
Abstract
Providencia stuartii has emerged as an important nosocomial pathogen. We describe an outbreak due to a multidrug-resistant strain over a 4-month period in a critical care unit in Athens. Molecular typing revealed each of the isolates to be clonally related with coresistance to cephalosporins, carbapenems, aminoglycosides, and quinolones. Each isolate contained a 220-kb multi-replicon (IncA/C and IncR) conjugative plasmid encoding TEM-1, SHV-5, VEB-1, and VIM-1 β-lactamases and the 16S rDNA methylase RmtB. Antimicrobial therapy was unsuccessful in 3 of 6 cases, and resistance was readily transmissible to susceptible strains of Escherichia coli by transformation and conjugation. This highlights the clinical importance of P. stuartii and its ability to disseminate critical resistance determinants to other bacterial pathogens.
Introduction
T
P. stuartii displays intrinsic resistance to many common antimicrobials, including aminopenicillins, first-generation cephalosporins, polymyxins, and glycylcyclines (tigecycline). It also has inducible (AmpC)-mediated resistance to expanded-spectrum cephalosporins. 24 This limits treatment to carbapenems, aminoglycosides, or quinolones; however, resistance to these agents is increasingly reported. The emergence and dissemination of resistance among Providencia spp. are facilitated by horizontal gene transfer and particularly the acquisition of large conjugative MDR plasmids. 17
As well as being difficult to treat, P. stuartii is frequently involved in nosocomial outbreaks, particularly in nursing homes and burn wound and critical care units. These have often proved difficult to control and impact significantly on patient morbidity, mortality, treatment, and management costs. 44
In this study, we describe an outbreak of pan-drug-resistant (PDR) P. stuartii infections in a Greek intensive care unit (ICU), involving a virulent strain carrying a unique complement of resistance determinants on a large multi-replicon conjugative plasmid.
Materials and Methods
Bacterial isolates and susceptibility testing
Over a 4-month period (December 2012 to March 2013), Providencia stuartii was recovered in BS and UTIs from six patients admitted to a critical care unit in a tertiary care hospital in Athens, Greece. Isolates were identified using the VITEK-2 automated system (bioMerieux, Marcy-L'Etoile, France) and by MALDI-TOF Mass Spectrometry (Bruker, Coventry, United Kingdom). Susceptibility testing was performed using VITEK-2, according to the Clinical and Laboratory Standards Institute (CLSI) Guidelines and minimal inhibitory concentrations (MICs) of aminoglycosides, cephalosporins, carbapenems, colistin, and tigecycline determined by Etest (bioMerieux) on Mueller–Hinton agar. Phenotypic screening for extended-spectrum β-lactamase (ESBL) and carbapenemase production was performed using β-lactam–inhibitor combination disc sets (Total ESBL + AmpC Confirm; ROSCO Diagnostica, Taastrup, Denmark), Modified Hodge Test, and a Rapid CARB Blue Kit (ROSCO Diagnostica). Medical records of the patients were reviewed retrospectively.
Characterization of antibiotic resistance genes
DNA was extracted from exponential phase Luria-Bertani (LB) broth cultures using a Wizard® Genomic DNA Purification Kit (Promega, Southampton, United Kingdom) and quantified using a NanoDrop® Spectrophotometer (NanoDrop Technologies, Wilmington, DE). A screen for β-lactamase genes was performed using Check-MDR CT103 microarrays following amplification, labeling, and detection, according to the manufacturer's protocol (Check-Points, Wageningen, The Netherlands). A series of multiplex polymerase chain reactions (PCRs) were used to confirm and/or determine the presence of genes encoding blaTEM, blaSHV, blaOXA-1/40, blaOXA-48-like, blaCTX-M-groups-1/2/9/8/25, blaACC, blaMOX, blaDHA, blaCIT, blaEBC, blaVEB, blaPER, blaGES, blaIMP, blaVIM, blaNDM, and blaKPC as previously described.9,21
Full coding sequences of all gene fragments detected were amplified and sequenced on both strands using the amplification primers by the Sanger method (Source BioScience, Cambridge, United Kingdom). Consensus sequences were aligned and compared to reference nucleotide sequences curated in the GenBank and the Lahey database (www.lahey.org/Studies/). PCR was also used to search for the presence of genes encoding 16S rDNA methylase (ArmA and RmtA-H) 19 acetyl [AAC(6′)-Ib, AAC(6′)Ih, and AAC(3)-II], nucleotidyl [(ANT(2"-I)], and phosphotransferases [APH(3′)-I and APH(3′)-VI] associated with aminoglycoside resistance. 36 Plasmid-mediated quinolone resistance determinants (qnrA,-D, S, aac(6)′-Ib-cr, oqxAB, and qepA) were sought by multiplex PCR as previously described. 8
Resistance genes contained within integron-like structures were sought using a multiplex PCR for class 1, 2, and 3 integrase genes. 11 PCR mapping sequencing and alignment of products obtained with internal primers were used to determine the content and arrangement of genes within cassettes. All PCRs were carried out using ThermoPrime Plus PCR Master Mix (ABgene, Epsom, United Kingdom), and amplicons were visualized following electrophoresis through 1.5% agarose gels.
Molecular typing
All isolates were analyzed for genetic relatedness by Random Amplified Polymorphic DNA (RAPD) PCR analysis and pulsed-field gel electrophoresis (PFGE). Briefly, RAPD was performed using 50 ng of P. stuartii template DNA and 40 pmol of the oligonucleotide primer ERIC2 (5′-AAGTAAGTGACTGGGGTGAGCG-3′). Thermocycling conditions included a low-temperature (36°C) annealing step, according to the protocol of Renders et al. 39 Amplicons were separated on 1% agarose gels, and patterns were considered indistinguishable if they showed identical banding profiles or if minor differences in the intensity of one or two bands were not confirmed in replicate experiments.
PFGE was carried out on genomic DNA digested with NotI enzyme using a CHEF DR-III apparatus (Bio-Rad Laboratories, Hercules, CA) with the following pulse conditions: initial switch time: 2.2 sec, final switch time: 63.8 sec, voltage: 6 V, included angle: 120°, and running time: 19 hr. XbaI-digested genomic DNA from Salmonella enterica serotype Braenderup strain H9812 was used as a molecular reference marker. 22 Image normalization and construction of similarity matrices were carried out using BioNumerics, version 6.6 (Applied Maths, Sint-Martens-Latem, Belgium), while clones were defined according to the already established criteria. 42
Plasmid analysis and transferability of resistance
Plasmid DNA was extracted from P. stuartii using a modified miniprep method, according to the protocol of Hordijk et al. 20 Purified plasmid DNA was transformed into E. coli DH10B cells (Invitrogen, Van Allen Way, CA) via electroporation under the following conditions: 1.25 kV/cm, 200 Ω, and 25 μFar. 20 Transformants (trf) were selected on LB agar plates supplemented with cefotaxime (1 mg/L), ertapenem (0.5 mg/L), or kanamycin (25 mg/L). On the recovered transformants, the presence of the resistance genes was assessed by PCR, and PCR-based replicon typing (PBRT) was used to identify plasmid replicon types, according to the scheme of Carattoli 7 using a PBRT Kit (Diatheva, Fano, Italy) and the manufacturer's protocol. Transformants obtained for the representative isolate PS70 were subjected to S1-PFGE using the Low-Range PFG Marker (New England Biolabs, Hitchin, United Kingdom) as a size standard to accurately determine the molecular sizes of the plasmids. 5
Plasmid transfer via conjugation was assessed between PS70 (donor) and E. coli CSH26 (recipient). 41 Conjugation experiments were set up by mixing 0.5 ml of exponential phase cultures in 4 ml of LB broth and incubating overnight at 37°C without shaking. Transconjugants (trc) were selected on LB agar plates containing rifampicin (100 mg/L), etrapenem (1 mg/L), and kanamycin (25 mg/L). The conjugation frequency was calculated as the number of trc/donor cell. Resistance genes and plasmids transferred from PS70 to recipients were confirmed by PCR, and replicon typing of transconjugants were recovered. Antimicrobial susceptibility was also determined by Etest (BioMerieux).
Results
Six patients were infected with P. stuartii during the outbreak period, three of whom subsequently died. Four isolates were from BS or catheter-related BS infections and two from complicated UTI. Each patient had a prolonged duration of hospitalization ranging from 24 to 106 days before recovery of P. stuartii. All the patients were nursed in the ICU and received multiple antimicrobial agents (β-lactams, quinolones, and aminoglycosides) before isolation of MDR P. stuartii, three received prior treatment with colistin and one also with tigecycline.
All the isolates were resistant to second-, third-, and fourth-generation cephalosporins, piperacillin/tazobactam, ertapenem, imipenem, aminoglycosides, quinolones, colistin, fosfomycin, and tigecycline but remained susceptible to meropenem (MIC 0.125–0.5 mg/L; Table 1). All the strains were positive for carbapenemase production in a Modified Hodge Test and the acidometric Rapid CARB Blue assay. Combination disc testing suggested coproduction of both an ESBL and a metallo-β-lactamase.
AMP, ampicillin; TAZ, piperacillin/tazobactam; CEPHL, cephalothin; CTX, cefotaxime; CPX, cefpodoxime; CAZ, ceftazidime; FOX, cefoxitin; ERT, ertapenem; IMP, imipenem; MEM, meropenem; GEN, gentamicin; AMK, amikacin; COL, colistin; TGC, tigecycline; MIC, minimal inhibitory concentration.
Checkpoint arrays identified the presence of a wild-type blaTEM allele, blaVIM-1, and a blaSHV-like ESBL (SHV E240K) in every isolate. No pAmpC, blaCTX-M, blaKPC, blaNDM, or blaOXA-48-like genes were detected using the arrays. PCR analysis and sequencing confirmed the presence of genes encoding not only TEM-1, SHV-5, and VIM-1 but also VEB-1 and OXA-10 β-lactamases. The 16S rDNA methylase rmtB and aadA2 encoding pan-aminoglycoside resistance were also found. The latter contained within a 2-kb class 1 integron structure downstream of dfrA12 (trimethoprim resistance). PCR mapping and sequencing of the blaVIM-1 environment localized it to an integron consisting of blaVIM-1, aacA7, dfrA1, and aadA1 identical to the sequence deposited in the GenBank under accession number FR748152. 31 No plasmid-mediated quinolone resistance determinants were detected by PCR.
Molecular typing revealed each of the isolates to be closely related, belonging to a single RAPD or PFGE-defined clone (Fig. 1). Replicon typing revealed IncA/C-, IncR-, and IncFIA-like replicons in all the P. stuartii isolates. E. coli DH10B transformants were positive for all resistance genes tested (blaTEM-1, blaSHV-5, blaVEB-1, bla-VIM-1, and rmtB), and only IncA/C and IncR replicons irrespective of the selective antibiotic used (cefotaxime, trfCTX, ertapenem, trfERT, or kanamycin, trfKAN). Analysis of E. coli DH10B transformed from PS70 by S1 digestion confirmed in all colonies the presence of an IncA/C and IncR-like plasmid of ∼220 kb (Fig. 2).
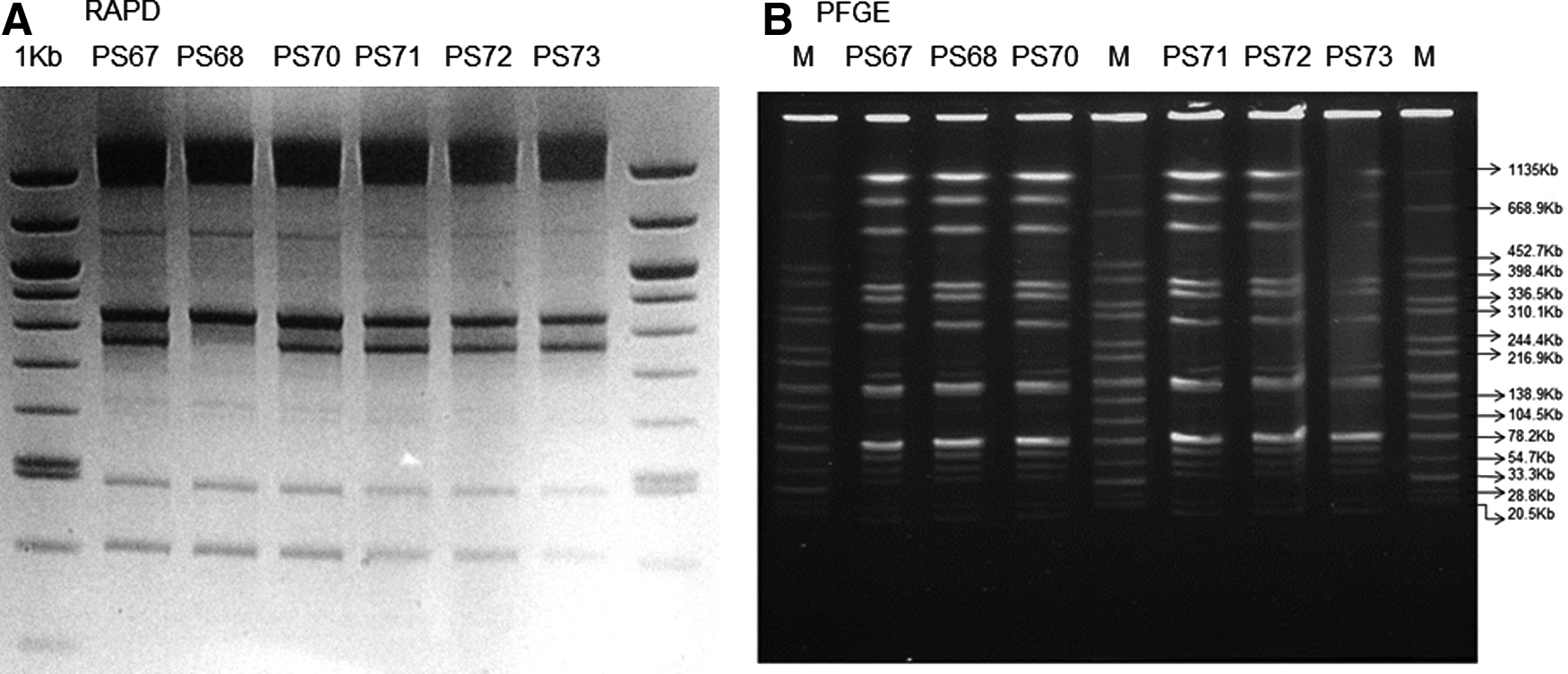
Molecular typing of Providencia stuartii outbreak strains by
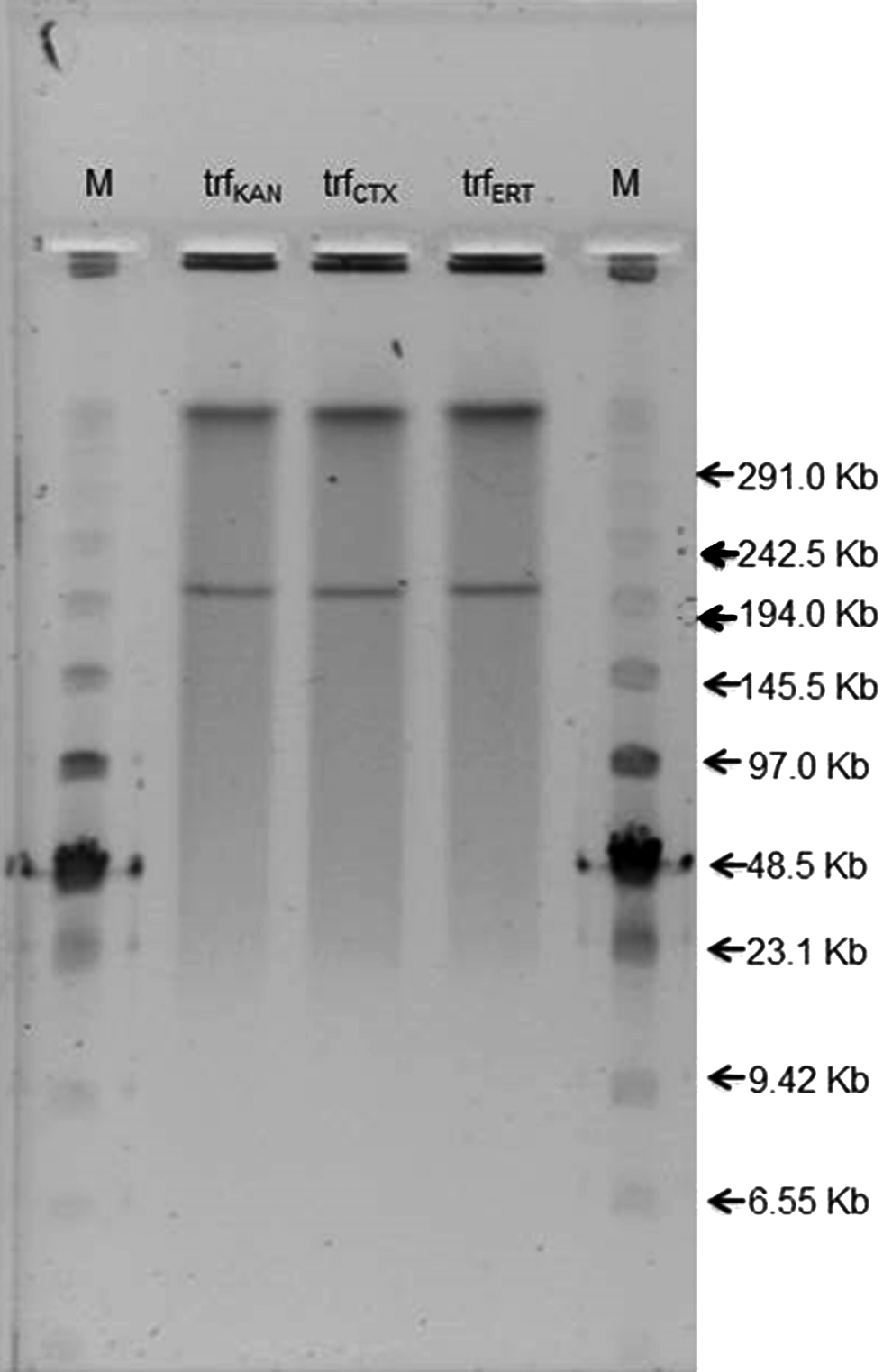
S1 digest of Escherichia coli DH10B transformants selected on kanamycin, cefotaxime, and ertapenem after electroporation with PS70 plasmidic DNA.
Analysis of transconjugants (trcERT and trcKAN) confirmed transfer of plasmids of the same replicon type and the blaTEM-1, blaSHV-5, blaVEB-1, bla-VIM-1, and rmtB genes from P. stuartii PS70 to E. coli CSH26 but at a low frequency (<10−9 trc/donor cell). Antimicrobial susceptibility testing of transconjugants confirmed resistance to aminoglycosides, ampicillin, and cephalosporins and a reduced susceptibility to carbapenems (Table 2).
Discussion
Over the past decade, MDR strains of Enterobacteriaceae have emerged as a significant threat to healthcare systems worldwide. The success of these organisms is partly down to the evolution of high-risk epidemic clones, which display heightened virulence, resistance to critical antimicrobial agents, and/or the ability to colonize individuals for long periods. These include strains, such as E. coli ST131 025b and K. pneumoniae ST258, which have spread globally and are increasingly involved in community-acquired infections. 30
The success of many of these clones is due to the acquisition and maintenance of plasmids, which may confer fitness and competitive advantages as well as antimicrobial resistance. Important in the dissemination of plasmid-mediated resistance is the ability of plasmids to replicate efficiently within the host cells. Plasmids can therefore be assigned to specific replicon (Rep) types based on their host range and their ability to coexist in the same cell. 6
Resistance to third-generation cephalosporins and carbapenems (CRO) due to the production of ESBLs and carbapenemases is particularly concerning as it promotes the wider use of aminoglycosides, polymyxins (colistin), and tigecycline as treatments, which are usually considered options of last resort. Providencia spp. are intrinsically resistant to both colistin (COL) and tigecycline (TGC), and increased usage has been identified as a factor in the growing prevalence of P. stuartii infections worldwide. 18 In this study, we identified and characterized an extensively drug-resistant clone involved in a nosocomial outbreak in a critical care unit in Greece that carried a plasmid-encoded novel complement of resistance genes.
MDR and PDR strains of P. stuartii have become a particularly important problem in North Africa and the southern Mediterranean. Reports of plasmid-mediated β-lactam resistance in P. stuartii from these regions have increased over the past decade and are summarized in Table 3. The dissemination of ESBLs in P. stuartii has been highlighted in a report from Italy. In an epidemiological study from 1999 to 2002, three MDR clones were defined, each producing either a TEM-52 or a TEM-72 β-lactamase. 43 Cefotaxime resistance mediated by blaTEM-92 carried on a 50-kb plasmid was also found in a P. stuartii isolate recovered in France. 10 Plasmids encoding class A β-lactamases (PER-1 and VEB-1) have been identified in Kosovo, Algeria, and Tunisia,3,26,38 initially not only in single isolates but also during an outbreak of blaVEB-1-producing P. stuartii in a military hospital. 26 These genes were localized to medium-sized plasmids ranging in size from 50 to 100 kb, although information on their replicon types was not then available (Table 3).
N/R, not reported; PDR, pan-drug resistant; PFGE, pulsed-field gel electrophoresis.
P. stuartii producing class B metallo-β-lactamases were first identified in Greece. These strains, although susceptible to some carbapenems, carried blaVIM-1 in a class 1 integron structure (blaVIM-1, aacA4, dfrA1, and aadA1) maintained on IncN conjugative plasmids. 33 The blaVIM-1 carried by the strain involved in our outbreak was contained within a similar cassette (aacA7, dfrA1, and aadA1), a structure that appears to be common among carbapenemase-producing Enterobacteria in Greece. 31 Spread of blaVIM-1 IncN-like plasmids into E. coli and K. pneumoniae has subsequently occurred throughout Greece and Italy. 16
MDR P. stuartii carrying plasmids with IncA/C-like replicons is a more recent development. These are often >100 kb in size and have been found to contain genes also encoding NDM-1 and class D carbapenemases (OXA-48), as well as AmpC-like cephalosporinases (CMY and DHA) and plasmid-mediated quinolone resistance determinants (QnrA and AAC-6′-Ib-cr). Almost all these strains have been recovered in or have links with travel to North Africa (Tunisia and Algeria) or the southern Mediterranean (Table 3). 29 Plasmids with IncA/C-like origins of replication have a broad host range, are able to replicate in most species of Enterobacteriaceae, and can be transferred in vitro and in vivo by conjugation.
The plasmids extracted from the outbreak strains we studied here were large (>200 kb). They contained genes that were able to transfer cephalosporins (blaSHV-5 and blaVEB-1), carbapenem (blaVIM-1), and aminoglycoside (aacA7, aadA1, and rmtB) resistance to susceptible strains of E. coli and also displayed a multi-replicon structure consisting of both IncA/C and IncR elements. A 180-kb multi-replicon IncA/C, IncR plasmid, pTC2, was recently sequenced from another MDR P. stuartii isolate recovered in Greece. This contained both blaSHV-5 and the same blaVIM-1 class 1 cassette as PS70 but not blaVEB-1 or the rmtB methyltransferase. 14
Interestingly, this mosaic plasmid is thought to have evolved in vivo, being recovered from a patient following antimicrobial therapy and also in transconjugants selected in the gut of gnotobiotic mice. 13 Therefore, the 220-kb plasmid we have described likely represents a further evolution of the pTC2 backbone, incorporating now blaVEB-1 and rmtB into the structure. Multi-replicon pTC2-like plasmids could therefore become efficient vehicles for a wider dissemination of carbapenem and MDR resistance among pathogenic Enterobacteriaceae.
Strains carrying these plasmids will be increasingly difficult to treat. The six P. stuartii we analyzed retained some susceptibility to meropenem and doripenem. Although meropenem by prolonged infusion has been used to successfully treat carbapenemase-producing Enterobacteriaceae with MIC ≤4 mg/L, 27 all three of the patients with P. stuartii infections who were treated with meropenem in this outbreak died. Combination therapies are often considered as a treatment option for MDR and PDR Gram-negative infections. Most of these rely on synergy observed between antimicrobials in vitro, most frequently involving colistin. Although success has been seen in some cases, the high level of intrinsic resistance to colistin and other cationic peptides in Providencia makes this unlikely to be a viable solution for this organism or other members of the Proteeae.
The increasing number of reports of MDR strains of Providencia in the Mediterranean region, their role in outbreaks, evolution, and transmission of MDR plasmids is a cause for concern. Enhanced surveillance and identification of the factors contributing to the success of these strains are now urgently required to limit their spread and impact on human health.
Footnotes
Disclosure Statement
L.M.P., J.B., and D.W.W. were supported by internal funds from Queen Mary University London. This work was generated as part of routine work. L.M.P., J.B., and D.W.W. have received research funding from Pfizer/Wyeth, Basilea, and Astellas in the preceding 5 years for unrelated projects, which do not represent any conflict of interest.