
Abstract
The complete sequences of two previously reported plasmids carrying plasmid-mediated quinolone resistance genes from Shigella flexneri in China have not been available. The present study using the p5-C3 assembly method revealed that (1) the plasmid pSF07201 with aac(6′)-Ib-cr had 75,335 bp with antibiotic resistance genes CTX-M-3, TEM-1, and FosA3; (2) seven fragments of pSF07201 had more than 99% homology with the seven corresponding plasmids; (3) the other plasmid pSF07202 with qnrS had 47,669 bp with antibiotic resistance gene TEM-1 and 99.95% homology with a segment of pKF362122, which has the qnrS gene from location 162,490 to 163,146. A conjugation and electrotransformation experiment suggested that these two plasmids might horizontally transfer between and coexist in Escherichia coli J53 and S. flexneri 2a 301. Either the aac(6′)-Ib-cr or qnrS gene contributed to, but only the coexistence of the two genes conferred to the resistance to ciprofloxacin in these two strains. To the best of our knowledge, this is the first report of the complete sequences of the aac(6′)-Ib-cr- and qnrS-positive plasmids in Shigella isolates. Our findings indicate that two genes probably evolve through horizontal plasmid transfer between the different bacterial types.
Introduction
B
In our previous study, we reported seven fluoroquinolone-resistant Shigella isolates that were qnrS- and/or aac(6′)-Ib-cr-positive in China from 1998 to 2013. 21 Among the seven isolates, six were found to harbor the qnrS gene, three the aac(6′)-Ib-cr gene, and two both the qnrS and aac(6′)-Ib-cr genes.
The complete sequences of plasmids containing qnr or aac(6′)-Ib-cr have been reported in Escherichia coli, S. enterica, and Klebsiella pneumonia (GenBank: JX997935, LK056646, CP008933, KF362122, JQ60935, CP009115, etc.). However, only the partial sequences of aac(6′)-Ib-cr- and qnr-positive plasmids in Shigella isolates have been available (Genbank: EF501990 and AB187515). To the best of our knowledge, the complete sequences of aac(6′)-Ib-cr- and qnr-positive plasmids in Shigella isolates have not been reported. The aims of this study were to completely sequence and reveal the evolution of PMQR plasmids harboring the qnrS or aac(6′)-Ib-cr gene, to study the location of the quinolone resistance genes aac(6′)-Ib-cr and qnrS in the plasmid scaffold, and to characterize plasmids harboring PMQR genes responsible for antibiotic resistance in Enterobacteriaceae isolates.
Materials and Methods
Bacterial strains and plasmids
The wild-type isolate of S. flexneri 4c 072 (strain 13 in our previous study 20 ), S. flexneri 2a 301 (NC_004337), 13 and E. coli J53 were used in this study. Strain S. flexneri4c 072, harboring the PMQR genes qnrS and aac(6′)-Ib-cr, was isolated in 2007 and is resistant to antibiotics such as nalidixic acid, ciprofloxacin, levofloxacin, ampicillin, norfloxacin, aztreonam, cefotaxime, cefalotin, kanamycin, and tetracycline. The pSF07201 and pSF07202 plasmids used for sequencing were originally from strain S. flexneri 4c 072. The E. coli J53 strain was provided by Dr. Yunsong Yu (Zhejiang University, China). The wild-type strain of S. flexneri 2a 301 was provided by the Chinese Center for Disease Control and Prevention, Beijing, China.
Complete nucleotide sequencing of pSF07201 and pSF07202
Plasmid sequences were determined by PacBio sequencing. For the purpose of a de novo assembly, we carried out both Personal Genome Machine 200 bp shotgun sequencing and 3K mate-pair sequencing with P5 DNA polymerase-C3 chemistry. Sequences were checked, trimmed, and assembled in CLC Bio Genomics Workbench 6.0 (CLC Bio). Trimming using quality scores was limited to 0.05; trimming of ambiguous nucleotides was set to 2; removal of 5′- and 3′-terminal nucleotides was set to 3; and discarded reads was set to a length of 15 bp. The de novo assembly was performed using all trimmed reads. The graph parameter of word-size values was varied from 25 to 35 bp, and the bubble sizes were set to 50, 60, or 70 bp. The map reads back to contigs (slow) parameters and was as follows: mismatch cost of 2, insertion cost of 2, deletion cost of 3, length fraction of 0.5, and similarity fraction of 0.8. The best word size and bubble size were 29 and 60 bp, respectively.
The de novo scaffolds generated by hybrid assembly were linked up and the degenerate nucleotides were resolved according to the following strategy: (1) Two de novo scaffolds were linked up if they were aligned and bridged by more than two mate pair reads; (2) The degenerate nucleotides (N's) were replaced with the corresponding region of the contig if these contigs overlapped with the flanking regions of N's; (3) The N's were removed if the flanking regions aligned to contiguous regions in the contig set. The complete genome sequence obtained using this assembly strategy consisted of a chromosome scaffold of 3.5 Mb in length (average coverage 261) and a plasmid scaffold of 110.5 kb (average coverage 160).
Gene prediction analysis
Open reading frames in the P5 draft genome were predicted using the Rapid Annotation by Subsystem Technology pipeline. 18 The antibiotic resistance genes were searched using the Antibiotic Resistance Genes Database (ARDB) and the Comprehensive Antibiotic Resistance Database (CARD). The resulting sequence data were assembled using the BLAST tools available at the National Center for Biotechnology Information (www.ncbi.nlm.nih.gov). The map was drawn using the program OrganellarGenomeDRAW. 17 The gray arrows indicate the direction of transcription for the two DNA strands. The circle inside the GC content graph marks the 50% threshold.
Conjugation and electrotransformation of pSF07201 and pSF07202 plasmids
To test the transferability of the qnrS- and aac(6′)-Ib-cr-positive plasmids, conjugation experiments were conducted using S. flexneri 4c 072 harboring the qnrS and aac(6′)-Ib-cr genes as the donor strain and azide-resistant E. coli J53 as the recipient strain. Transconjugants (J53pSF07202) were selected on Luria Broth (LB) agar plates containing sodium azide (150 μg ml−1) and ciprofloxacin (0.05 μg ml−1). Transconjugants (J53pSF07201) were selected on LB agar plates containing sodium azide and kanamycin (50 μg ml−1). The presence of the qnrS gene in strain J53 pSF07202 and the aac(6′)-Ib-cr gene in strain J53 pSF07201 was verified by polymerase chain reaction (PCR) using primers specific for the qnrS and aac(6′)-Ib-cr genes, 20 respectively. The plasmids pSF07201 and pSF07202 from the strains J53pSF07201 and J53pSF07202, respectively, were extracted using the Pure Plasmid Midi Kit (Qiagen), according to the manufacturer's instructions. The plasmids pSF07201 and pSF07202 were confirmed to contain the genes qnrS and aac(6′)-Ib-cr, respectively, by PCR, using primers specific for qnrS and aac(6′)-Ib-cr, respectively. 20
The plasmid pSF07201 or pSF07202 was electrotransformed into S. flexneri 2a 301. The electrotransformants containing pSF07201 were selected on LB agar plates with kanamycin (50 μg ml−1). The electrotransformants containing pSF07202 were selected on LB agar plates with ciprofloxacin (0.05 μg ml−1). The electrotransformants containing both pSF07201 and pSF07202 plasmids were obtained by electrotransformation of the pSF07201 plasmid into J53pSF07202 or F301pSF07202. The electrotransformants were selected on LB agar plates containing kanamycin (50 μg ml−1). The electrotransformation was performed in a MicroPulser (Bio-Rad), according to the manufacturer's instructions.
Analyses of antimicrobial activity
The antimicrobial susceptibility of S. flexneri 2a 301, E. coli J53, and their derivatives of pSF07201 and/or pSF07202 plasmids to nalidixic acid, ciprofloxacin, levofloxacin, ampicillin, norfloxacin, aztreonam, cefotaxime, cefalotin, cefepime, kanamycin, tobramycin, streptomycin, amikacin, fosfomycin, cefoxitin, chloromycetin, imipenem, gentamicin, and tetracycline were analyzed by the disc diffusion method in accordance with the Clinical and Laboratory Standards Institute (CLSI) guidelines (Table 1). The interpretive criteria were according to the CLSI criteria for Enterobacteriaceae. 5 The minimum inhibitory concentrations (MICs) of ciprofloxacin for S. flexneri 2a 301, E. coli J53, and their derivatives were determined using the broth microdilution method. E. coli ATCC 25922 was used for quality control. All assays were repeated at least three times.
pSF07201: aac(6′)-Ib-cr-positive plasmid.
pSF07202: qnrS-positive plasmid.
I, antibiotic intermediary; R, antibiotic resistance.
Nucleotide sequence accession number
The complete sequences of pSF07201 and pSF07202 were submitted to the GenBank nucleotide sequence database under the accession numbers KJ201887 and KJ201886, respectively.
Results
Complete sequences of pSF07201 and pSF07202
The aac(6′)-Ib-cr-positive plasmid was found to be 75,335 bp in length (Fig. 1) and to contain four antibiotic resistance genes: aac(6′)-Ib-cr (Location:144–743), CTX-M-3 (Location: 8294–9169), TEM-1 (Location: 6775–7512), and FosA3 (Location: 5,213–5,629). The NCBI BLAST results demonstrated that the complete sequence of pSF07201 could be divided into seven fragments, which had more than 99% homology with the seven plasmids in Table 2, respectively. The 54,159 bp fragment (Location: 18,811–72,960) had 99.96% homology to most of the regions from location 28,487–76,626 and 1–6007 of the plasmid pHN3A11 (JX997935, 76,626 bp) (Table 2). The plasmid pLK056646 (Location: 156,386–156,985) contained the gene aac(6′)-Ib-cr. The plasmid pSF07202, which harbored the qnrS gene, was found to be 47,669 bp in length and to contain the two antibiotic resistance genes qnrS (Location: 38,905–39,561) and TEM-1 (Location: 44,981–45,862) (Fig. 2). The plasmid pSF07202 had the highest homology with the plasmid pEQ2 (KF362122, 287,616 bp; qnrS location: 162,490–163,146) segment from location 119,868–167,552 in E. coli strain 63743, sharing 99.95% sequence homology (Table 2).
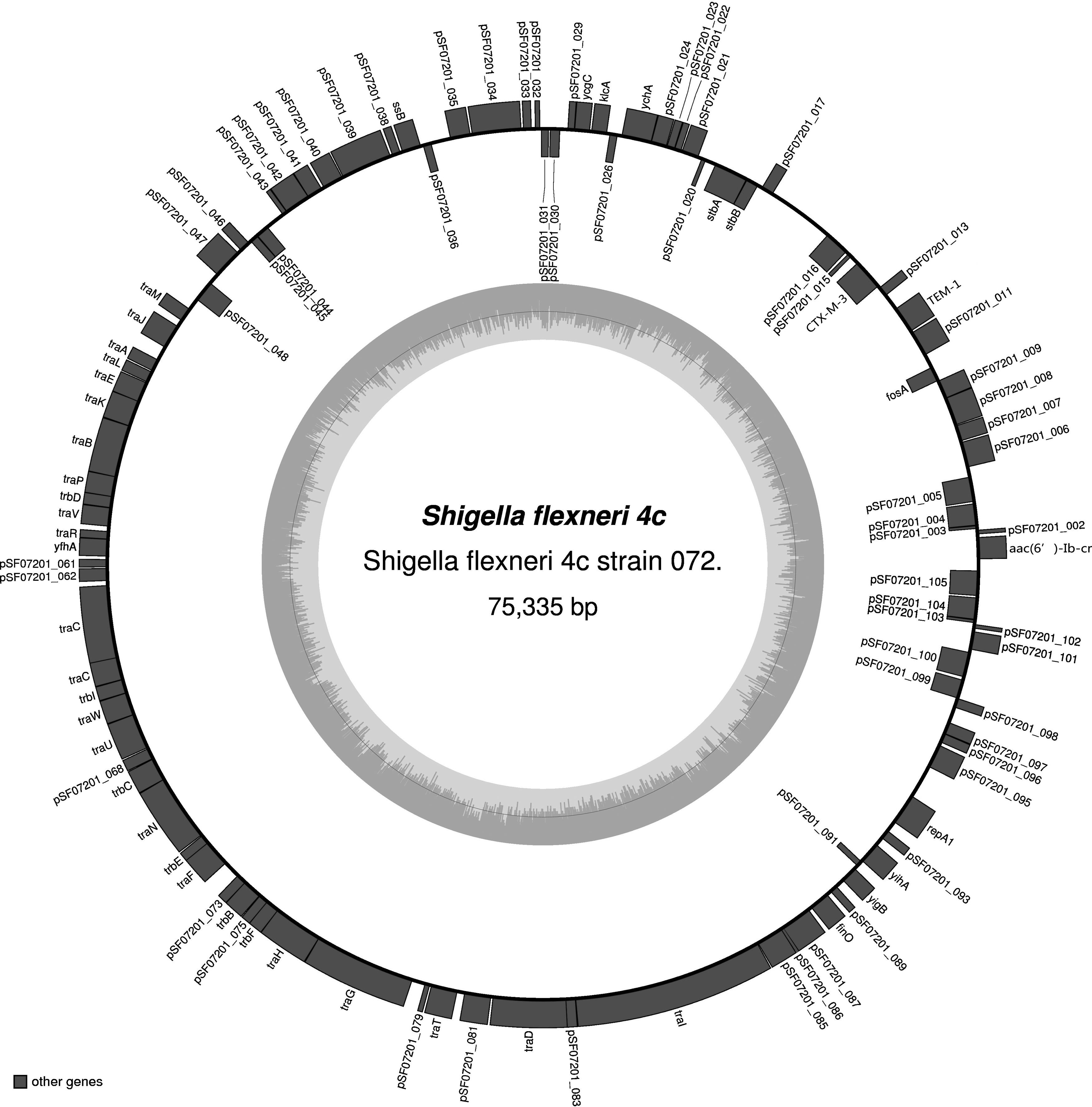
Map of the Shigella flexneri 4c strain 072 plasmid PSF07201 genome.
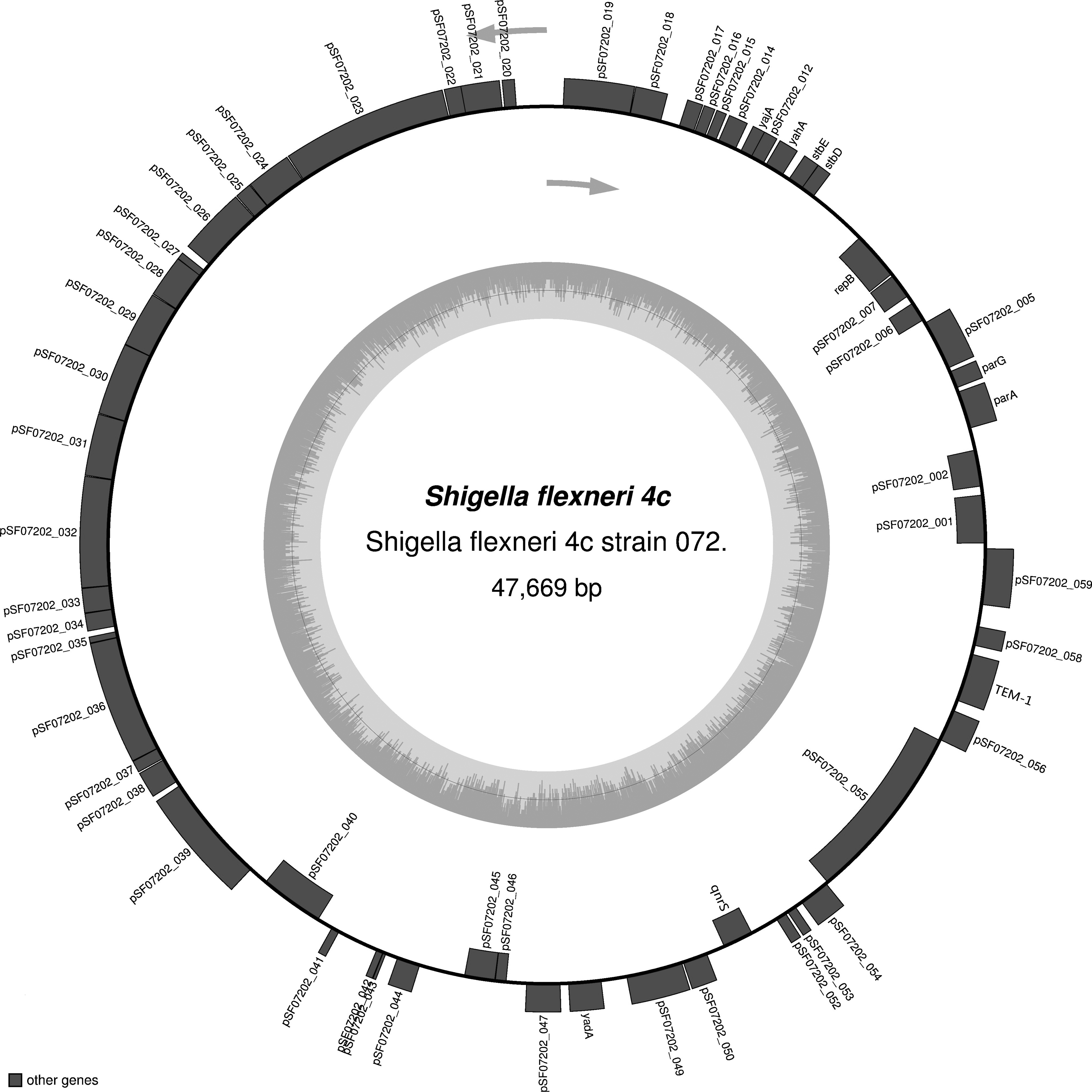
Map of the S. flexneri 4c strain 072 plasmid PSF07202 genome.
Transferability and compatibility of plasmids pSF07201 and pSF07202
Conjugation experiments demonstrated that both the plasmids pSF07201 and pSF07202 were successfully transferred to E. coli J53 and electrotransformed into S. flexneri 2a 301. The two plasmids coexisted compatibly in the same E. coli or Shigella strain.
Plasmids pSF07201 and pSF07202 contributed to antimicrobial activity
Next, we examined whether the plasmids pSF07201 and pSF07202 affected antibiotic resistance (Table 1). The pSF07201 plasmid in E. coli J53 and S. flexneri 2a 301 mainly enhanced the resistance to ampicillin, aztreonam, cefotaxime, cefalotin, cefepime, kanamycin, tobramycin, and fosfomycin, with the resistance to the other 11 antibiotics remaining almost unchanged. The pSF07202 plasmid mainly enhanced the resistance to nalidixic acid, ciprofloxacin, levofloxacin, ampicillin, and norfloxacin, with the resistance to the other 14 antibiotics remaining almost unchanged. The plasmids pSF07201 and pSF07202 together mainly enhanced the resistance to nalidixic acid, ciprofloxacin, levofloxacin, ampicillin, norfloxacin, aztreonam, cefotaxime, cefalotin, cefepime, kanamycin, tobramycin, and fosfomycin, with the resistance to the other seven antibiotics remaining almost unchanged (Table 1).
The MICs of ciprofloxacin in E. coli J53 and S. flexneri 2a 301 transformed with the plasmids pSF07201 and pSF07202 were increased by 4-fold (0.125 vs. 0.03125 μg ml−1) and 32-fold (1.0 vs. 0.03125 μg ml−1), respectively. Therefore, E. coli J53 and S. flexneri 2a 301 transformed with either plasmid pSF07201 or pSF07202 still remained susceptible to ciprofloxacin according to the CLSI standards (MIC ≥4 μg ml−1). The MIC of ciprofloxacin in E. coli J53 and S. flexneri 2a 301 with both plasmids pSF07201 and pSF07202 was increased by 128-fold (4.0 vs. 0.03125 μg ml−1), which almost caused E. coli J53 and S. flexneri 2a 301 to be resistant to ciprofloxacin (Table 2).
+, the plasmid was electrotransferred into the strain; −, the plasmid was not electrotransferred into the strain; MIC, minimum inhibitory concentration.
Discussion
In our previous study, multidrug-resistant Shigella isolates with plasmids harboring the qnrS- and/or aac(6′)-Ib-cr genes were identified. 20 However, these plasmids in Shigella have not been completely sequenced, and the origin of these two genes and whether the plasmids containing either of these two genes can be horizontally transferred between and coexist in different bacterial types has been unknown.
In this study, (1) the complete fluoroquinolone resistance correlative plasmids containing either the aac(6′)-Ib-cr or qnrS gene in S. flexneri were sequenced; (2) the characterization of the antibiotic resistance of two plasmids was performed; (3) the origins of the plasmid pSF07201 and its harboring gene aac(6′)-Ib-cr and the plasmid pSF07201 and its harboring gene qnrS were explored; (4) the two plasmids could be horizontally transferred between and coexisted in E. coli J53 and S. flexneri; (5) without other fluoroquinolone resistance mechanisms, the aac (6′)-Ib-cr or qnrS gene alone contributed to decrease the susceptibility to ciprofloxacin in these two strains, but not to the extent of the resistance level to ciprofloxacin. However, the coexistence of the two genes conferred to the resistance to ciprofloxacin.
The sequencing results indicated that qnrS and aac(6′)-Ib-cr are located in different plasmids, which is consistent with the previous finding that the aac(6′)-Ib-cr gene is not associated with the qnr gene. 19 However, the two genes were found to be compatible when they coexisted in a single bacterial type in this study. The conjugated plasmid containing the aac(6′)-Ib-cr or qnrS gene was successfully transferred from S. flexneri to E. coli J53. In fact, the qnrS1 gene, which harbors in transferable/conjugative plasmids 23 disseminated on different plasmids in several Salmonella serotypes, 11 and Salmonella plasmids bearing qnr alleles are widely spread in Spain. 10 The BLAST results demonstrated that the plasmid pSF07201 might be a plasmid that has recombined different plasmids from E. coli, S. enterica, and K. pneumonia, suggesting that Shigella probably have acquired the novel plasmid pSF07201 by horizontal transmission and recombination among plasmids in Enterobacteriaceae.
In this study, the plasmid pSF07201 was found to contain the antibiotic resistance genes aac(6′)-Ib-cr, CTX-M-3, TEM-1, and FosA3. This result was supported by the following findings. The aac(6′)-Ib-cr gene-encoded protein conferred resistance to tobramycin and kanamycin and decreased fluoroquinone susceptibility; 6 the CTX-M-3 gene is related to cephalosporin resistance; 14 TEM-1 mainly mediates the resistance to penicillin and some first-generation cephalosporin antimicrobial agents and confers sensitivity to third-generation cephalosporin derivatives; 15 and the gene FosA3 correlates with the resistance to fosfomycin in E. coli. 3 In this study, the plasmid pSF07201 in the transformant strain provided resistance to the antibiotics cephalosporin, ampicillin, kanamycin, tobramycin, and fosfomycin; however, the bacteria were not resistant to fluoroquinolone antibiotics. In particular, the plasmid pSF07201 simultaneously carried the PMQR gene aac(6′)-Ib-cr and the ESBL gene CTX-M-3.
The complete sequence of the plasmid pSF07202 had 99.95% homology with one segment of the plasmid pEQ2 of E. coli strain 63743 (KF362122, Location: 119,868–167,552), suggesting that the plasmids pSF07202 and pEQ2 are probably correlated through plasmid horizontal transmission. The plasmid pSF07202 comprises the antibiotic resistance genes qnrS and TEM-1. Therefore, the bacteria containing this plasmid would have decreased sensitivity to quinolone and be resistant to penicillin and some first-generation cephalosporin antibacterial drugs. This qnrS-positive plasmid was different among Enterobacteriaceae. For example, the qnrS-positive plasmid in a clinical K. pneumonia isolate was a 98-kb plasmid conferring resistance to SHV-2 beta-lactamase, aminoglycosides (aacC2, strA, and strB), chloramphenicol (catA2), sulfonamides (sul2), tetracycline (tetD), and trimethoprim (dfrA14). 4 Therefore, the antimicrobial activity confirmed that the bacteria containing the pSF07202 plasmid were resistant to ampicillin and had decreased sensitivity to fluoroquinolone and norfloxacin.
In this study, the aac(6′)-Ib-cr or qnrS gene alone was found to decrease fluoroquinolone susceptibility. In fact, the aac(6′)-Ib-cr and qnrS genes have been identified in many Enterobacteriaceae and may be responsible for high-level resistance to fluoroquinolone.2,19 In addition, three strains of S. flexneri carrying the aac(6′)-Ib-cr gene have shown high levels of fluoroquinolone resistance (MIC of ciprofloxacin: 64 μg ml−1).20,21 The coexistence of the two quinolone resistance genes were also found to almost render either of the two types of bacteria to be resistant to ciprofloxacin, suggesting that the PMQR gene qnrS may be synergistic with and require aac(6′)-Ib-cr gene function. Interestingly, in our previous study, S. flexneri containing mutations at both gyrA83 and parC80, the quinolone resistance-determining region (QRDR) mutations, changed from ciprofloxacin-susceptible to ciprofloxacin-resistant after introduction of a plasmid with qnrS, while the S. flexneri without the QRDR gene mutations remained susceptible, 21 suggesting that the aac(6′)-Ib-cr and qnrS genes probably provide a high level of fluoroquinolone resistance through synergistic effects through other mechanisms such as QRDR gene mutations.
However, we only studied the plasmids isolated from the same strain, whereas different plasmids carrying the aac(6′)-Ib-cr and qnrS genes in S. flexneri may exist. In addition, analyses of the plasmid sequence and origin are complex in reality; therefore, more information regarding the two plasmids should be further obtained based on the present preliminary data in a future study.
Nevertheless, to the best of our knowledge, this is the first report of the complete sequences of the aac(6′)-Ib-cr- and qnrS-positive plasmids from Shigella isolates. Our findings provide information for further PMQR studies and insights into the evolution of qnrS- and aac (6′)-Ib-cr-positive plasmids.
Footnotes
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (No. 81301466), the Zhejiang Provincial Natural Science Foundation (No. LQ12H19001), and the Bureau of Science and Technology of Hangzhou Project of the Zhejiang Province (No. 20110733Q05).
Disclosure Statement
No competing financial interests exist.