
Abstract
Bacteria encoding the New Delhi metallo-β-lactamase gene (blaNDM-1) are regarded as superbugs for their resistance to multiple antibiotics. Plasmids encoding blaNDM-1 have been observed to be spreading among gram-negative bacteria around the world. Previous studies have demonstrated that multiple modifications of blaNDM-1-harboring plasmids might contribute to the spread of the gene. In this study, we analyzed blaNDM-1-encoding plasmids from two Klebsiella pneumoniae isolates, DU7433 and DU1301, found to be unrelated by pulsed field gel electrophoresis and multilocus sequencing typing (DU7433: ST14 and DU1301: ST11), and compared them with previously published plasmids. Although strains DU1301, DU7433, and previously published strain DU43320 carried unrelated plasmids, their transconjugants exhibited similar antimicrobial resistance profiles. Transconjugants lacked the resistance to aztreonam, ciprofloxacin, gentamicin, tetracycline, and trimethoprim/sulfamethoxazole when compared with the corresponding clinical isolates. Plasmids pTR1 from DU1301 and pTR2 from DU7433 had completely different plasmid backbones except a short conserved region of blaNDM-1 and ble flanked with truncated or nontruncated ISAba125 and trpF. The presence of this common region among known blaNDM-1-carrying plasmids implies that the dissemination of blaNDM-1 may be facilitated by mobilization of this conserved immediate region among different plasmids. Control measures should be strictly enforced whenever increasing incidences of epidemiological unrelated strains were identified.
Introduction
N
A previous report in Taiwan has observed that NDM-1-encoding plasmids from Escherichia coli and Klebsiella pneumoniae could be closely related. A common backbone of genetic elements could be identified and categorized by incompatibility (Inc) grouping. 2 In Singapore, two related NDM-1-encoding plasmids were isolated from epidemiologically unlinked patients. 3 Moreover, it is also known that the dissemination of blaNDM-1 occurs predominately by promiscuous plasmids. 14 This plasmid promiscuity may be reflected by gene modification and acquisition events during or after transferring among bacteria.3,9,23 Although the spread of blaNDM-1 may not be explained by an individual mechanism as described above, comparative analysis of existing sequences could suggest a genetic basis for how this encoding gene circulates in a locality. In this study, comparative analysis was performed on two NDM-1-encoding plasmids in Singapore and is further compared with the previously published plasmid from the same locality.
Materials and Methods
Bacterial strains and molecular typing for NDM-1-carrying K. pneumoniae
DU1301 and DU7433 were isolates from patients' urine and were the first blaNDM-1-carrying bacteria to be described in Singapore. 12 Previously sequenced DU43320 was selected for comparison because it was isolated in the same locality. 3
Multilocus sequencing typing and pulsed field gel electrophoresis have been previously performed for all three blaNDM-1-carrying K. pneumoniae strains, and they have been shown to be unrelated (Fig. 1). 11 DU43320, DU1301, and DU7433 belonged to ST273, ST11, and ST14, respectively.
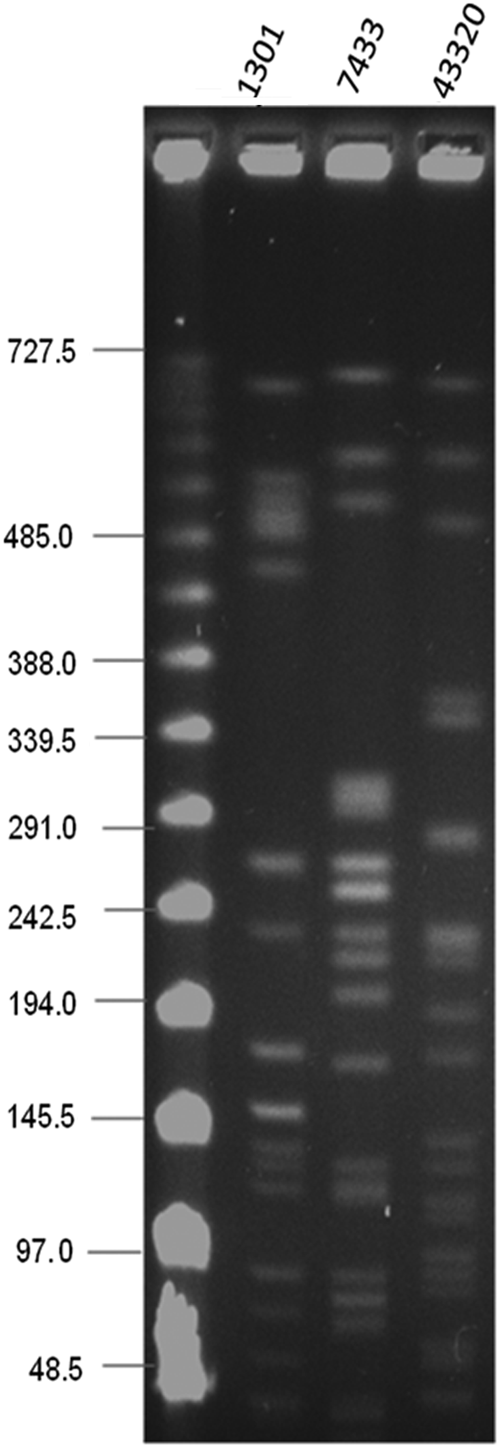
Pulsed field gel electrophoresis of metallo-β-lactamase gene (blaNDM-1)-carrying clinical Klebsiella pneumoniae isolates.
Conjugation of plasmid and antimicrobial susceptibility tests
Plasmid transfer was carried out by conjugation. A rifampin-resistant strain of E. coli (strain JP-995) was used as the recipient. 19 The recipient and donor strains were separately inoculated into brain heart infusion broth (Oxoid Ltd., Basingstoke, England) and incubated at 37°C for 4 hr. The cells were then mixed at a ratio of 1:10 (by volume) and incubated overnight at 37°C. A 0.1 ml sample of the overnight broth mixture was then spread onto a MacConkey agar plate containing rifampin (100 mg/ml) and imipenem (1 mg/ml). We did the polymerase chain reaction for transconjugants to confirm the transfer of blaNDM-1. Only one plasmid was observed in each transconjugants.
Antimicrobial susceptibility tests were performed by the broth microdilution method (Sensititre; Trek Diagnostics, West Essex, England) and E-test.
Plasmid sequencing and analyses
DNA sequencing of these blaNDM-1-carrying plasmids was performed with a whole-genome shotgun approach using 3-kb paired-end libraries. Sequencing was performed using a 454 GS Junior (Roche, Indianapolis, IN). De novo assembly was performed using Newbler 2.6 (Roche). The contigs were manually inspected and reassembled in Consed with additional Sanger reads. 1 The complete nucleotide sequences of plasmid pTR1 and pTR2 have been submitted to GenBank and assigned sequence accession numbers KJ187751 and KJ187752, respectively. Accession numbers for the other plasmids taken to comparison are HQ451074 (pNDM-HK), JQ349086 (pTR3), JQ349085 (pTR4), and AP012055 (pKPX-1).
Results and Discussion
Conjugation of plasmid and antimicrobial susceptibility of isolates and their transconjugants
Antimicrobial susceptibility testing results revealed that strains DU1301 and DU7433 had the same resistance profiles as the previously published strain of DU433203 and were resistant to all tested antibiotics (Table 1). The addition of a β-lactamase inhibitor did not increase the susceptibility to β-lactams. The E. coli transconjugants, TCJ-P2 and TCJ-P3, respectively, from DU1301 and DU7433 showed a loss of resistance to aztreonam, ciprofloxacin, gentamicin, tetracycline, and trimethoprim/sulfamethoxazole when compared with the corresponding clinical isolates. A significant reduced meropenem minimum inhibitory concentration was observed for both transconjugants. When comparing with the transconjugant TCJ-P1 obtained from DU43320, the susceptibility profiles were the same among the three transconjugants.
DU43320 and its transconjugant (TCJ-P1) were used as a control for comparison and were published previously. 3
The minimum inhibitory concentration is presented according to the concentration of trimethoprim.
Genetic features of plasmid pTR1
Sequencing of the blaNDM-1-carrying plasmid from K. pneumoniae strain DU1301 revealed a 31,466-bp circular plasmid named pTR1 (Fig. 2). As indicated in the figure, the plasmid carries three β-lactamase genes, including blaNDM-1, blaCTX-M-15, and a putative β-lactamase gene similar in part to blaOXA-1. A 3.5-kb region spanning the plasmid replication gene, rep, is 99% identical to the repFIIK1 rep region of K. pneumoniae plasmid pKPN-IT from Italy (208,193-bp JN233704) and K. pneumoniae plasmid pKPN3 from the United States (175,879-bp CP000648). The rest of the plasmid backbone of pTR1 is different from those IncFIIK plasmids. Except the rep gene, the majority of the pTR1 sequence, indicated with a thin black line in Figure 2, is very similar to the multidrug resistance region of an 118,033-bp carbapenem resistance plasmid (JN233704) from a K. pneumoniae ST395 strain. 21 This region contained a qnrS1 responsible for reduced susceptibility to fluoroquinolones, and two antimicrobial resistance gene regions associated with insertion sequence (IS) mobile elements. One contained a blaCTX-M-15 β-lactamase gene, the other contained a blaNDM-1 gene adjacent to a bleomycin resistance protein-encoding gene, which is common in blaNDM-1-carrying plasmids. 5 Another antimicrobial resistance region contained a putative β-lactamase gene and is also associated with IS mobile elements. The three β-lactamase gene regions, together with the nearby mobile elements genes and transposase pseudogenes, are identical to those reported in IncH plasmid pNDM-MAR from K. pneumoniae. 22
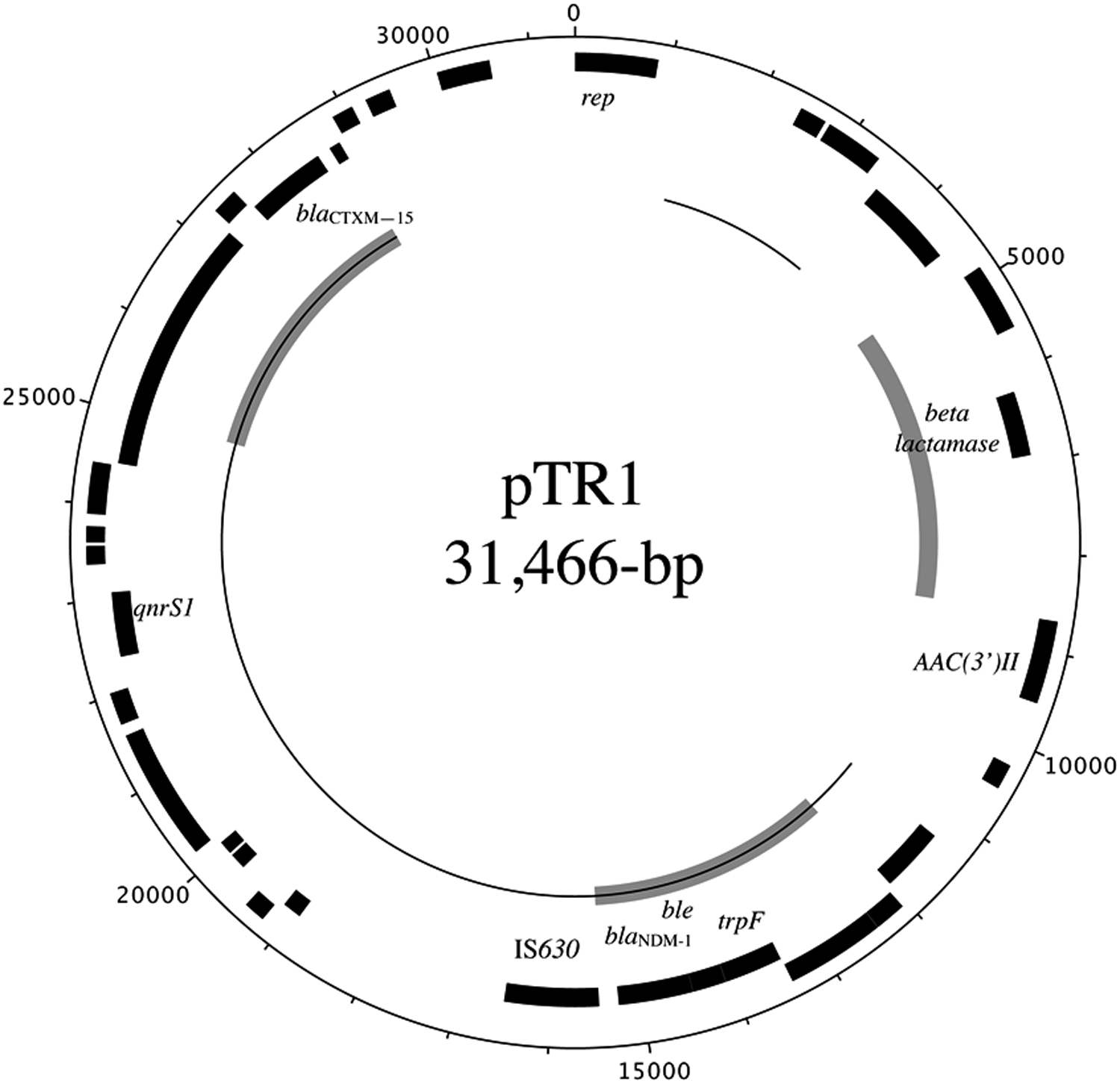
Schematic diagram of plasmid pTR1 from K. pneumoniae DU1301. Black blocks on the outer circles are the coding DNA sequence (CDS) of the positive strand, and the black blocks on the inner circle are the CDS of the negative strand. Annotations for the important CDS are named. The multidrug resistance region similar to plasmid2 from carbapenem resistance K. pneumoniae ST395 was marked with thin black lines inside the circles. The three common β-lactamase gene regions comparable to pNDM-MAR were marked with thick gray lines.
Plasmid pTR2, from K. pneumoniae strain DU7433, is 133,650-bp in length and contained a mosaic multidrug resistance region (Fig. 3). The majority of the pTR2 backbone is similar in part to the IncA/C region of plasmid pTC2 from a Greek Providencia stuartii multiresistant strain 6 (Fig. 3, inner circle). Plasmid pTC2 is 180,184-bp in length and contained two replicon modules (JQ824049). It contained a repA gene and two conjugative transfer regions Tra1 and Tra2 that resembled the IncA/C replication module and also an additional repB-parAB region for the IncR system. 6 The repB region in pTR2 is identical to the IncR region of pTC2, which contained repB and the nearby genes responsible for plasmid partitioning and stability. A 49-kb region flanked by truncated IS26 in pTR2 that contained the repA and Tra2 region was also found identical to pTC2 (Fig. 3). The Tra1 region in pTC2, which contained many of the genes responsible for conjugal transfer, however, is not found in pTR2. The rest of the pTR2 plasmid is a mosaic multidrug resistance region, which contained a blaNDM-1 region similar to pTR1, blaSHV-11 for beta lactamase, sul1 for dihydropteroate synthase, strAB genes for streptomycin resistance, and also a class 1 integron with ere for erythromycin esterase, aadA for aminoglycoside resistance protein, cmlA7 for chloramphenicol resistance protein, and qacEdelta1 for quaternary ammonium resistance (Fig. 3). The plasmid contained two additional replication initiation genes that reside in the mosaic region: a putative DNA replication protein gene, rep, which is identical to the rep gene of plasmid pMDR-ZJ06 from Acinetobacter baumannii MDR-ZJ06 (CP001938), and a repE gene, which is identical to the repE of K. pneumoniae plasmid pK245 (NC_010886).4,25

Schematic diagram of plasmid pTR2 from K. pneumoniae DU7433. Black blocks on the outer circles are the coding DNA sequence (CDS) of the positive strand, and the black blocks on the inner circle are the CDS of the negative strand. Annotations for the important CDS are named. Three regions similar to pTC2 (JQ824049), including Tra2, IncR, and a merR mercury resistance region, were marked with black lines inside the circles. IS26* represents the IS26 pseudogenes flanking the Tra2 region.
The blaNDM-1 gene has been reported on many different plasmid types among enterobacterial isolates. Although on different plasmid backbones, it was reported that the immediate genetic environment of the blaNDM-1 gene is conserved.3,14,15 This conserved region includes an upstream ISAba125, which is usually truncated, and a downstream bleomycin resistance protein (BRPMBL) gene, ble, and trpF (complete or truncated; Fig. 4). The adjacent sequences outside this “blaNDM-1 common region” are quite different in various plasmids and are mostly associated with various transposons and insertion sequences. The adjacent region to the “blaNDM-1 common region” of pTR1 is most similar to pKPX-1 form K. pneumoniae KPX from Taiwan (GenBank: AP012055) 9 and pNDM-MAR from Morocco (JN420336). 22 Downstream of the trpF gene, both pKPX-1 and pNDM-MAR contained genes for groES and groEL chaperonin (Fig. 4). In pTR1, the chaperonin genes were replaced by the putative transposases. The adjacent region to the “common NDM-1 region” of pTR2 is similar to pNDM-HK from Hong Kong (HQ451074). 8 Both pTR2 and pNDM-HK contained a conserved blaDHA-1 pseudogene downstream of trpF. For pTR3 and pTR4, which were also identified from Singapore, the adjacent region to the “common NDM-1 region” is flanked by immediately adjacent ISEc33 (Fig. 4) and putative inverted repeat sequences (JQ349086 and JQ349085). 3 A comparison of such common immediate region, from various blaNDM-1-carrying sequences, is shown in Figure 4.
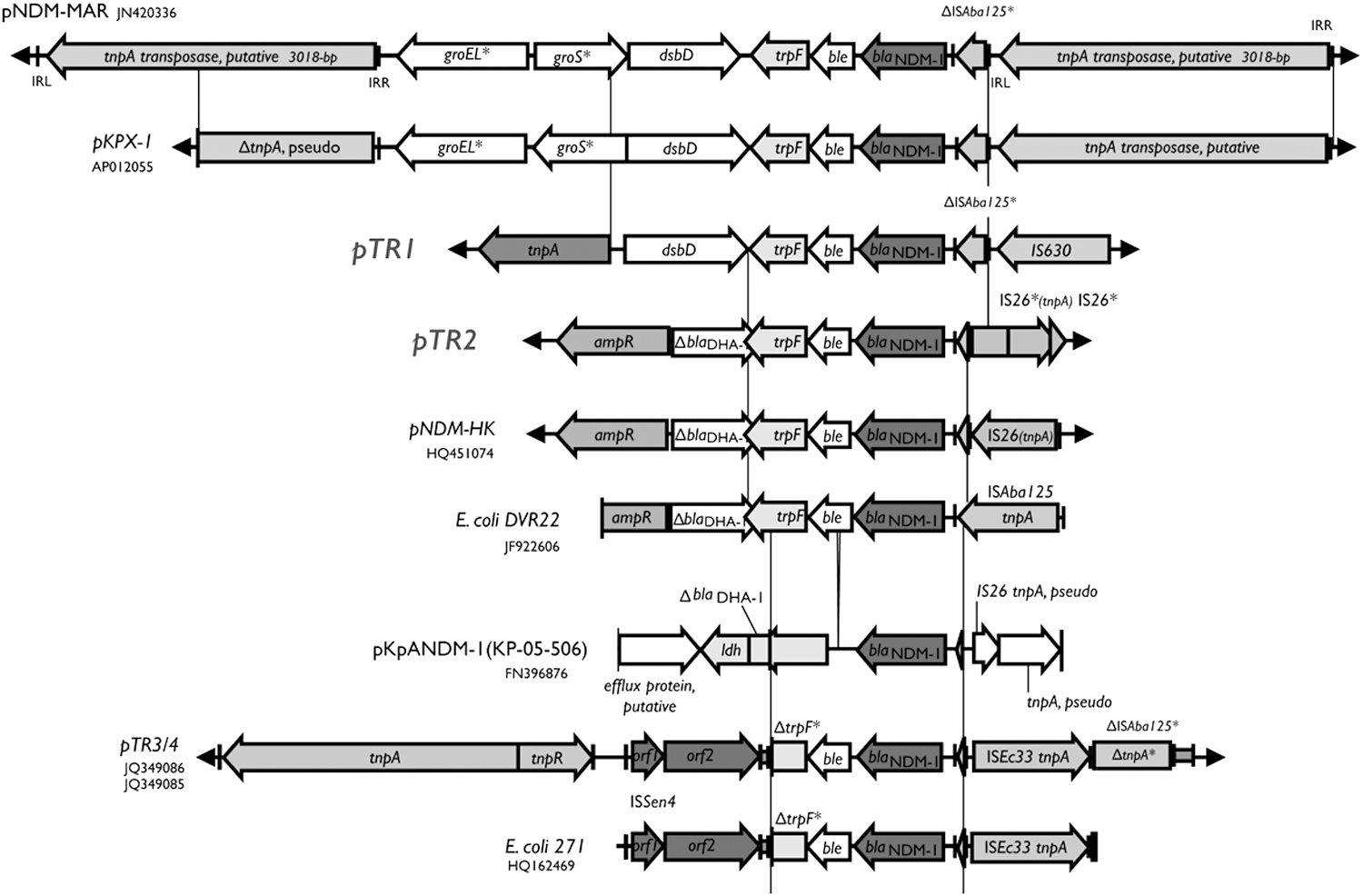
Comparison of the blaNDM-1 common region and their adjacent sequences among several blaNDM-1-carrying plasmids/sequences. The blaNDM-1 (black) and nearby IS elements are shown. Open reading frames are depicted with arrows and the inverted repeats (IRs) were depicted by short vertical bars. Asterisk was applied to the pseudogenes and truncated genes/features. The GenBank accession numbers for each of the blaNDM-1-containing sequences are labeled with the plasmid or strain name.
Previous studies have shown that identical plasmid carrying blaNDM-1 could be found in epidemiological unrelated patients, indicating resistance by plasmids. 3 In the present study, plasmids carrying blaNDM-1 obtained from the same locality were promiscuous. Comparison of published plasmids has shown that only a short region found in the neighborhood of blaNDM-1 is conserved. This conserved region started from ISAba125 to trpF in either truncated or nontruncated form, indicating that the flanking genetic elements are perhaps the hot spot of recombination facilitating the mobilization of the conserved blaNDM-1 region. 7 The movement of these genetic flanking elements may explain the dissemination of blaNDM-1 without apparent epidemiological linkage in the same locality.
In conclusion, comparative plasmid analysis suggests that the dissemination of blaNDM-1 probably occurs through the mobilization of a small common region, which makes tracing the source of dissemination in a particular geographical location difficult. Control measures should be strictly enforced whenever increasing incidences of blaNDM-1-carrying strains are observed even if the strains are apparently epidemiologically unrelated.
Footnotes
Acknowledgments
This work was supported by grants from the Ministry of Science and Technology and the National Health Research Institutes, Taiwan (MG-104-PP-17). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the article.
Disclosure Statement
No competing financial interests exist.