was of no surprise. These groups are commonly isolated from aquatic systems and intrinsically resistant to carbapenems.27,35 Resistance in S. maltophilia is determined by the expression of blaL1, encoded in a plasmid-like element considered intrinsic to this species. 4 All S. maltophilia strains isolated in this study contained the blaL1 gene. Likewise, the majority of members of the genus Aeromonas show resistance toward carbapenems due to the expression of chromosomal class B metallo-beta-lactamase genes like blaCphA. 54 This gene was also detected in the majority of aeromonas here isolated.
The genus Pseudomonas was the most represented. In general and with the exception of ertapenem, pseudomonads are sensitive to carbapenems. Nevertheless, several carbapenem resistance mechanisms have been described in pseudomonads, particularly in P. aeruginosa, by far the most studied species in this genus due to its clinical importance. Pseudomonads might carry one, or combinations of two or more, resistance mechanisms. These include high-level expression of chromosomally encoded class C beta-lactamases, reduced outer membrane permeability, and overexpression of efflux pumps with wide substrate specificity.26,30,48 These combinations act differently, depending on the antibiotic molecule. 48 Plasmid-mediated class A (e.g., BIC-1, GES-, and KPC-types) and class B (e.g., IMP- and VIM-types) carbapenemases 35 have also been detected in Pseudomonas isolates. Recently, a 3-year surveillance study performed on P. aeruginosa isolates obtained in hospitals from 14 European countries, showed an increase in the number of metallo-beta-lactamase producers (most frequently VIM-2). The same study showed that most isolates carried one or more resistance mechanisms, being the loss of OprD (implying reduced permeability) the most common cause for the high minimal inhibitory concentration (MIC) values observed. 10 Moreover, combinations of these mechanisms have been associated to resistance to unrelated classes of antibiotics, for example, resistance to all beta-lactams and also to quinolones and aminoglycosides. 48 In the present study, the majority of pseudomonads showed MIC values over 32 μg/ml for ertapenem, meropenem, and imipenem (Supplementary Table S2).
The majority of these isolates were resistant to several classes of non-beta-lactam antibiotics, mainly aminoglycosides, quinolones, and chloramphenicol. Several genes encoding resistance to these antibiotics have been described previously in Pseudomonas spp.51,53 Although we have targeted a large number of these genes, none was detected by PCR with the primers used. Most probably, other nonacquired resistance mechanisms are responsible for the phenotypes observed. In fact, there is a lack of information concerning antibiotic susceptibility profiles of pseudomonads other than P. aeruginosa. Our results indicate that different intrinsic resistance mechanisms may be present in different species of this genus. For example, all P. geniculata and P. hibiscicola isolates are resistant to all beta-lactams and P. protegens isolates are resistant to penicillins, third-generation cephalosporins, and phenicols. As this genus comprises a large number of different species, further studies are necessary to elucidate the diversity of intrinsic resistance mechanisms of pseudomonads.
We detected acquired blaVIM-2 genes 100% identical to those reported in clinical isolates. This is the most common VIM variant reported so far worldwide, 35 including in Portugal although sporadically.9,33 The blaVIM-2 was identified as a cassette included in integron arrays with other resistance genes, as frequently described. 35 Hence, this multiresistance apparatus covers several classes of antibiotics. Both gene cassette arrays were already described in clinical P. aeruginosa strains (www.integrall.bio.ua.pt). 32 These genes were detected only in isolates from low-quality river water, an alert for the fact that human-driven pressures can haste these events. Their association to mobilizable genetic platforms like integrons contributes to facilitate their dispersion.
In conclusion, the prevalence of carbapenem-resistant bacteria in aquatic environments is still low and mostly related to the presence of intrinsically resistant bacteria, at least in countries where carbapenems prescription policies are restrictive, as in Portugal. However, we have detected acquired carbapenem resistance mechanisms in river water and with poor quality according to several physicochemical and microbiological parameters. This might be related to selective pressures of human origin or with the direct discharge of antibiotic-resistant bacteria into these settings (e.g., from wastewater treatment plants). There is a need to identify the human-derived impacts that contribute to deterioration of water quality and, at the same time, promote dissemination of antibiotic resistance. The disposal of chemicals like antibiotics and other pollutants, such as metals and disinfectants, in aquatic environments needs to be strictly controlled. The same applies to materials of biological origin suspected of containing antibiotic resistance genes or antibiotic-resistant bacteria. These contaminants synergistically act to promote antibiotic resistance dissemination, with mobile genetic elements as main vehicles.5,7,16 It is important to carry out more studies focusing on these natural compartments, to identify contaminants and also the sources of contamination. Carbapenem resistance is apparently at an early stage of dissemination, making even more important the follow-up of its evolution on aquatic environments.
Footnotes
Acknowledgments
This work was supported by European Funds (FEDER) through COMPETE and by National Funds through the Portuguese Foundation for Science and Technology (FCT) within project PEst-C/MAR/LA0017/2013 and grant SFRH/BD/43468/2008 (M.T.). It was also supported by the Observatoire Hommes-Milleux, Estarreja through project REFRESH (minimizing microbial hazards in vegetables cultivated in the Estarreja municipality). I.S.H. was supported by ESF (EU) and POPH funds (Programa Investigador FCT).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.