
Abstract
To emphasize the importance of the appropriate use of antibiotics in aquaculture systems, the prevalence of resistance to 25 antimicrobials was investigated in 42 Aeromonas veronii strains isolated from farm-raised channel catfish in China in 2006–2012. All experiments were based on minimal inhibitory concentrations (MICs), and susceptibility was assessed according to the Clinical and Laboratory Standards Institute. Some isolates displayed antibiotic resistance to the latest-generation fluoroquinolones (i.e., ciprofloxacin, levofloxacin, and norfloxacin) in vitro. Therefore, we screened for genes conferring resistance to fluoroquinolones and performed conjugation experiments to establish the resistance mechanisms. The antibiotic resistance rates were 14.29–21.42% to three kinds of fluoroquinolones: ciprofloxacin, levofloxacin, and norfloxacin. Among the 42 strains isolated, 15 carried the qnrS2 gene. The MICs of the fluoroquinolones in transconjugants with qnrS2 were more than fourfold higher compared with the recipient. Among the fluoroquinolone-resistant A. veronii strains, eight had point mutations in both gyrA codon 83 (Ser83→Ile83) and parC codon 87 (Ser87→Ile87). However, five isolates with point mutations in parC codon 52 remained susceptible to the three fluoroquinolones. In conclusion, the mechanisms of fluoroquinolone resistance in A. veronii isolates may be related to mutations in gyrA codon 83 and parC codon 87 and the presence of the qnrS2 gene.
Introduction
A
Quinolones are synthetic antibiotics that are highly active against Aeromonas species.10,11 Aquaculturists frequently use quinolones or fluoroquinolones as first-line antibiotics to prevent or cure Aeromonas-induced infections in fish.12,13 However, the widespread and unreasonable use of these drugs increases the levels of quinolone-resistant Aeromonas strains.13,14 A targeted gene mutation in the quinolone-resistance-determining region (QRDR) is reported to be the main mechanism of quinolone resistance in Aeromonas.15,16 To achieve their antibacterial effects, quinolones combine with their target DNA enzyme complex (i.e., DNA gyrase and topoisomerase IV) to block bacterial DNA replication. Changes in the target enzymes directly affect the binding capacity of the quinolones. 17 Therefore, we detected mutations in the QRDR of the gyrA and parC genes of the A. veronii isolates with PCR amplification and sequence analysis to better understand the relevant gene mutations and provide a basis for further discussion of the quinolone-resistance mechanisms of A. veronii.
Recent studies have found that quinolone-resistance mechanisms are also mediated by various plasmids (plasmid-mediated quinolone resistance [PMQR]) and their associated genes, including qnr (qnrA, qnrB, qnrS, qnrC, qnrD, and qnrVC), aac(6′)-Ib-cr, qepA, and oqxAB. 18 The Qnr-type pentapeptide proteins (QnrA, QnrB, and QnrS) prevent DNA gyrase from binding to quinolones, 19 and the presence of PMQR can facilitate the emergence of high-level fluoroquinolone resistance through mutations in the DNA gyrase and topoisomerase IV genes.20–22 PMQR genes are usually located on a plasmid, which not only enhances the adaptability of bacteria to drugs but also facilitates the spread of resistance genes among bacteria of different species. Clinical plasmid-mediated quinolone-resistant strains are currently emerging in aquaculture.21,23 Therefore, research into PMQRs of aquatic animal origin may be important in suppressing the spread of quinolone resistance.
At present, little information is available on the prevalence of fluoroquinolone-resistant A. veronii in the channel catfish. Therefore, to prevent the potential spread of resistant pathogens among fish and humans and to improve the therapeutic treatment of fish, we investigated the prevalence and mechanism of fluoroquinolone resistance in this bacterium using 42 A. veronii strains isolated from diseased channel catfish.
Materials and Methods
Bacterial strains
All the clinical strains were isolated from diseased channel catfish between 2006 and 2012 in fish farms located in Sichuan Province, China, by the Laboratory of Fish Disease Research Center. They were identified to the species level with biochemical tests, as previously described, together with partial 16S rDNA gene sequences. 24 Escherichia coli strain DH5α was provided by the Fish Disease Research Center of Sichuan Agricultural University. E. coli J53 (resistant to azide), used as the recipient in conjugation experiments, was a gift from Associate Professor Lin of the Department of Pharmacy, Sichuan Agricultural University. The reference strains were purchased from the American Type Culture Collection (ATCC): A. veronii ATCC51106 and E. coli ATCC25922. All A. veronii strains were routinely grown at 28°C in brain heart infusion broth (BHI; Becton Dickinson and Co., Franklin Lakes, NJ) and stored in BHI with 10% glycerol at −80°C. To assess the purity of the strains, single colonies were selected and subcultured thrice. The pure bacteria were harvested for subsequent experiments.
Minimal inhibitory concentration determination
The minimal inhibitory concentrations (MICs) of a range of antibiotics were determined with the microdilution method, according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI), 25 including cefazolin (CZO), cefoxitin (FOX), cefotaxime sodium (CTX), ceftriaxone (CRO), ceftazidime (CAZ), cefepime (FEP), penicillin (PEN), oxacillin (OXA), ampicillin/sulbactam (SAM), amoxicillin (AMX), piperacillin (PIP), aztreonam (ATM), imipenem (IMP), tetracycline (TCY), doxycycline (DOX), chloramphenicol (CHL), florfenicol (FFC), ofloxacin (OFX), levofloxacin (LVX), norfloxacin (NOR), ciprofloxacin (CIP), streptomycin (STR), gentamicin (GEN), amikacin (AMK), and tobramycin (TOB). All the antimicrobials were purchased from Hangzhou Microbial Reagent Co., Ltd, Hangzhou, China. These antibacterial agents were diluted 0.125–128 μg/ml. E. coli ATCC25922 was used as the quality control bacterial strain. The A. veronii strains were defined as susceptible (S), intermediate (I), and resistant (R) based on the interpretative criteria of the CLSI.
Molecular identification of quinolone-resistant genes
Chromosomal DNA was extracted with a TIANamp Bacteria DNA Kit (Tiangen Biotech Co., Ltd, Beijing, China) and used as the PCR template. To analyze the chromosomal quinolone-resistance determinants, the QRDRs of the gyrA and parC genes (encoding subunits of DNA gyrase and topoisomerase IV, respectively) were amplified and sequenced. Plasmid DNA was also extracted with a TIANprep Mini Plasmid Kit (Tiangen Biotech Co., Ltd) and used as the template from which to amplify the qnr genes (qnrA, qnrB, and qnrS). All the primers used for the detection of resistance genes are shown in Table 1. Primers gyrA F/R and parC F/R were designed from the nucleotide sequence of the A. veronii gyrA and parC genes, respectively (GenBank accession numbers 10487827 and 10486430, respectively). All the primers were synthesized by Sangon Biotech Co., Ltd (Shanghai, China). PCRs were performed in total volumes of 50 μl containing 1 μl of each primer, 2 μl of template DNA, 25 μl of 2× Taq Master Mix (TaKaRa Biotechnology Co., Ltd, Dalian, China), and 21 μl of ddH2O. The amplified PCR products were analyzed with electrophoresis (Bio-Rad Laboratories, Inc., Hercules, CA) on 1.0% agarose gels.
DNA sequencing and analysis
Each PCR product was ligated into the pMD19-T vector (TaKaRa Biotechnology Co., Ltd), which was used to transform E. coli strain DH5α. The recombinant plasmid was extracted with the TIANprep Mini Plasmid Kit and sequenced by Sangon Biotech Co., Ltd. The nucleotide sequences and the putative cognate amino acid sequences were aligned and compared among all the resistant and reference Aeromonas strains and published sequences (www.ncbi.nlm.nih.gov/Genbank). BLAST analyses were performed using the available Web-based tools (www.ncbi.nlm.nih.gov). The gyrA and parC nucleotide sequences of some reference strains of A. veronii were obtained from the GenBank databases (accession numbers 10487827 and 10486430, respectively). The amplified qnrS genes and known qnrS genes obtained from GenBank (accession number 221360495 for A. veronii; 334691835 for E. coli; 94959076 for Salmonella enterica; JN315884.1 for Aeromonas sobria; 157713492 for Aeromonas punctata; EU439941.1 for Aeromonas media; and 198286625, JN315883.1, and FJ233867.1 for Aeromonas hydrophila) were compared. A phylogenetic analysis was performed with Clustal X 2.0 and the Molecular Evolutionary Genetics Analysis (MEGA) 4 software. The neighbor-joining method was used to construct the phylogenetic tree.
Conjugation
Conjugation experiments were performed in Luria–Bertani (LB) broth, with E. coli J53 as the recipient. Cultures of donor and recipient cells in logarithmic phase (0.5 ml each) were added to 4 ml of fresh LB broth and incubated overnight without shaking. Transconjugants were selected on LB agar plates containing sodium azide (100 μg/ml) for counterselection. To determine whether quinolone resistance was transferred, the colonies were replica plated on LB agar with tetracycline (8 μg/ml). In a previous study, we demonstrated that the tetA and qnrS genes coexist (data not shown). The plasmid DNA isolated from the transconjugants was used as the template to amplify the qnrS genes with primers 5′-GCAAGTTCATTGAACAGGGT-3′ and 5′-TCTAAACCGTCGAGTTCGGCG-3′. 5 The MICs of the antimicrobials CIP, LVX, and NOR were measured in the donor, recipient, and transconjugant strains with microdilution, according to the guidelines of the CLSI.
Results
Resistance to antimicrobials
The susceptibility of 42 clinical isolates of A. veronii to 25 antimicrobial agents was evaluated, and the results are summarized in Table 2. The resistance rates for oxacillin and penicillin G reached 100%. The resistance rates for other β-lactams were generally 7.14–71.43%. The tetracycline resistance rate was 66.67% and that of doxycycline was 57.14%. The resistance rates to chloramphenicol and florfenicol were both 14.29%, and the A. veronii isolates were susceptible to the latest-generation cephalosporins, but the resistance rates to first-generation cephalosporins were about 50.00%.
AMK, amikacin; AMX, amoxicillin; ATM, aztreonam; CAZ, ceftazidime; CHL, chloramphenicol; CIP, ciprofloxacin; CRO, ceftriaxone; CTX, cefotaxime sodium; CZO, cefazolin; DOX, doxycycline; FEP, cefepime; FFC, florfenicol; FOX, cefoxitin; GEN, gentamicin; IMP, imipenem; LVX, levofloxacin; NOR, norfloxacin; OFX, ofloxacin; OXA, oxacillin; PEN, penicillin; PIP, piperacillin; SAM, ampicillin/sulbactam; STR, streptomycin; TCY, tetracycline; TOB, tobramycin.
The CIP and LVX MICs ranged from <0.125 to 16 μg/ml, and the NOR MICs ranged from <0.125 to ≥128 μg/ml (Table 3). Thirty-three strains were sensitive to CIP, three were intermediate, and six were resistant, with a resistance rate of 14%; 32 were sensitive to LVX, two were intermediate, and eight were resistant, with a resistance rate of 19%; 30 strains were sensitive to NOR, six were intermediate, and six were resistant, with a resistance rate of 14%. Among the quinolone-resistant isolates, the level of resistance to NOR was high, and SC008, SC010, and SC042 were highly resistant to NOR, with MICs ≥128 μg/ml.
MIC, minimal inhibitory concentration; I, intermediate; R, resistant; S, sensitive.
Analysis of the QRDRs of gyrA and parC
The nucleotide sequences of the gyrA and parC QRDRs in 12 fluoroquinolone-resistant clinical isolates (SC004, SC006, SC008, SC009, SC010, SC012, SC017, SC026, SC035, SC037, SC040, and SC042) and the A. veronii ATCC51106 reference strain were determined with PCR amplification and DNA sequencing. Fragments of the appropriate sizes (823 bp for gyrA and 669 bp for parC) were amplified. We compared the QRDR sequences with the sequences of A. veronii obtained from GenBank. In this study, the reference strain had no point mutations. Among the 12 strains from the diseased channel catfish, eight had an amino acid modification in the QRDR of gyrA at codon 83 and one in the QRDR of parC at codon 87, which produced the substitutions Ser83→Ile83 and Ser87→Ile87, respectively (Table 4). However, among the eight fluoroquinolone-resistant strains, no resistant mutant had only the gyrA mutation. Five isolates were only modified in the QRDR of parC at codon 52, but were still susceptible to fluoroquinolones.
PMQR, plasmid-mediated quinolone resistance.
Analysis of PMQR qnrS genes
No qnrA or qnrB gene was detected in any clinical isolate, whereas 15 isolates were positive for the qnrS gene. The sequences of the qnrS2 genes from the isolated strains were 100% identical to the sequence of the qnrS2 gene from the reference A. veronii strain. On the phylogenetic tree (Fig. 1), the amplified qnrS gene clustered closely with the known qnrS2 group. The distribution of the qnrS2 genes of the isolated strains is shown in Table 4. All the strains that contained the qnrS2 gene showed reduced susceptibility to fluoroquinolones. Among the 15 positive strains, some had mutations in the gyrA and parC genes and contained the qnrS2 gene, whereas the others only contained the qnrS2 gene. The fluoroquinolone MICs were usually higher for strains carrying a double gyrA–parC mutation and the qnrS2 gene than for those with only the qnrS2 gene.
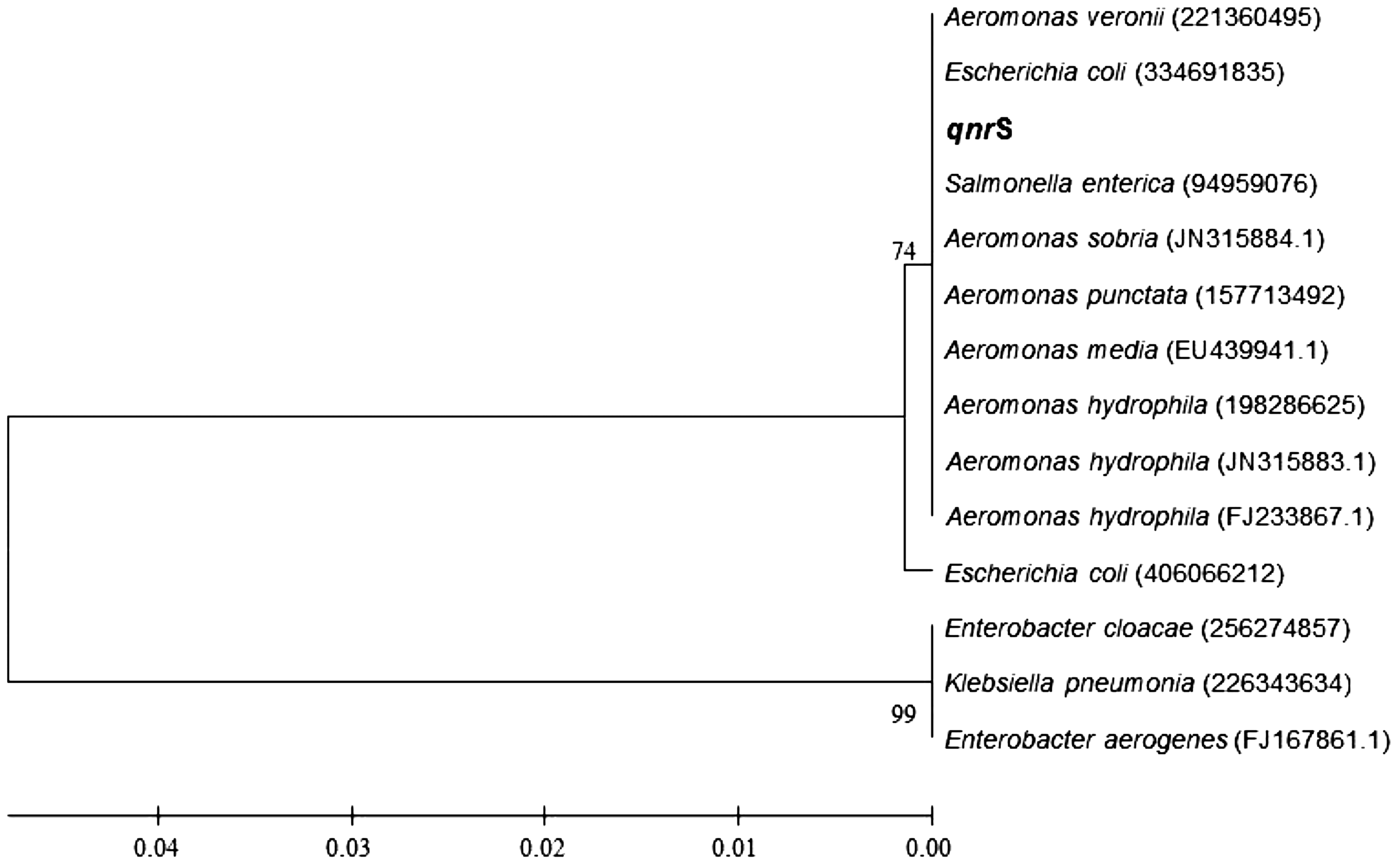
Phylogenetic tree based on qnrS2 sequences shows the relationships of selected aeromonads. The sequences were determined and compared with those available in the GenBank database using NCBI BLAST. Phylogenetic tree was constructed with the neighbor-joining method in the MEGA 4 program.
Transfer of quinolone resistance
Quinolone resistance was transferred by conjugation from two qnrS-positive donors, and the qnrS gene was detected with PCR in the plasmid DNAs isolated from the transconjugants. According to the MIC values (Table 5), the transconjugants showed reduced susceptibility to quinolones. The MICs of CIP and NOR in the transconjugants were 0.5 μg/ml, representing an increase of more than fourfold relative to the recipient, E. coli J53Az r . The MICs of quinolones were identical in the two transconjugants, suggesting the similar and stable expression of the qnrS gene.
Discussion
The diagnosis of fish diseases is often presumptive in aquaculture systems, and the first-line treatment for bacterial infection is usually antibiotics, administered without susceptibility data for the causative pathogen. However, the administration of inappropriate antibiotics is the primary cause of the increasing prevalence of antibiotic-resistant strains of bacteria, creating transferable resistance genes in fish pathogens and other bacteria in aquatic environments. The susceptibility to 25 antibiotics of 42 A. veronii strains isolated from the channel fish was assessed on the basis of the MIC breakpoint values recommended by the CLSI. We found that the latest-generation cephalosporins (i.e., CTX, CAZ, and FEP), monobactams (i.e., ATM), and aminoglycosides (i.e., AMK and TOB) had the best inhibitory activities against the A. veronii isolates tested in vitro, similar to the results described by others.26,27 In particular, most of isolates were resistant to β-lactam agents, such as penicillin and amoxicillin; so the proportion of isolates resistant to β-lactams was high, as in previously published reports. 28 The estimated antibiotic resistance rates among the A. veronii isolates were 66.67% for TCY and 57.14% for DOX. The greater prevalence of TCY resistance than fluoroquinolone resistance among Aeromonas strains is consistent with other environmental and clinical studies.29,30 Aeromonas strains are almost universally susceptible to fluoroquinolones. However, the large amounts of fluoroquinolones used in aquaculture in the last decade have been associated with an increasing trend in the prevalence of quinolone resistance. According to our results, 14.29% (6/42), 19.05% (8/42), and 14.29% (6/42) of the isolates were resistant to CIP, LVX, and NOR, respectively.
The acquisition of quinolone resistance by Aeromonas is principally associated with mutations in QRDRs, particularly in gyrA codons 83, 87, and 92 and parC codons 80 and 84.15,16,31 In this study, eight isolates showed serious resistance to fluoroquinolones. A genetic analysis based on QRDR sequences revealed that these resistant strains all carried point mutations in gyrA codon 83 and parC codon 87. All eight isolates had Ser83→Ile83 and Ser87→Ile87 substitutions in gyrA and parC, respectively. A mutation at residue 83 (Ser83→Ile83) of gyrA is most frequently encountered, together with a substitution in parC codon 80 (Ser80→Ile80) in strains of Aeromonas,32,33 but little information is available about the substitution in parC codon 87. The substitution in parC codon 87 may be another antimicrobial resistance determinant in A. veronii. The fluoroquinolone MICs detected in this study were higher than those for A. veronii strains carrying double gyrA and parC mutations, particularly at positions 83 and 80, respectively.16,33,34 Five strains only carried a single point mutation in parC codon 52 and showed no resistance to the three fluoroquinolones tested. This indicates that the mutation of residue 52 of ParC is not related to fluoroquinolone resistance and that this residue may therefore be outside the QRDR.
Apart from the point mutations in the QRDRs, the emergence of quinolone-resistant determinants on mobile genetic elements may also accelerate the dissemination of resistance among bacteria by plasmids or transposons. 35 Recently, increasing numbers of PMQR determinants have been identified in Aeromonas 20 and we screened for several qnr genes (qnrA, qnrB, and qnrS), the primary PMQR mechanism. Fifteen A. veronii strains carried the qnrS2 gene, whereas neither the qnrA nor the qnrB gene was detected in any of the 42 strains. Only four qnr determinants have been identified in Aeromonas species, qnrS2, qnrS5, qnrB1, and qnrVC.21,34,36–38 qnrS2 was the first identified and confirmed in A. veronii, in 2008, 34 and seems to be the most commonly detected qnr determinant in aeromonads isolated from diseased fish or environmental samples.16,22,39
Our results show that resistance plasmids can be transferred from A. veronii to E. coli J53, thus reducing the susceptibility of E. coli J53 to fluoroquinolones. This demonstrates that Aeromonas may play an important role in the dissemination of resistance plasmids in aquatic environment and also that this resistance plasmid can be spread among different bacterial species. As we know, the horizontal transfer of resistance plasmid is a very important factor in the generation of bacterial drug resistance. From these reservoirs, resistance genes can be disseminated by horizontal gene transfer to human pathogens, or drug-resistant pathogens from aquatic environment can reach humans directly.
Currently, several antibiotics can be used in animals in the worldwide aquatic industry, 40 but antibiotic resistance is becoming more frequent.41,42 In this study, fluoroquinolone resistance was described in terms of chromosomal mutations in the QRDRs of the gyrA and parC genes and the presence of the PMQR qnrS2 gene. Other mechanisms, such as reduced antibiotic accumulation arising from the impermeability of the bacterial membrane and/or the overexpression of efflux pump systems, may also be responsible for quinolone resistance in these A. veronii strains, but were not explored in this study. The detection of multidrug resistance in Chinese strains of A. veronii suggests that antibiotic resistance in aquaculture poses a risk to both humans and animals. Therefore, stricter guidelines for the use of fluoroquinolones are necessary to prevent the dissemination and acquisition of antibiotic resistance in aquaculture.
Footnotes
Disclosure Statement
No competing financial interests exist.