
Abstract
Macrolides are often used to treat and control bacterial pathogens causing respiratory disease in pigs. This study analyzed the whole genome sequences of one clinical isolate of Actinobacillus pleuropneumoniae, Haemophilus parasuis, Pasteurella multocida, and Bordetella bronchiseptica, all isolated from Australian pigs to identify the mechanism underlying the elevated minimum inhibitory concentrations (MICs) for erythromycin, tilmicosin, or tulathromycin. The H. parasuis assembled genome had a nucleotide transition at position 2059 (A to G) in the six copies of the 23S rRNA gene. This mutation has previously been associated with macrolide resistance but this is the first reported mechanism associated with elevated macrolide MICs in H. parasuis. There was no known macrolide resistance mechanism identified in the other three bacterial genomes. However, strA and sul2, aminoglycoside and sulfonamide resistance genes, respectively, were detected in one contiguous sequence (contig 1) of A. pleuropneumoniae assembled genome. This contig was identical to plasmids previously identified in Pasteurellaceae. This study has provided one possible explanation of elevated MICs to macrolides in H. parasuis. Further studies are necessary to clarify the mechanism causing the unexplained macrolide resistance in other Australian pig respiratory pathogens including the role of efflux systems, which were detected in all analyzed genomes.
Introduction
P
Macrolide resistance has previously been identified in several bacterial species associated with PRDC in Australia.5,6 In examining the antimicrobial susceptibility of 71 A. pleuropneumoniae, 97 H. parasuis, 51 P. multocida, and 18 B. bronchiseptica, the percentage of isolates with elevated minimum inhibitory concentrations (MICs) for erythromycin was 89%, 7%, 14%, and 94%, respectively.5,6 Additionally, there was tilmicosin resistance (elevated MICs in case of H. parasuis) in 25%, 21%, 0%, and 22% of isolates and tulathromycin resistance in 0%, 9%, 0%, and 0% of these isolates, respectively.5,6
Macrolide resistance has been associated with several genes that confer resistance by target site methylation (erm genes), efflux (e.g., mef or msr genes and AcrAB multidrug efflux system) and drug inactivation (e.g., ere or mph genes).7,8 The most commonly detected macrolide resistance genes in Gram-positive and Gram-negative veterinary pathogens are erm(A), erm(B), erm(C), mef(A), and msr(A). 8 However, these genes were not found in Australian A. pleuropneumoniae, H. parasuis, and P. multocida isolates with elevated macrolide MICs. 9 Furthermore, the genes, erm(42), msr(E), and mph(E), which have been detected in P. multocida and Mannheimia haemolytica from cattle isolated from the United States,10,11 were not detected in the Australian pig isolates. 9
Another important mechanism of resistance to macrolides is mutations in the 23S rRNA and proteins L22 and L4 (encoded by rplD and rplV, respectively) of the bacterial 50S ribosomal subunit. Particular locations have a key role in macrolide binding and thus action on the bacterial cell.12–14 Alterations of specific nucleotides in the conserved binding regions have been associated with many cases of macrolide resistance in clinical isolates. 13 For example, in M. hyopneumoniae, a mutation at position 2058 (Escherichia coli numbering) of the 23S rRNA (A2058G) has been associated with resistance to macrolides. 15
The aim of this study was to identify the genes and mutations associated with the elevated MICs and resistance to macrolides identified in A. pleuropneumoniae, B. bronchiseptica, H. parasuis, and P. multocida5,6 isolates cultured from Australian pigs with respiratory disease by analysis of the whole genome sequences (WGSs) of one isolate from each bacterial species.
Materials and Methods
WGS analysis
Bacterial strains
The four bacterial isolates tested were obtained from Australian pigs during diagnostic disease investigations and submitted to the Microbiology Research Group, EcoSciences Precinct, Department of Agriculture and Fisheries (DAF), Queensland, Australia, for confirmatory identification and/or serotyping. The A. pleuropneumoniae, H. parasuis, and P. multocida isolates (one of each type) had been identified by the relevant species specific PCR,16–18 as previously reported.5–6 The B. bronchiseptica isolate had been identified by 16S DNA sequencing 19 and a BLAST analysis of the sequence (www.ncbi.nlm.nih.gov/BLAST) as previously reported. 5 The antimicrobial susceptibility of these isolates had been determined by an MIC method as previously reoprted.5,6 The isolates HS 3572 (A. pleuropneumoniae), BR 467 (B. bronchiseptica), and PM 1186 (P. multocida) were selected for whole genome sequencing as they showed elevated MIC for erythromycin or resistance to tilmicosin (Table 1). The isolate HS 3572 was also resistant to tetracycline, however, none of nine known tetracycline resistance genes [tet(A), tet(B), tet(C), tet(D), tet(E), tet(G), tet(H), tet(M), and tet(O)] were detected by PCR. 9 Isolate BR 467 was resistant to beta-lactams (ampicillin, ceftiofur, and penicillin) and florfenicol, while, isolate PM 1186 had elevated MIC only for erythromycin. The H. parasuis isolate (HS 3151) was selected for this study as it showed elevated MIC levels to erythromycin, tilmicosin, and tulathromycin (Table 1).
The MICs were considered elevated for erythromycin, except for HS 3411.
This isolate was tetracycline resistant with MIC of 2 μg/ml.
This isolate was resistant to ampicillin, ceftiofur, florfenicol, and penicillin with MICs of 8, 64, 32, and 64 μg/ml, respectively.
Isolates used in the 23S rRNA analysis only.
MICs, minimum inhibitory concentrations.
Sample preparation
The isolates were grown overnight, with A. pleuropneumoniae and H. parasuis grown on BA/SN plates, 20 and B. bronchiseptica and P. multocida isolates on 5% sheep blood agar plates. Genomic DNA was extracted using the blood and tissue DNeasy kit (Qiagen) as per manufacturer's instructions. The DNA quality was assessed and quantified using a Nano Drop (Thermo Fisher Scientific) spectrophotometer. A total of 200 ng of DNA was used for the library preparation and sequencing using Ion Xpress Plus and Ion PGM (Life Technologies), respectively. The sequencing was performed by the Australian Genome Research Foundation (AGRF, The University of Queensland, Australia).
Genome assembly and analysis
Initially, the quality of the raw sequence reads was checked using FastQC (www.bioinformatics.babraham.ac.uk/projects/fastqc/). All the bioinformatic procedures were performed using Geneious 8.0.4 (www.geneious.com). 21 The reads were trimmed to filter out low-quality reads (<20 and >350 nucleotides for all the samples except for BR 467 where <15 and >350 nucleotides were trimmed). The genome of each bacterial isolate was assembled using the Geneious de novo assembler. The assembled consensus of the contiguous sequences (contigs) of each bacterial genome was submitted to Rapid Annotation using System Technology (RAST) v.2.0 (rast.nmpdr.org/), ResFinder (threshold of 98% identity) (http://cge.cbs.dtu.dk/services/ResFinder/), CARD (http://arpcard.mcmaster.ca), and ARG-ANNOT (www.mediterranee-infection.com/article.php?laref_282&titer_arg-annot) for the detection of antimicrobial resistance genes.22–25
The sequence reads of each isolate were mapped to the sequences of the following reference genomes obtained from NCBI (www.ncbi.nlm.nih.gov/nuccore); A. pleuropneumoniae serovar 12 str. 1096 NZ_ADOL01000000, B. bronchiseptica MO149 NC_018829, H. parasuis SH0165 NC_011852, and P. multocida subsp. multocida str. Pm70 NC_002663.
Analysis of the 23S rRNA sequences
The sequences of all the 23S rRNA copies of the sample genomes (six for A. pleuropneumoniae, H. parasuis, and P. multocida; and three for B. bronchiseptica) were aligned with the nucleotide sequence of the 23S rRNA of the reference genomes and with the E. coli K-12 substrain MG1655 (sourced from BioEdit database) for numbering of nucleotides. 10
As a mutation in the 23S rRNA was detected in the H. parasuis isolate HS 3151, a further six isolates were evaluated for this gene. Five H. parasuis isolates with elevated erythromycin, tilmicosin, or tulathromycin MICs and one isolate with low macrolide MICs (Table 1), with varied serovar or geographical profile, 5 were selected and the 23S rRNA gene sequenced. Genomic DNA from these isolates was extracted using Capture Column Kit (Qiagen). The primer sequences used to amplify the 23S rRNA were as follows: DD forward—5′-AAGTTCCGACCTGCACGAAT-3′ and p85 reverse—5′-GGCAAGTTTCGTGCTTAGAT-3′. 11 The PCRs and cycles were based on the published literature with a few modifications. 11 The Taq polymerase enzyme and buffer (100 mM Tris–HCl, 15 mM MgCl2, and 500 mM KCl, pH 8.3) from Roche was used. The annealing temperature was 58°C. The visualization of the 800 bp PCR product by electrophoresis was done using 10 μl of the amplified product and 2 μl of EZ-Vision® dye (Amresco) in an agarose gel (Progen) containing 1% Tris-acetate-ethylene diaminetetraacetic acid buffer. A low mass ladder (Invitrogen) was used to determine the specific band size. The PCR products were purified using Exo-sap-IT (In Vitro Technology) as the per manufacturer's recommendations. The purified PCR products were sequenced and aligned with 23S rRNA sequences of H. parasuis SH0165 reference genome (Accession No. NC_011852), HS 3151, and Escherichia coli strain K-12 substrain MG1655 for nucleotide numbering (Fig. 2).
Analysis of the efflux regulatory acrR gene sequences
The acrR gene sequences of HS 3572 and HS 3151 were aligned with the nucleotide sequences of acrR gene sequence of the reference genomes.
Results
A. pleuropneumoniae (HS 3572)
A total of 1,751,945 reads were obtained from Ion Torrent PGM Sequencing of the sample HS 3572. A total of 9,890 sequences were generated by the Geneious software 21 from the de novo assembly, with a minimum sequence length of 24 nucleotides and a maximum of 83,601 nucleotides. Analysis of the assembled genome in the three programs (ResFinder, ARG-ANNOT, and CARD) identified the gene strA (804 bp, 99.87% identity) that encodes streptomycin resistance, and sul2 (816 bp, 100% identity) (Table 2) that encodes sulfonamide resistance, both contained in the 7,174 nucleotide contig 1. The gene sul2 was detected in another four contigs listed in Table 2. Since these antimicrobial resistance genes have been found together in plasmids,26–29 the nucleotide sequence of contig 1 was compared against the BLAST database (www.ncbi.nlm.nih.gov/BLAST) for possible similarity with plasmids associated with antimicrobial resistance genes. The contig 1 was similar (99% pairwise identity and coverage) to a plasmid previously described in P. multocida HN06 (Accession No. CP003314) 30 and the pIG1 plasmids from P. multocida (Accession No. U57647) 28 and P. aerogenes strain BB1084 (Accession No. JQ319769) (blast.ncbi.nlm.nih.gov/BLAST.cgi). The schematic comparison of contig 1 and HN06 plasmid, and plG1 plasmid with the annotated genes is shown in Fig. 1.
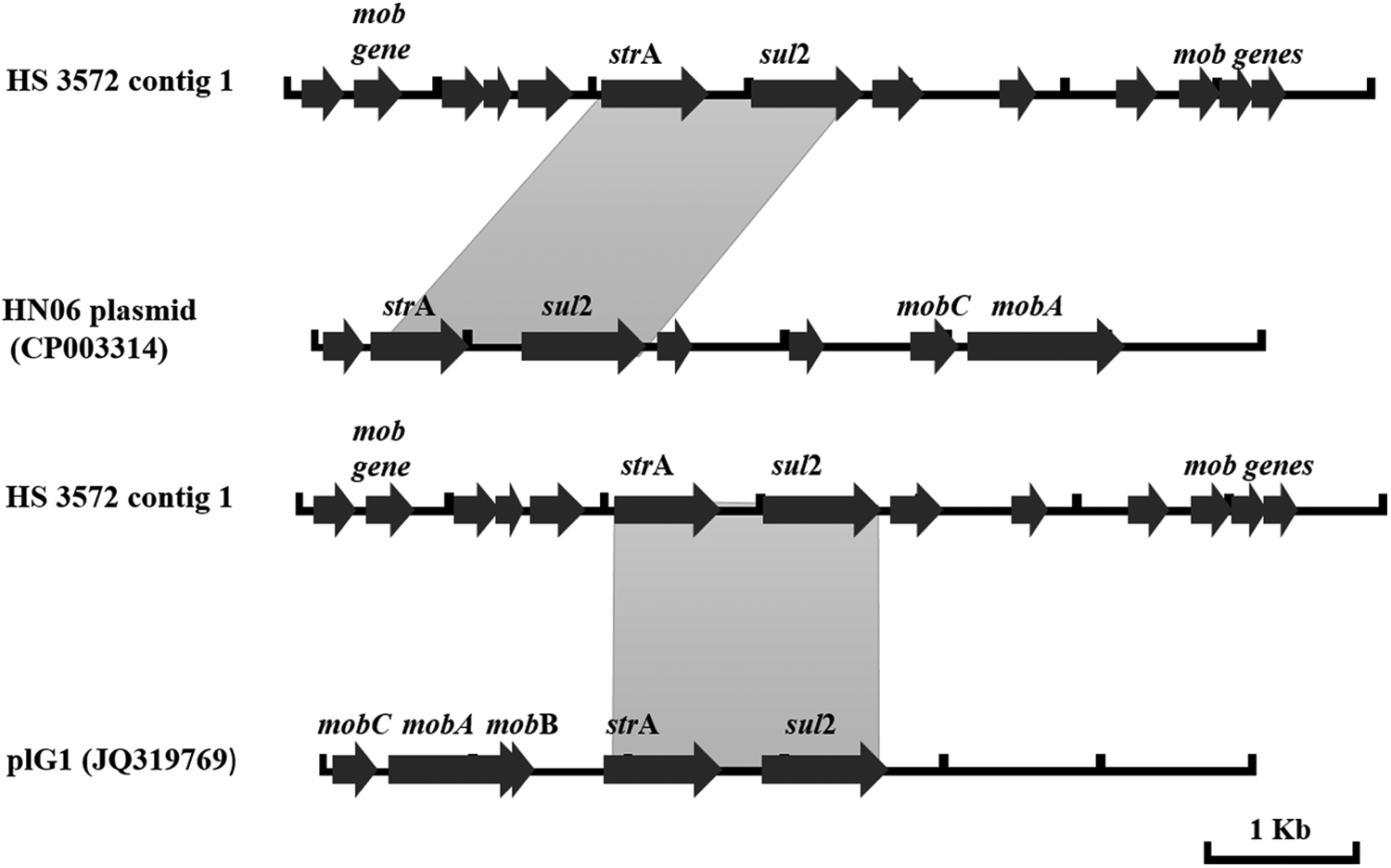
Schematic comparison of HS 3572 contig 1 (7,174 bp) and plasmids HN06 and p1Gl (GenBank Accession Nos. CP003314 and JQ319769 both with 5,360 bp) with the annotated genes. Reading frames are indicated by arrows, with arrowheads showing the direction of transcription. The annotated harbored genes of HN06 plasmid and plG1 are shown—plasmid mobilization genes (mobA, mob, and mobC), aminoglycoside (strA) and sulfonamide (sul2) resistance genes. The annotation for HS 3572 contig 1 was performed using RAST. Gray shades between sequences indicate ≥99% nucleotide sequence identity.
There were no genes found associated with resistance to other antimicrobials.
RAST annotated efflux systems in the assembled bacterial genome (Table 3). These efflux systems have been reportedly associated with antimicrobial resistance when overexpressed. 31 Analysis of the efflux regulator gene, acrR, revealed a C to T nucleotide transition at position 531.
Efflux systems that were found in BR 467 only.
None of clinically important 23S rRNA mutations that have been associated with macrolide resistance12,13,25 were found.
H. parasuis (HS 3151)
There were 2,799,545 reads obtained from the whole genome sequencing of the H. parasuis sample HS 3151. The assembly of these reads created 9,663 contigs, with 500 and 73,442 nucleotides being the minimum and maximum sequence length, respectively. There was no antimicrobial resistance gene detected by the three programs. However, analysis of 23S rRNA revealed a transition of nucleotide A to G at position 2059 (Fig. 2) in six 23S rRNA operons. The 23S rRNA sequences of the additional five H. parasuis isolates with elevated macrolide MICs and one isolate with low MICs did not contain mutations at either position 2058 or 2059, nor in any other significant positions known to confer macrolide resistance.

Alignment of partial sequences of the 23s rRNA of Escherichia coli K-12 substrain MG1655 for the nucleotide numbering (starts at nucleotide number 2051), the reference strain (Haemophilus parasuis SH0165 NC_011852), and H. parasuis (six isolates with elevated macrolide minimum inhibitory concentrations (MICs) and one with low macrolide MICs). At the position 2059 (nucleotide numbering that corresponds to E. coli numbering are shown) each operon of 23S rRNA of HS 3151 has the replacement of nucleotide A by G (underlined). There was no A2059G detected in other H. parasuis isolates.
The RAST analysis of HS 3151 annotated the same efflux systems as those described above for A. pleuropneumoniae HS 3572 (Table 3). Analysis of the acrR gene sequence revealed the following variations from the acrR gene sequence of the reference genome: tandem repeat of nucleotide T at position 316, C411T, T438C, and C480T.
P. multocida (PM 1186) and B. bronchiseptica (BR 467)
De novo assembly generated 4,838 and 603 contigs from 1,422,684 (PM 1186) and 753,028 (BR 467) raw sequencing reads, respectively. There were no known antimicrobial resistance genes detected by the programs in both draft genomes. However, RAST annotated efflux systems were identified in both genome assemblies. Additionally, in the B. bronchiseptica genome, the membrane fusion component of the tripartite multidrug resistance system and multiple antibiotic resistance locus were annotated (Table 3).
The 23S rRNA sequence of P. multocida was 100% identical to the reference genome (Pm70 NC_002663). In B. bronchiseptica, there was also no mutation found at macrolide resistance-associated positions12,13,25 of 23S rRNA sequences.
Discussion
Macrolides are widely used for the treatment of porcine respiratory disease and are categorized as critically important antimicrobials to veterinary medicine by the World Organization for Animal Health. 32 Our previous study did not find a genetic explanation for the elevated MICs and resistance of A. pleuropneumoniae, H. parasuis, and P. multocida Australian isolates to macrolides. 9 In this study, a transition of nucleotide A to G was detected at nucleotide 2059 (E. coli numbering) of the 23S rRNA gene in H. parasuis isolate HS 3151. Mutational changes in 23S rRNA in positions 750, 2057, 2058, 2059, 2452, 2609, and 2611 have been described as conferring macrolide resistance in both Gram-positive and Gram-negative bacteria, altering the function of the 23S rRNA and/or proteins resulting in varied rates of decreased susceptibility to macrolides, lincosamides, and streptogramin. 12 A similar mutation in 23S rRNA has been described in Campylobacter jejuni and Campylobacter coli associated with high erythromycin MIC. 33 M. hyopneumoniae and Mycoplasma gallisepticum, respiratory pathogens of pigs and chicken, respectively, have been reported to carry mutations in nucleotide positions 2058, 2059, 2061, 2447, and 2503 of 23S rRNA gene with A2058G or A2059G mutants exhibiting cross-resistance to the macrolides erythromycin, tilmicosin, and tylosin.15,34
The phenotype conferred by modification of the 23S rRNA target varies according to the mutated base. Mutations at A2058, or at A2059 for certain macrolides, confer the highest levels of resistance to all drugs in macrolides-lincosamides-streptograminB (MLSB) group. 35 Lower-level drug resistance has been reported by mutations at positions 2057, 2452, and 2611, which are close in the secondary structure although slightly outside the focal point of macrolide interaction. 13 Additionally, the rate of resistance depends on the number of mutated copies of the 23S rRNA as bacteria have several copies of this rRNA gene. The finding of the A2059G in the six copies of 23S rRNA of HS 3151 in this study is strongly suggestive that the high macrolide MICs is caused by this mutation. The isolate HS 3151 had the highest tilmicosin and tulathromycin MICs compared with other H. parasuis isolates tested (Table 1). However, the other five isolates did not carry any previously reported point mutations in their 23S rRNA sequences, indicating more than one mechanism of resistance to macrolides in this species. The 23S rRNA mutations are normally passed to daughter cells during replication and are generally not passed between strains or between different genera. 8 As strains containing mutations that confer the highest resistance can be selected with continuous antimicrobial therapy, 4 it would be worthwhile to determine the prevalence of the A2059G in isolates within the same clonal group as HS 3151.
Although the mechanism of macrolide resistance in A. pleuropneumoniae isolate HS 3572 was not identified, we have detected the presence of strA and sul2 genes that have been associated with aminoglycoside and sulfonamide resistance, respectively. These two resistance genes have been found on plasmids (p1Gl plasmid in P. aerogenes [JQ319769] and P. multocida [U57647]; HN06 plasmid in P. multocida [CP003314]; and the pARD3079 plasmid in A. pleuropneumoniae [AM748707]) and are widely distributed among epidemiologically unrelated isolates of the family Pasteurellaceae.28–31 The occurrence of phenotypic resistance to aminoglycosides or to sulfonamide (alone) in strain HS 3572 has not been determined. However, our isolate HS 3572 was found to be susceptible to trimethoprim and sulfamethoxazole (SXT) combination. 5 This suggests that HS 3572 might be phenotypically resistant to aminoglycosides and sulfonamide in which the resistance might be associated with a plasmid carrying the resistance genes. However, future investigation is needed to confirm the presence of plasmid in this isolate.
Additional study is needed to understand the possible cause of elevated macrolide MICs of other isolates including further isolates of A. pleuropneumoniae, P. multocida, and B. bronchiseptica. It is possible that the resistance might have been caused by other resistance mechanism similar to the reported efflux of macrolides and other antimicrobials by multidrug resistant H. parasuis. 7 According to the report of Feng et al. 7 the macrolide resistance in Chinese H. parasuis isolates were associated with the AcrAB multidrug efflux system, a system that is an important cause of multidrug resistance in Gram-negative bacteria. They showed that the deactivation of the resistance-nodulation-division (RND) transporter acrB resulted in increased susceptibility of H. parasuis to antimicrobials including macrolides particularly erythromycin, clarithromycin, and azithromycin. These efflux systems, which were found in the four bacterial isolates included in this study, are innate in antimicrobial susceptible and resistant strains and an overexpression that causes antimicrobial resistance has been described from mutation of regulatory genes. 33 This study found mutations in acrR genes on isolates HS 3572 and HS 3151. Further studies are needed to determine whether such mutations are associated with overexpression of efflux systems contributing to the macrolide resistance of the isolates examined in this study. Moreover, the efflux systems of P. multocida and B. bronchiseptica isolates should also be investigated. Future works may include phenotypic assay such as the technique described by Peric et al. 35 The assay involved the indirect measurement of the radioactive macrolide accumulation in the presence of a protonophore, substance when added to the cell results in rapid dissipation of the electrochemical gradient of protons across the cytoplasmic membrane. 34
This study did not reveal any possible genotypic explanation for beta-lactam and florfenicol resistance in B. bronchiseptica. The beta-lactamase encoding genes such as ampC, blaBOR-1, blaOXA-2, blaCMY-2, and blaTEM-1 have been reported from beta-lactam-resistant strains.37,38 However, low susceptibility to cephalosporins has appeared to be caused by low membrane permeability of B. bronchiseptica and not based on the production of a beta-lactamase.37,38 Florfenicol resistance has been characterized to be conferred by floR gene and might also be due to an unidentified efflux systems. 39 There was also no genetic explanation found in this study for the tetracycline resistance of A. pleuropneumoniae isolate HS 3572. These findings suggest that further work is also needed to explain the underlying cause of phenotypic resistance. These future studies could include an investigation of the cell envelope stress responses, which regulate the assembly of structures including outer membrane porins, pili, and fimbrae in Gram-negative bacteria, and have been previously described to affect susceptibility of B. bronchiseptica to environmental stresses including exposure to beta-lactams. 40
In summary, this study describes that an A2059G transition in the 23S rRNA is likely to be the mechanism that confers high macrolide MICs in one strain of H. parasuis. This acquired resistance mechanism is the first report in H. parasuis with high macrolide MICs. However, further investigation is necessary to elucidate the mechanism behind the unexplained macrolide resistance in other Australian pig respiratory bacterial pathogens.
Footnotes
Acknowledgments
This study was supported by the Australian Pork Cooperative Research Centre project 2A-107 1213 and Australian Infectious Disease Research Centre (AID) for the operating expenses and the Australian Centre for International Agricultural Research project AH/2009/022 John Allwright Fellowship for the PhD scholarship to support D. Dayao.
Disclosure Statement
No competing financial interests exist.