
Abstract
The aim of this study was to evaluate the presence of Vibrio isolates recovered from four different fish pond facilities in Benin City, Nigeria, determine their antibiogram profiles, and evaluate the public health implications of these findings. Fish pond water samples were collected from four sampling sites between March and September 2014. A total of 56 samples were collected and screened for the isolation of Vibrio species using standard culture-based methods. Polymerase chain reaction (PCR) was used to confirm the identities of the Vibrio species using the genus-specific and species-specific primers. Vibrio species were detected at all the study sites at a concentration on the order of 103 and 106 CFU/100 ml. A total of 550 presumptive Vibrio isolates were subjected to PCR confirmation. Of these isolates, 334 isolates tested positive, giving an overall Vibrio prevalence rate of 60.7%. The speciation of the 334 Vibrio isolates from fish ponds yielded 32.63% Vibrio fluvialis, 20.65% Vibrio parahaemolyticus, 18.26% Vibrio vulnificus, and 28.44% other Vibrio species. In all, 167 confirmed Vibrio isolates were selected from a pool of 334 confirmed Vibrio isolates for antibiogram profiling. The susceptibility profiles of 20 antimicrobial agents on the isolates revealed a high level of resistance for AMPR, ERYR, NALR, SULR, TMPR, SXTR, TETR, OTCR, and CHLR. The percentage of multiple drug resistance Vibrio isolates was 67.6%. The multiple antibiotic resistance index mean value of 0.365 for the Vibrio isolates found in this study indicated that the Vibrio isolates were exposed to high-risk sources of contamination when antibiotics were frequently used. The resistant Vibrio strains could be transmitted through the food chain to humans and therefore constitutes a risk to public health.
Introduction
T
Antimicrobial agents are included in fish feed and water, principally to treat and prevent diseases in fish farming.4,26 As a result, antimicrobial agents persist in sediment and aquatic milieu, leading to deteriorating environmental conditions and enabling the development of antimicrobial resistance in bacteria.32,46 The control of antibiotic use in aquaculture varies from country to country, as reported by Alderman and Hastings 2 and Cabello et al. 4 In developed countries, there is a limited number of products, and regulatory control is resilient; furthermore, the use of antibiotics is declining as a result of improvements in the management and development of vaccines.28,52 Bondad-Reantaso et al. 3 noted that 90% of aquaculture production occurs in developing countries where regulatory controls are weak and compromised, and the use of antibiotics appears to be widespread. The rise of antibiotic resistance is considered to be closely linked with the widespread use of antibiotics in human health and animal production. In particular, more than one-half of the antibiotics used in developed and developing countries are administered to livestock for the purpose of promoting growth or treating infections. However, fish pond facilities are considered reservoirs for antibiotic-resistant bacteria, thereby increasing the risk of the transfer of resistance determinants associated with the consumption of food-producing animals to humans.31,32 As with other dangerous pollutants that spread to the environment and threaten human health, there is a need for environmental research to help address the critical problem of microbial resistance to antibiotics. 42 The extensive use of antibiotics on food animal farms in some countries has resulted in the emergence of resistance among food-borne pathogens, opportunistic pathogens, and commensal flora of food animals and has significantly contributed to the development of antibiotic resistance in human commensal flora.4,29
Vibrio species inhabit aquatic environments at temperatures ranging from 10°C to 30°C.17,26 Several species (e.g., Vibrio vulnificus, Vibrio harveyi, Vibrio parahaemolyticus, Vibrio alginolyticus, Vibrio anguillarum, and Vibrio splendidus) have been found to induce infections in aquaculture systems.21,26 These species are known to be environmental microorganisms autochthonous to the aquatic milieu that can be sporadically transmitted to humans through unhygienic food-producing animals or contaminated water sources, which is considered an important vehicle for Vibrio spp. infections. The aim of this study was to evaluate the presence of Vibrio isolates in several fish pond facilities in Benin City, Nigeria; we then characterized these isolates, determined their antibiogram profiles, and evaluated the public health implications of our findings.
Materials and Methods
Sample collection and isolation of presumptive Vibrio isolates
Four fish pond facilities, two earthen ponds (Farm A and B), and two concrete ponds (Farm C and D), were selected in the Ikpoba-Okha Local Government Area in Benin City, Edo State, Nigeria. Because of confidentiality considerations, the sampling sites were designated as Farms A–D. All selected fish ponds discharge their untreated effluents directly into nearby rivers. Fish pond water samples were obtained monthly between March and September 2014. Samples were collected in sterile 1,000-ml Nalgene bottles and transported on ice from the sampling site to the laboratory for analyses. Samples were serially diluted and concentrated on nitrocellulose membrane filters (0.45-μm pore size; Millipore) by passing 100 ml of each dilution through the filter using the membrane filtration technique. The filters were then placed onto agar plates containing thiosulfate citrate bile salts sucrose agar (TCBS agar). The spread plate technique was also employed by spreading 100 μl volumes of effluent samples onto TCBS agar as previously described by Igbinosa et al. 20 Yellow and green colonies were considered Vibrio colonies presumptively and counted as described in Igbinosa et al.18,20 Presumptive Vibrio colonies were then isolated and subjected to Gram staining and oxidase testing. The Gram-negative and oxidase-positive isolates were selected for further characterization.
Molecular identification
Polymerase chain reaction (PCR) was used to confirm the identities of the Vibrio species using the genus- and species-specific primers stated in Table 1. V. parahaemolyticus DSM 11058, Vibrio fluvialis DSM 19283, and V. vulnificus DSM 11507 were used as positive control strains. Deoxyribonucleic acid (DNA) extraction and PCR were carried out as described in Igbinosa et al. 20 with modification. Single colonies of the presumptive Vibrio isolates grown overnight at 37°C on brain–heart infusion (BHI) agar plates were picked and suspended in 200 μl of filtered sterile distilled water, and the bacterial cells were harvested by centrifugation at 11,000 g for 2 min. The pellet was suspended in 100 μl of filtered sterile distilled water and boiled for 15 min at 100°C. The cell lysates (5 μl) were used as a template for the PCR assays immediately after extraction.
Screening for antibacterial susceptibility
The Vibrio isolates that were positively identified to belong to one of the three species and other Vibrio spp. were subjected to antibiogram characterization. All the bacterial isolates were tested for resistance or sensitivity to different antibiotics using the standard disc diffusion method (Kirby Bauer test). For the disc diffusion assay, bacteria were grown between 18 and 24 hr on Mueller-Hinton agar, harvested, and then suspended in 0.85% sterile physiological saline solution adjusted to a 0.5 McFarland turbidity standard, corresponding to 108 CFU/ml. The inoculum was streaked onto plates of Mueller-Hinton agar using a sterile cotton swab and impregnated with appropriate antibiotics. The results were recorded after 24 hr of incubation at 37°C. Commercially available antibiotic discs, obtained from Oxoid (Mast Diagnostics), were used to determine the resistance patterns of the isolates against 20 different antibiotics (one dose/disc), grouped into eight different classes according to their mechanisms of action.
Cell envelope antibiotics
(1) Beta-lactams, including penicillins (ampicillin [AMP, 10 μg], carbenicillin [CAR, 10 μg]), monobactams (aztreonam [ATM, 30 μg]), cephalosporins (cefotaxime [CTX, 30 μg], ceftazidime [CAZ, 30 μg], cefuroxime [CXM, 30 μg]), and carbapenems (imipenem [IMP, 10 μg])
Nucleic acid inhibitors
(2) macrolide (erythromycin [ERY, 15 μg])
(3) quinolones (nalidixic acid [NAL, 30 μg]) and fluoroquinolones (ciprofloxacin [CIP, 5 μg], norfloxacin [NOR, 10 μg])
(4) sulfonamides (sulfamethoxazole [SUL, 25 μg], trimethoprim [TMP, 30 μg]) and potentiated sulfonamides (sulfamethoxazole+trimethoprim [SXT, 25 μg])
(5) DNA inhibitors (nitrofurantoin [NIT, 300 μg])
Protein synthesis inhibitors
(6) aminoglycoside (amikacin [AMK, 30 μg], gentamicin [GEN, 20 μg])
(7) tetracyclines (tetracycline [TET, 30 μg], oxytetracycline [OTC, 30 μg])
(8) phenicol derivatives (chloramphenicol [CHL, 30 μg])
These antibiotics were selected because they are important veterinary antimicrobials. The diameter of the zone of inhibition around each disc was measured and interpreted as Resistant (R), Intermediate resistant (I), or Sensitive (S) in accordance with the recommended standard established by the Clinical Laboratory Standards Institute. 7 Multiple antibiotic resistance (MAR) was defined as the resistance to three or more chemical classes of antimicrobial agents. For all isolates, the MAR index values were calculated as a/b, where a represents the number of antibiotics the isolate was resistant to, and b represents the total number of antibiotics the isolate was tested against. A MAR index value >0.2 is observed when the isolate is from an area exposed to high-risk sources of contamination, where antibiotic use is common. A MAR index value ≤0.2 is observed when antibiotics are rarely or never used. 23
Phenotypic traits of potential virulence factors
Colonies grown on BHI agar were suspended in 3 ml of Mueller-Hinton broth. The density of this suspension was adjusted to 0.5 McFarland standard, which is the equivalent of 108 cells/ml. A 5 or 10 μl sample of this suspension was added to media or substrates for the phenotypic characterization of their virulence factors. The phenotypic characteristics of the 167 Vibrio isolates were assessed using the methods described in Moreno and Landgraf. 33 The hemolysis activity of the isolates was assayed on a sheep blood agar plate at 37°C. The lipase activity of the isolates was assayed on Tryptic soya agar (TSA) plates supplemented with 1% Tween 80 (v/v). The extracellular protease activity of the isolates was assayed on TSA plates supplemented with 1% casein (v/v). The gelatinase production of the isolates was assayed in a gelatin medium (3% beef extracts, 5% peptone, and 15% gelatin, pH 7.0). To test for the DNA degrading activity, the isolates were assayed on DNase agar plates. All strains were tested in triplicate and incubated at 37°C.
Results
Vibrio detection isolated from fish pond facilities
Vibrio species were detected at all study sites on the order of 103–106 CFU/100 ml. The isolated Vibrio species were different for each sampling location (Table 2). The overall correlation between the Vibrio counts for each sampling site was moderate (r = 0.69), but the correlation was strongly variable for the different sites, ranging from strong for the earthen pond (r = 0.84) to moderate for the concrete ponds (r = 0.57). When a randomized sample of the 550 presumptive Vibrio isolates was subjected to PCR confirmation for Vibrio spp., 334 isolates tested positive, giving an overall Vibrio prevalence rate of 60.7%. Overall, 334 isolates from the fish pond facilities were obtained and identified, of which 32.63% were V. fluvialis (n = 109), 20.65% were V. parahaemolyticus (n = 69), 18.26% were V. vulnificus (n = 61), and 28.44% were Vibrio species (n = 95) (Table 2). All the Vibrio spp. isolates were confirmed by PCR (Table 1) using genus- and species-specific primers.
Antimicrobial resistance characterization
Overall, 167 confirmed Vibrio isolates were randomly selected from a pool of 334 confirmed Vibrio isolates taken from across all sampling areas. These isolates were classified into three species and other Vibrio spp. using PCR as described in Table 1. The resistance profiles obtained with the 167 Vibrio isolates against 20 antibiotics are shown in Table 3. All the isolates showed 100% resistance to tetracycline and 99.4% to oxytetracycline. In addition, the highest resistances encountered were 91% to erythromycin, 85% to ampicillin, 83% to sulfamethoxazole+trimethoprim, 73.1% to chloramphenicol, 72.5% to trimethoprim, 68.8% to sulfamethoxazole, and 50.8% to nalidixic acid, while the rest of resistances were under 30% (Fig. 1). In contrast, all isolates were 100% susceptible to ciprofloxacin. The MAR index mean value of 0.365 indicated that the Vibrio isolates were exposed to high-risk sources of contamination where antibiotics were often used. The MAR index also revealed a high risk of multidrug resistance in the Vibrio isolates originating from fish ponds.
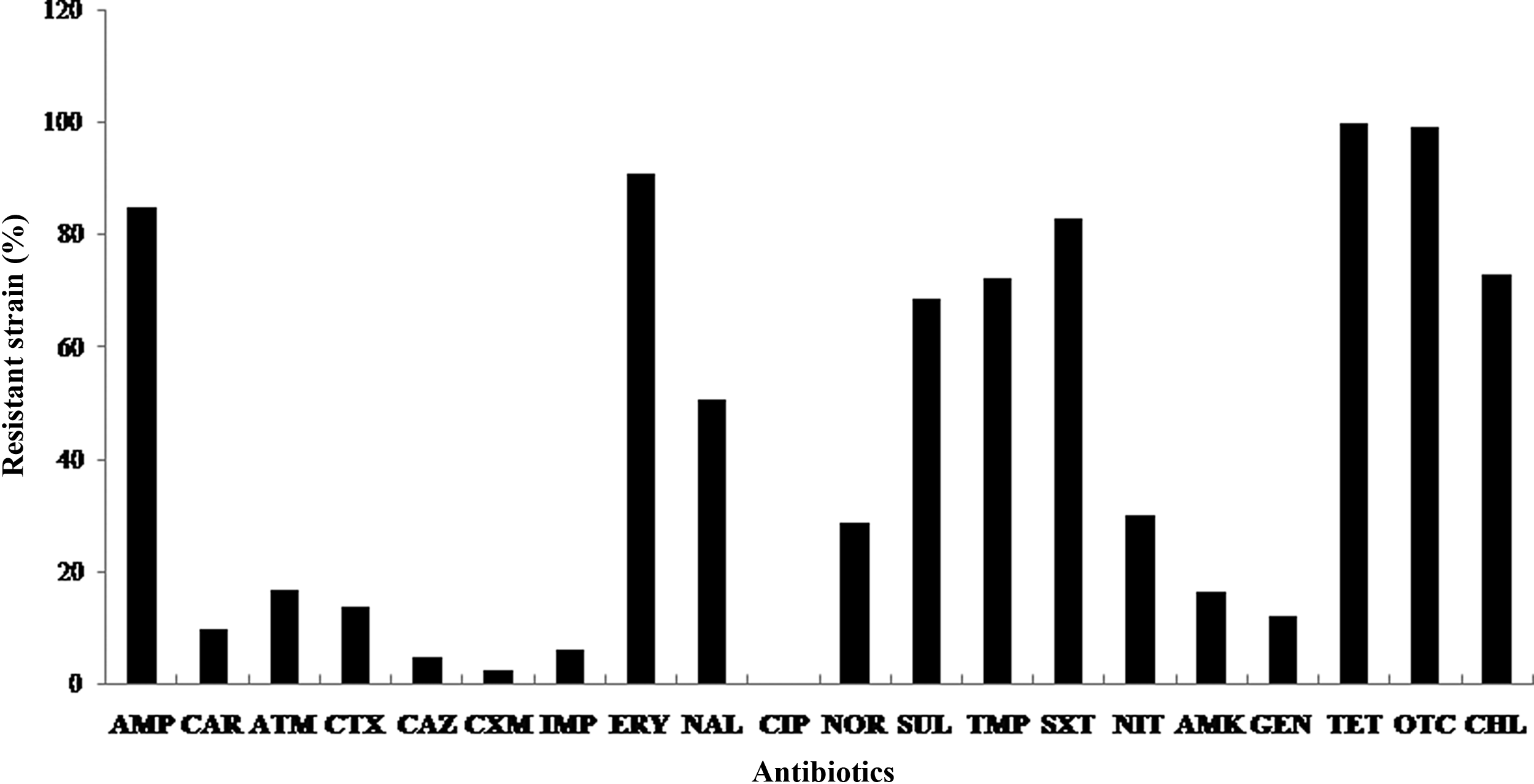
Percentage of the frequency of resistance to antibiotics for Vibrio isolates recovered from fish pond (aquaculture) sources. AMP, ampicillin; CAR, carbenicillin; ATM, aztreonam; CTX, cefotaxime; CAZ, ceftazidime; CXM, cefuroxime; IMP, imipenem; ERY, erythromycin; NAL, nalidixic acid; CIP, ciprofloxacin; NOR, norfloxacin; SUL, sulfamethoxazole; TMP, trimethoprim; SXT, sulfamethoxazole+trimethoprim; NIT, nitrofurantoin; AMK, amikacin; GEN, gentamicin; TET, tetracycline; OTC, oxytetracycline; CHL, chloramphenicol.
AMP, ampicillin; CAR, carbenicillin; ATM, aztreonam; CTX, cefotaxime; CAZ, ceftazidime; CXM, cefuroxime; IMP, imipenem; ERY, erythromycin; NAL, nalidixic acid; CIP, ciprofloxacin; NOR, norfloxacin; SUL, sulfamethoxazole; TMP, trimethoprim; SXT, sulfamethoxazole+trimethoprim; NIT, nitrofurantoin; AMK, amikacin; GEN, gentamicin; TET, tetracycline; OTC, oxytetracycline; CHL, chloramphenicol; R, resistant; I, intermediate; S, susceptible.
Multiple antibiotic resistances
Of the 167 Vibrio isolates, 70.6% (n = 118) showed multiple resistances to the eight classes of antibiotics (Fig. 2). Twelve isolates (10.16%) were resistant to one class of antibiotics, 10 (8.47%) were resistant to two classes, and 30 (25.42%) were resistant to three classes. Fifteen isolates (12.71%) were resistant to four classes, 20 (16.94%) were resistant to five classes, and 16 (13.55%) were resistant to six classes. Eleven isolates (9.32%) were resistant to seven classes and four (3.38%) were resistant to eight classes.

Multiple antibiotic resistances of Vibrio isolates recovered from fish pond sources.
Virulence potential
The phenotypic expression and characteristics related to virulence determination were evaluated for the 167 Vibrio isolates (Table 4). Hemolytic and DNase activity were exhibited by 97% and 97.6% of the isolates, respectively. Extracellular protease, gelatinase, and lipase activities were also detected in 93.4%, 92.2%, and 94% of the isolates, respectively.
Discussion
The use of antimicrobial agents to control livestock infections for prophylaxis and for growth promotion has led to the development of resistant strains of pathogenic bacteria. This issue is a major concern for human and animal health, as it leads to a greater risk of therapeutic failure of standard infection management. The diversity of resistance profiles was revealed in this study, but we were unable to link them with antibiotic use, as there is no register of antibiotic use in aquaculture systems in Nigeria. However, the results are similar to those found in other countries where antibiotics are used in aquaculture environments. Antibiotic resistance has been documented in animal pathogens, in commensal bacteria, and in disease agents that can spread to humans through the food chain.12,26 Resistance can be transmitted through sequential mutations in chromosomal genes or through the acquisition of genetic elements, such as plasmids, bacteriophages, or transposons.27,46 Many pathogenic bacteria can survive antibiotic treatment by the presence of an antibiotic resistance-encoding gene in a plasmid. Monitoring the spread of antibiotic resistance is particularly important for the risk assessment related to Vibrio species, as many of these species are reported to cause fish and shellfish diseases.26,34,48 Once these Vibrio species get into environmental waters, they convert to conditionally viable environmental cells within 24 hr.11,36 Such Vibrio species are infectious when reintroduced into the human body, although the infectious dose in this form is not known. 15
Despite the limited number of bacterial strains that were examined in this study, basic information on the spread of antibiotic resistance within some aquaculture environments was revealed. The results obtained in this study showed that 85% of the examined strains were resistant to beta-lactams (ampicillin). This high resistance could be a result of the production of inducible chromosomal beta-lactamases, as the isolates were 91% resistant to the macrolide (erythromycin) and 50.8% were resistant to quinolone (nalidixic acid). A high number of the isolates was susceptible to other beta-lactams (i.e., carbenicillin, aztreonam, cefotaxime, ceftazidime, cefuroxime, imipenem), as well as to other quinolones (i.e., ciprofloxacin and norfloxacin). The resistance to ampicillin is not surprising, as it agrees with many previous studies performed in a number of countries.6,16,43 The resistance of Gram-negative bacteria to erythromycin is expected because of the intrinsic resistance of many such organisms to macrolide antibiotics. 1 Quinolone resistance has been reported in environmental isolates at a relatively low frequency. 30 However, Chelossi et al. 6 reported that 70% of their isolates were resistant to nalidixic acid. In this study, 50.8% of the Vibrio isolates were resistant to nalidixic acid. Authors have observed differences in antibiotic resistance levels in different production systems, sampling areas, sampling types, and numbers of samples. This resistance poses a great concern because of the risks of cross-resistance to other fluoroquinolones or the development of ciprofloxacin resistance through a second mutation. 8
In contrast, no resistance to ciprofloxacin was observed. Comparing these data to other previous reports, Ferrini et al. 12 reported that 82% of Vibrio isolates isolated from fish settings as well as both national and imported seafood showed resistance to ampicillin, followed by sulfamethoxazole, tetracycline, and trimethoprim/sulfamethoxazole. Only a low percentage of the isolates (ranging from 1% to 7% of the total) were resistant to these additional antibiotics. The Vibrio isolates also showed a high frequency of resistance to sulfonamides (sulfamethoxazole 68.8%), trimethoprim (72.5%), and potentiated sulfonamides (sulfamethoxazole+trimethoprim [83%]), which is possibly mediated by the permeability barrier and/or multidrug efflux system. Resistance to tetracyclines, oxytetracycline, and chloramphenicol has been previously reported.6,16,22,44 In this study, the Vibrio isolates were resistant to tetracyclines (100%), oxytetracycline (99.4%), and chloramphenicol (73.1%). According to Wang et al., 51 antibiotics such as tetracyclines, oxytetracycline, and chloramphenicol are usually added to fodder to stimulate growth because of their low cost and spectrum of action against bacteria. Oxytetracycline is widely used in the treatment of bacterial infections in aquatic livestock. A United Nations publication on the use of antibiotics in aquaculture highlights the risk of using oxytetracycline in aquatic environments. Molecules of oxytetracycline bind to Ca++ and Mg++ ions in the water, thereby significantly reducing the biological activity of the drugs. 10 The occurrence of diseases on fish farms, the alarming increase in antimicrobial resistance, and the use of fodder with added antibiotics require extensive investigation. The establishment of appropriate therapeutic doses of antibiotics can help minimize the potential impacts on the environment and human health. 37
The MAR index mean value of 0.365 indicated that the Vibrio isolates were exposed to high-risk sources of contamination where antibiotics were often used. The occurrence of multidrug resistance strains in the environment could be an indication of the excessive use of antibiotics in aquaculture fields. However, similar MAR values were found in other reports.20,38 The multiple resistance and MAR index values observed for the Vibrio aquaculture isolates worldwide suggest that aquaculture contributes to antibiotic resistance in the environment and thus poses a risk to water quality and human health. Vibrio isolates readily develop single or MAR, suggesting that these isolates may be a good indicator of antimicrobial resistance in freshwater aquaculture. 41 However, the occurrences of antibiotic resistance and multiple resistances are moderately consistent with various reports on fish pathogens and bacteria in aquatic environments.32,50,53 These antimicrobial resistant isolates might be hazardous for public and environmental health. Thus, it is important to continue surveillance and implement preventive control measures concerning antibiotic-resistant bacteria in fish and aquaculture systems.
The presence of putative virulence factors in the Vibrio isolates is an important subject. However, the distribution of virulence factors described is still unknown in strains of environmental origin. The pathogenicity mechanisms of the Vibrio isolates involve both structural and extracellular components. Studies of the bacteria have elucidated a number of products important for its virulence, including cholera toxin, whose action is largely responsible for the host secretory response and toxin-coregulated pili, which greatly enhances its colonization of the intestinal epithelium.25,55 Other factors include potential toxins, accessory colonization factors, outer membrane proteins, proteases, hemolysins, hemagglutinins, and in some strains, a capsular polysaccharide, all of which may contribute to the survival and proliferation of Vibrio species within the host.14,25,55 Hemolysin and various extracellular hydrolytic enzymes are produced in Vibrio species and play important roles in the invasiveness and establishment of infections.25,40 The presence of hemolytic activity may contribute to diarrheal-related virulence. The phenotypic data presented in this study suggest a potential pathogenicity of the isolates from the aquaculture system and a possible risk to human and public health. A V. harveyi strain with high virulence toward salmonids was found to have a high hemolytic activity in fish erythrocytes and contained two copies of the hemolysin gene. 56 Other hemolytically active components such as hemolysins have also been shown to have cytotoxic, enterotoxic, and cardiotoxic activities.
Proteases constitute a second important group of lytic enzymes and include metalloproteases, serine proteases, cysteine proteases, collagenases, caseinases, and gelatinases. Proteases have been reported in V. alginolyticus, V. harveyi, and V. parahaemolyticus and have been found to digest a range of host proteins, including gelatin, fibronectin, and collagen.25,40 Similar to hemolysins, proteases have been linked to virulence toward both shrimp and fish. 25 Many extracellular proteases have been suggested to play important roles in the virulence of Vibrio spp. In a study by Lee et al., 25 a protease was identified as the major virulence factor in V. parahaemolyticus isolates, and this enzyme showed a cytotoxic activity in Chinese hamster ovary and Vero cells. 40 Little is known about the phenotypic and genotypic markers. Thus, more studies are needed, but the high incidence of positive results demonstrates the high potential for hazards in the aquaculture settings. Several studies have suggested that biofilms may be important for the survival, virulence, and stress resistance of Vibrio species. 11 Biofilm production in Vibrio species depends on specific genes (flagella, pili, and exopolysaccharide biosynthesis) and regulatory processes (two component regulators, quorum sensing, and c-di-GMP signaling). 54 The data presented herein support the view that Vibrio species are a significant reservoir of potential virulence and exchange of bacterial DNA material in aquaculture. Presently, a series of experiments are in preparation to study the in vitro transfer of mobile genetic elements from environmental Vibrio species to other Vibrio strains occupying the same ecosystem to investigate the possible spread of genetic elements carrying virulence and antibiotic-resistant determinants in aquaculture. The emergence of environmental bacteria with virulence characteristics may constitute a direct concern to public health.
Conclusion
The inappropriate or indiscriminate use of antimicrobial agents in aquaculture systems is a source of antimicrobial-resistant bacteria and, thus, presents a risk to public and environmental health. The findings in this study revealed that aquaculture environments can act as reservoirs for the potential spread of multiresistant bacteria. The high presence of virulence factors and antimicrobial resistance detected in these strains should not be underestimated. Further studies are required to investigate the extent of antibiotic use in aquaculture milieu and to determine the molecular basis of antimicrobial resistance and virulence traits.
Recommendation
The development and spread of strains that are resistant to currently applied antibiotics should result in restrictions on the use of these compounds in aquaculture. Biocontrol methods could rely on the disruption of regulatory mechanisms that control the expression of virulence genes (e.g., quorum sensing) or on the specific inhibition of the expression of certain critical virulence factors (i.e., antivirulence therapy). These characteristics should be considered for the development of future management strategies for bacterial infections in aquaculture.
Footnotes
Acknowledgment
The author thanks the “The World Academy of Science” (TWAS), Italy (Grant No. 14-091 RG/BIO/AF/AC_1-UNESCOFR: 324028575) for providing the financial support for this research.
Disclosure Statement
No conflicts of interest to declare.