
Abstract
Methicillin resistance among staphylococci isolated from patients in northern Egypt has escalated alarmingly in the past decade. Data about the prevalence of fusidic acid (FA) resistance in Egyptian clinical isolates are limited. This work investigates the prevalence and mechanism of FA resistance among 81 methicillin-resistant staphylococcal isolates from major hospitals of Alexandria, Egypt. Some combinations for treating infections due to resistant isolates were studied. Twenty-six isolates (32.1%) were FA resistant (minimum inhibitory concentrations [MICs] = 2–1,024 μg/ml), and fusB and fusC genes coding for FA resistance were detected in 30.77% and 34.62% of the FA-resistant strains, respectively. One highly resistant isolate, S502 (MIC = 1,024 μg/ml), possessed both genes. Plasmid curing resulted in fusB loss and MIC decrease by 16–64 folds. Conjugation caused acquisition of FA resistance among susceptible isolates. Serial passages in subinhibitory FA concentrations produced mutants with increased MIC by 4–32 folds. The combination of FA with rifampin, gentamicin, or ampicillin/sulbactam, in a subinhibitory concentration, was synergistic against the isolates, including serial passage mutants, decreasing number of survivors by an average of 2–4 logs. A relatively moderate rate of FA resistance was detected in Alexandria hospitals. Combination therapy with gentamicin, rifampin, or ampicillin/sulbactam is crucial to preserve the effectiveness of FA.
Introduction
S
Methicillin resistance was first identified in staphylococci in the 1960s. 7 Nowadays, 40–70% of staphylococcal infections are caused by methicillin-resistant S. aureus (MRSA). 8 The majority of these infections are nosocomially acquired; however, community-associated MRSA infections have been reported globally and are now a reason for public health concern.9,10 Methicillin resistance among staphylococci usually signals the emergence of a difficult medical situation since these strains are usually multidrug resistant. 11 Such events raise healthcare cost and limit treatment options still available to clinicians to a restricted number of antibiotics. 12
Fusidic acid (FA) is one of these last resort agents, active in vitro against staphylococcus, including the methicillin-resistant strains.13,14 It is mainly a bacteriostatic agent, with cidal effect at higher concentrations. 14 FA locks elongation factor G (EF-G) to the ribosome, thus hindering protein synthesis. 15 Newer studies also point that the dissociation of the ribosome into subunits mediated by EF-G is also prevented by FA. 16 Resistance to FA was detected early on; however, it was not until the last two decades that the problem became clinically significant, probably due to FA abundant use and monotherapy application. 17 It is therefore recommended to combine FA with a second antistaphylococcal agent in clinical practice to minimize the risk of resistance development. 14
Several mechanisms are now recognized for FA resistance in staphylococci. The most prevalent is the horizontal acquisition of a plasmid-mediated fusB resistance determinant (a 21.9 kb plasmid pUB101) that also confers resistance to cadmium and β-lactamase.18–20 This family of protective proteins leads to the dissociation of the EF-G ribosome complex locked by the presence of FA. This allows for the release of EF-G from the ribosome, necessitating higher FA concentrations to bring about its inhibitory effect, that is, the cells become FA resistant.15,21 The same gene can also be located on a chromosome, 22 as a result of the integration of a phage-related chromosomal island.23,24
More recently, fusC and fusD, two fusB homologues have been detected in clinical staphylococcal isolates, 19 with fusC conferring resistance to S. aureus and CoNS and fusD being an intrinsic FA resistance factor among Staphylococcus saprophyticus.19,25 The fusC gene has been reported in association with three SCC (staphylococcal cassette chromosome) elements, three of which in MRSA isolates. These are SCCmecN1 (carrying mecA), pseudo-SCCmec-SCC-SCC CRISPR (carrying a clustered regularly interspaced short palindromic repeat [CRISPR] region and mecA), and SCCfusC.26–28
Another important mechanism of resistance is the mutations in fusA gene, leading to alteration in EF-G structure and reduced affinity to the drug. 29 Lannergard et al. reported frequent mutations in fusA in the presence of fusB and fusC among S. aureus isolates obtained from Danish hospitals. 30
Only a few studies dating back in the early 1980s and 1990s assessed the clinical effectiveness of FA in treating superficial bacterial skin infections 31 and conjunctivitis 32 in Egypt. However, very little information is available about the clinical usefulness of FA in treating infections in Egypt in the recent years. Also, scarce are the studies that describe resistance levels to FA among Egyptian staphylococcal isolates. In this work, we are reporting on the levels of FA resistance among staphylococcal clinical isolates obtained from patients in Alexandria, Egypt. We are also studying the mechanisms of resistance development and suggesting some useful combinations to treat infections caused by FA-resistant staphylococci.
Materials and Methods
Test organisms
Collection of methicillin-resistant staphylococcal isolates occurred over a period of 5 months (May through September 2011). A subset of these isolates (n = 81), for which the complete demographic data were available, was included in the current study. They were obtained from clinical microbiology laboratories affiliated with El-Meery tertiary teaching hospital (n = 39), Medical Research Institute Hospital (n = 22), and El Mowasah Hospital (n = 20). The clinical specimens were obtained from pus (n = 36), blood (n = 21), sputum (n = 11), urine (n = 9), and bronchoalveolar lavage (n = 4). The isolates were identified according to the conventional methods based on their morphological, tinctorial, cultural, and biochemical characteristics, including growth on mannitol salt agar and production of catalase and coagulase, with coagulase production being the determining factor distinguishing S. aureus from CoNS.33,34
Methicillin (β-lactam) resistance was determined based on the susceptibility of the isolates to oxacillin and cefoxitin using the disk diffusion method following the Clinical and Laboratory Standards Institute (CLSI) guidelines for susceptibility checking. 35 Frozen glycerol stocks (30%) of the identified isolates were stored at −70°C.
Antibiotics
FA was kindly supplied by Pharaonia Company. Gentamicin (CN), rifampin (RIF), and ampicillin/sulbactam (SAM) were purchased from the Egyptian pharmaceutical market.
Antimicrobial susceptibility tests
The minimum inhibitory concentration (MIC) of FA was determined using the agar dilution method according to the CLSI guidelines. 35 The MIC was defined as the least concentration causing complete inhibition of the organism or no more than five discrete colonies per plate, under the test conditions. Susceptibility was interpreted using the European Committee for Antimicrobial Susceptibility Testing (EUCAST) 2015/British Society of Antimicrobial Chemotherapy (BSAC) 2012 criteria (susceptible, MIC ≤1 μg/ml; resistant, MIC >1 μg/ml). The testing MIC range of FA was 0.25–1,024 μg/ml.
Susceptibility to CN, RIF, SAM, tetracycline (TE), ciprofloxacin (CIP), and sulfamethoxazole/trimethoprim (SXT) was carried out using the disk diffusion method, and the agar dilution technique was used to determine MIC of select antibiotics, following the CLSI guidelines. 35 S. aureus ATCC 6538 was used as quality control organism.
Study of genetic basis of resistance to FA
Detection of FA-acquired resistance determinants by PCR
The presence of acquired FA resistance determinants (fusB and fusC) was detected by colony PCR using the following primers: B-F: 5′-CCGTCAAAGTTATTCAATCG and B-R: 5′-ACAATGAATGCTATCTCGACA, C-F: 5′-GGACTTTATTACATCGATTGAC and C-R: 5′-CTGTCATAACAAATGTAATCTCC. 36 The cells were lysed at 95°C for 10 min, and then, PCR was conducted at the following cycling conditions: an initial denaturation at 95°C for 30 sec, followed by 34 cycles of denaturation (95°C for 15 sec), annealing (50°C for 15 sec) and extension (72°C for 20 sec), and a final extension step at 72°C for 10 min. The expected amplicons were 492 bp for fusB and 411 bp for fusC. 36 PCR products were separated by electrophoresis in 1% agarose gel.
Plasmid curing
Three clinical isolates with fusB as detected by PCR were subjected to elevated temperature (43°C)-mediated plasmid elimination by sequential passages in Luria-Bertani (LB) broth (∼100 cells into 100 ml), with shaking for about 30 generations. Cured strains were diluted and plated on LA plates (LB plus 1% agar) to obtain single colonies. Loss of ampicillin resistance was screened by replica plating at 37°C.30,37 Loss of the plasmid was confirmed by decreased FA MIC values and absence of fusB band following PCR.
Conjugation
A conjugation experiment was carried out between two FA-resistant isolates that carried the fusB gene and two FA-susceptible isolates. In one case, nutrient broth was simultaneously inoculated with ∼1 × 107 CFU of S412 (FA-susceptible S. aureus) and 1 × 107 CFU of S225 (fusB positive, CoNS). In the other case, nutrient broth was simultaneously inoculated with ∼1 × 107 CFU of S313 (FA-susceptible, RIF-resistant S. aureus) and 1 × 107 CFU of S410 (fusB-positive, RIF-susceptible S. aureus). Following overnight incubation at 37°C, the resulting cultures were 10-fold serially diluted. Two hundred microliters of the previously diluted cultures was spread on mannitol salt agar plates for the S412/S225 set and on nutrient agar plates for the S313/S410 set. The plates were incubated at 37°C for 24 hr. The dilutions that resulted in 50–200 colonies/plate, following incubation, were used to calculate the count.
Gain of FA resistance was checked for the yellow colonies in the first set, and in the second set, combined resistance to FA and RIF was checked by replica plating.
Frequency of selection of resistant mutants
Frequencies of a single-step mutation to resistance were determined by plating FA-susceptible isolates onto nutrient agar plates containing 1 μg/ml of FA, corresponding to 1, 2, or 4× MIC. The experiment was done in triplicate, with plating of a maximum of 1 × 1011 CFU. Mutation frequencies were calculated as the ratio of the number of resistant colonies at 24 hr to the number of CFU inoculated. 38
Laboratory induction of FA resistance among susceptible isolates
Induction of FA resistance among susceptible isolates was carried out in the laboratory by serial passages of the isolates in nutrient broth containing ½× MIC of FA. MIC was determined by agar dilution after 5 and 10 passages. The stability of the acquired resistance was checked by passing the isolates in nutrient broth free of FA before MIC determination.
Combined biocidal activity of FA and CN, RIF, or SAM against selected resistant strains
In vitro bactericidal activity of FA in combination with CN, RIF, or SAM was tested against three FA-resistant isolates and three resistant mutants using time–kill assays. Probe tubes contained FA/CN, FA/RIF, or FA/SAM at concentrations corresponding to ½× MIC for each strain. The starting bacterial density was ∼1 × 106 CFU/ml. Tubes were incubated aerobically with shaking in a shaking incubator (140 rpm) at 35°C for 24 hr. Aliquots were removed from each tube, and serial dilutions were plated in duplicates onto Mueller-Hinton plates after 0, 3, 6, and 24 hr of incubation. Colony counts were performed after 24 hr of incubation at 35°C. Controls lacking one or both antibiotics were performed alongside the experiment.
Results
Antimicrobial susceptibility tests
The in vitro antimicrobial activity of FA against the studied isolates is presented in Fig. 1. Of the 81 methicillin-resistant staphylococci included in the study, 65 (80.25%) were MRSA and 16 (19.75%) were CoNS. Resistance to FA was detected in 26 isolates, of which 16 (61.5%) were MRSA and 10 (38.5%) were methicillin-resistant CoNS, making FA resistance prevalent in 9.23% of MRSA and 62.5% of CoNS. The resistant isolates had MICs that ranged between 2 and 1,024 μg/ml, with an MIC50 of 0.25 μg/ml and MIC90 of 16 μg/ml. The MIC values of FA against the 26 resistant isolates, in addition to the susceptibility of these isolates to CN, RIF, and SAM, are shown in Table 1.
BAL, bronchoalveolar lavage; CN, gentamicin; FA, fusidic acid; MIC, minimum inhibitory concentration; MRI, Medical Research Institute Hospital; RIF, rifampin; SAM, ampicillin/sulbactam; R, resistant; S, susceptible.

FA MIC distribution against the clinical isolates determined by the agar dilution technique. FA, fusidic acid; MIC, minimum inhibitory concentration.
Fourteen of the 26 (53.9%) were also resistant to CN, 5 (19.2%) were resistant to RIF, and 17 (65.4%) were resistant to SAM. In addition, 15 (57.7%) of the 26 isolates were resistant to TE, 8 (30.8%) were resistant to CIP, and 4 (15.4%) were resistant to SXT (data not shown). Three isolates, S205, S328, and S502, were resistant to all four antibiotics (FA, CN, RIF, and SAM as shown in Table 1), and S328 and S502 were resistant to all tested agents except for SXT (data not shown). The three isolates were collected from different clinical specimens, from different hospitals, and had somewhat different antibiotic resistance patterns.
fusB-based and fusC-based resistance to FA
The prevalence of fusB and fusC among the 26 FA-resistant isolates was checked by PCR (Table 1). The genes fusB and fusC were detected in eight (30.8%) and nine (34.6%) of the isolates, respectively. Five (62.5%) out of the eight carrying fusB were S. aureus, whereas eight (88.9%) of the nine isolates carrying fusC were S. aureus. Only one isolate, S502, carried both fusB and fusC and had an FA MIC of 1,024 μg/ml.
Six (75%) of the isolates carrying fusB had MIC in the range of 4–16 μg/ml, and 2 (25%) of the isolates needed a higher FA concentration (1,024 μg/ml) for inhibition. The presence of fusC alone in the isolates was always associated with an FA MIC of 16 μg/ml. A curing experiment done on a subset of 3 of the isolates carrying fusB, S225, S410, and S501, resulted in the absence of the gene in the cured strains (Fig. 2 and the table). The table shows the dramatic decrease in S225, S410, and S501 MIC values by 32, 16, and 64 folds, respectively, after curing. Conjugation between the FA-resistant, fusB-positive S225 and S410 on the one hand and FA-susceptible S412 and S313 on the other hand resulted in 25% and 17% of the populations becoming FA resistant, respectively.

Gel of a PCR amplification of fusB before and after curing of the resistance plasmid in isolates 225, 410, and 501. Table indicates FA MIC before and after curing of resistance plasmid in isolates 225, 410, and 501.
Selection of resistant mutants
Exposure of 10 FA-susceptible isolates (9 S. aureus and 1 CoNS, S208) to 1 μg/ml of FA (EUCAST- and BSAC-susceptible breakpoint concentration), corresponding to 1, 2, or 4× MIC resulted in the selection of naturally occurring subpopulations resistant to FA at the rate of 10−6.3–10−8 (Table 2). In another experiment, the same set of isolates was serially passed in ½× MIC of FA for 10 times to induce the emergence of resistant mutants. The resulting strains showed an elevation in MIC in the range of 4–32-fold at the end of the experiment, with 50% of the strains showing an MIC increase by 8 fold (Table 2).
Time–kill studies
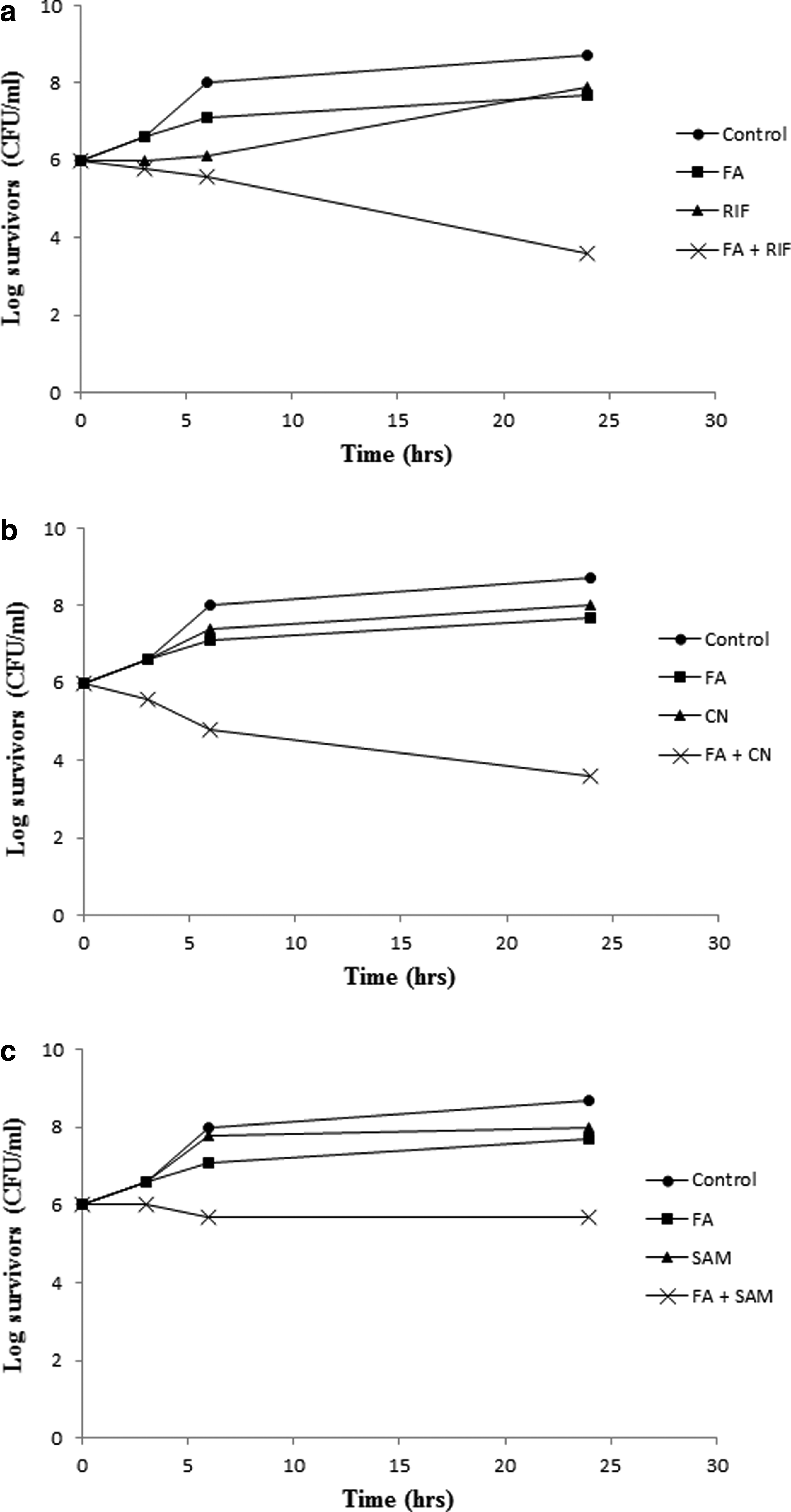
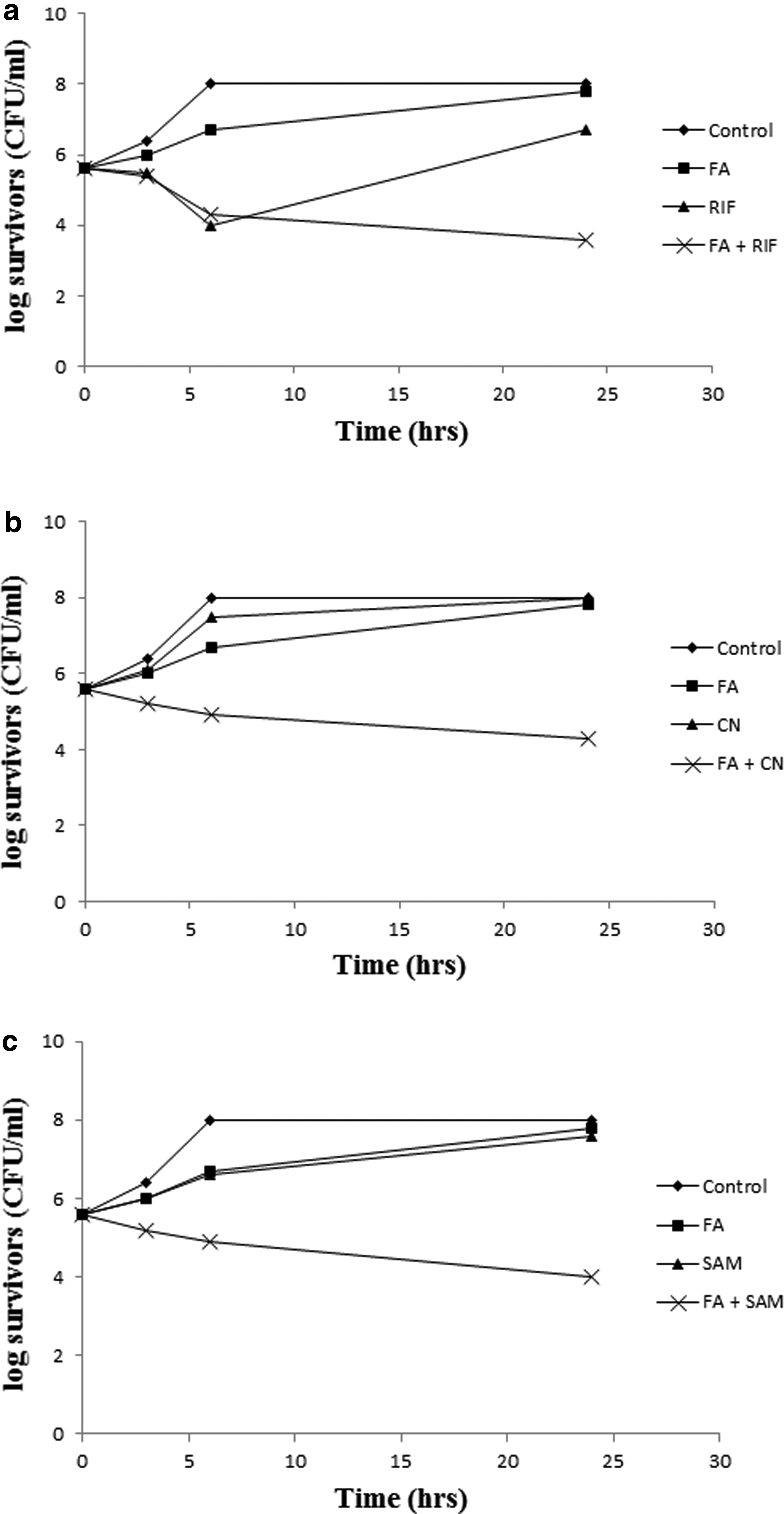
The in vitro bactericidal activity of FA combinations with CN, RIF, or SAM in ½× MICs was tested against three resistant isolates (S414, S501, and S502) and three resistant mutants (ind. S208, ind. S326, and ind. S505). Isolate S501 (MIC of FA, CN, RIF, SAM = >1,000, 100, >100, 100 μg/ml, respectively) represents the response of the tested resistant isolates to the combinations (Fig. 3), whereas the resistant mutant ind. S326 (MIC of FA after induction, CN, RIF, SAM = 1.6, 50, 0.4, 50 μg/ml, respectively) represents the response of the strains that acquired FA resistance following laboratory induction (Fig. 4).
Time–kill curves showing bactericidal activity of
Time–kill curves showing bactericidal activity of
Figure 3 demonstrates the synergistic effect of the combinations relative to either antibiotic alone, as demonstrated in a minimum of 4-log reduction in the count of surviving cells with FA/CN and FA/RIF and 2-log reduction with FA/SAM. Figure 4 shows a similar pattern against ind. S326. The beneficial effect of the combinations was also seen against FA-susceptible isolates (data not shown).
Discussion
Methicillin resistance in staphylococci and their simultaneous multidrug resistance is a real concern for clinicians worldwide. 39 Tens of thousands are afflicted by MRSA caused infections in Europe and United States annually costing the healthcare systems billions of dollars in extended hospital stays as well as increased patient morbidity and mortality.40,41 In Egypt, little is reported in the literature about the actual prevalence of methicillin resistance in staphylococcal isolates or the subsequent healthcare burden. Yet, our data point to a resistance rate of 38–60% among S. aureus in the past 2 years (unpublished data).
The multidrug-resistant status in methicillin-resistant isolates necessitates the use of nonconventional and alternative treatment approaches. 12 FA is one drug that has been recommended to fulfill this role. 14 FA has been in clinical use for more than four decades for the topical treatment of superficial skin infections. 42 A study from the mid-1990s reported a mean resistance rate to FA of 5% worldwide. 43 Over the following decade, the rate escalated to reach 92% in Kuwait in 2004, 1.4–3.1% in Italy, Poland, Spain, and Sweden, 62.4% in Greece, 19.9% in Ireland, 44 13–14.2% in Morocco in 2008–2009,45,46 and 7% in Canada and Australia. 47 Liu et al. reported a low rate of FA resistance in China between 2008 and 2009. 48 In the United States, the resistance rate is very low (0.3%), probably because the drug has only been recently proposed for clinical use there.47,49
Although the frequency of resistance remains fairly low, it is a problem that could limit the usefulness of FA in treating staphylococcal infections since the selective pressure due to the antibiotic use might prompt further increase in resistance rates.14,48 This is especially significant since Dobie and Gray reported FA resistance in isolates from children in Birmingham Children's Hospital, half of which with no history of FA treatment. 17
FA is available in the Egyptian market in the form of creams for the monotherapy treatment of skin infections. A lone study from the 1990s described the effectiveness of FA in the treatment of superficial bacterial skin infections in Egypt. 31 However, a literature survey revealed no studies in the last three decades that assessed the prevalence of FA resistance among Egyptian clinical isolates, neither are there any recent studies that re-evaluated the clinical effectiveness of this drug.
In this work, we are reporting the rate of FA resistance among 81 methicillin-resistant staphylococcal isolates collected from Alexandria hospitals. We are also studying the predominant mechanism of FA resistance in the resistant isolates and mutants, as well as suggesting useful combinations for the treatment of these strains.
FA was found to be active against 67.9% of the tested isolates, second to SXT and RIF that were active against 86.4% and 82.7% of the isolates, respectively. FA action was, however, superior to TE (37%), CN (35.8%), and SAM (23.5%). Howden et al. 13 reported a FA resistance rate of 8% among S. aureus in Australia, which is lower than our rate of 32.1%. Seven of our isolates resistant to FA (S216, S217, S218, S224, S225, S317, and S337) were found to be susceptible to CN, RIF, and SAM (Table 1). All of these isolates, but S218 and S224, were CoNS. It is to be noted that FA MIC ranged between 1.6 and 1,000 μg/ml among resistant S. aureus, 3.1 and 250 μg/ml among resistant CoNS. Another study by Chen et al. described a FA-resistant S. aureus population that was resistant to nearly all tested antibiotics, except for vancomycin, teicoplanin, linezolid, nitrofurantoin, quinupristin–dalfopristin, chloramphenicol, and RIF. 50
Two main mechanisms have been described to explain FA resistance in S. aureus. The predominant mechanism involves the protection of the drug target site by fusB-type resistance proteins. The second known mechanism of FA resistance in S. aureus is due to mutations in fusA, the gene encoding EF-G, thereby making the target site less susceptible to the action of the antibiotic.29,51
In the present work, fusB and fusC were detected in 30.8% and 34.6% of the studied isolates, respectively, with a single isolate S502 carrying both genes. Three of the isolates carrying fusB were CoNS, which highlights the potential of CoNS to serve as reservoir for resistance gene transfer to pathogenic S. aureus. Castanheira et al. reported a prevalence rate of fusB and fusC among European S. aureus isolates resistant to FA of 10.1% and 16.9%, respectively. 44 Huang et al. reported a fusB prevalence rate of 100% among the S. aureus Chinese isolates resistant to FA, none of which had fusC. 52 Another study also conducted on Chinese isolates revealed a prevalence rate for fusB and fusC of 50% each, with fusB detected in methicillin-susceptible S. aureus (MSSA) and fusC detected in MRSA. 48 On the contrary, the prevalence of fusC and fusB among 34 Taiwanese MRSA collected between 2008 and 2009 was 74% and 4%, respectively, where fusB has been so far only detected in MSSA. 50 A study of Moroccan isolates described similar findings with fusC, the most common resistance determinant present in 11.43% of the isolates. 46
The detection of fusB in our isolates corresponded with FA MICs that ranged between 4 and 1,024 (Table 1). O'Neill et al. associated the lower MIC value (4 μg/ml) with fusB presence on the chromosome and the higher value of 16 μg/ml with the gene being carried on plasmid. The reason behind the high FA MIC when fusB is located on a plasmid is probably due to the higher number of copies of the genes (11–14) relative to the single copy, when fusB is expressed from a chromosome. 22
A plasmid-curing experiment conducted on isolates S225, S410, and S501, whose MICs were 16, 16, and 1,024 μg/ml, respectively, resulted in MIC reduction by 32-, 16-, and 64-fold, respectively (Fig. 2 and table). These findings show that fusB was plasmid mediated and point fusB as a major mechanism of FA resistance in these isolates, which agrees with the previous studies mentioned above. In addition, a conjugation experiment between S225 or S410 and either of two susceptible isolates resulted in these isolates acquiring FA resistance at the rate of 17–25%. However, in the case of S501, plasmid curing dramatically reduced the resistance level but did not make the isolates susceptible to FA, with an MIC of 16 μg/ml, well above the resistance cutoff.
The above finding suggests the presence of another FA resistance mechanism other than fusC that was absent in that isolate, probably that mechanism would involve mutations in fusA. Chen et al. reported no change in FA MIC value after plasmid curing, probably due to the presence of fusC, which encodes a protein that acts in a way similar to fusB, without indication to whether they act synergistically or additively. 50 In the present study, the single isolate that displayed both genes, S502, showed a high FA MIC (1,024 μg/ml); however, the same value was recorded with S501 that only had fusB. This supports the reasoning of Chen et al. that fusB and fusC do not act in combination. On the contrary, all isolates displaying fusC, except for S502 carrying fusB as well, had an FA MIC of 16 μg/ml. The 16 μg/ml MIC value falls within the MIC range of 2–64 μg/ml previously described. 50
Staphylococcus subpopulations resistant to FA emerged at a rate of 10−6.3–10−8, on exposure to 1 μg/ml of FA, which corresponded to 1, 2, or 4× MIC. Moreover, exposing these susceptible isolates to ½× MIC of FA for 10 passages resulted in MIC increase by 2–32-fold (Table 2). This is consistent with the findings of O'Neill et al. that reported a mutation rate of 10−7–10−8 on exposure of MRSA and MSSA to 10 μg/ml of FA but not 15 or 30 μg/ml. 53 However, Munckhof et al. 54 suggested that FA resistance still developed when MRSA strains were exposed to 64× MIC of FA alone. Yet, Jensen and Kler 55 isolated S. aureus mutants resistant to FA due to mutations in fusA without any exposure to FA.
Although the rate of emergence of FA resistance among Egyptian isolates is not very high, it is reason for concern since FA was considered one of the last-resort agents still effective in treating resistant MRSA infections. This led us to devise some combinations of FA with other antibiotics in a trial to retard resistance development and save the usefulness of a valuable antimicrobial. The isolates included in the experiment were three FA-resistant MRSA, in addition to two MRSA, and one methicillin-resistant CoNS that acquired FA resistance upon laboratory induction. One methicillin-resistant CoNS and two MRSA susceptible to FA were tested alongside the resistant isolates. The agents included in the current combinations were CN, RIF, and SAM.
Combinations of FA with RIF and CN were also reported elsewhere.56,57 The SAM/FA combination was included in the current study as a nonconventional FA combination and a way to revive the SAM mixture that is otherwise inactive against methicillin-resistant staphylococcal infections. With the exception of RIF that showed higher inhibitory potential on its own, >50% of the isolates were resistant to CN and SAM. Combining either of these agents with FA in subinhibitory concentrations resulted in a synergistic effect, reducing the count of survivors by at least 4 logs with RIF and CN and 2 logs with SAM, relative to either antibiotic alone (Fig. 3).
The same behavior was also noted when higher concentrations of the antibiotics were used. The synergistic effect was also observed against the laboratory-induced mutants, including the CoNS isolate (Fig. 4) and the susceptible isolates, suggesting these combinations as a measure to safeguard the antimicrobial effect of FA.
Others also tested FA combinations, and Tang et al. found the combination of FA with RIF to be synergistic against MRSA in biofilm. 57 In addition, adding RIF or the β-lactams seemed to lower FA resistance development. 14 Another group also reported that a combination of CN with FA was better than CN alone in inhibiting growth of S. aureus and CoNS in biofilm mode. 56 Other combinations included FA with fosfomycin that was synergistic in 87.76% of the cases tested by Yu et al., 58 also, minocycline combination with FA showed promising bactericidal effect against biofilm-embedded MRSA. 59 The findings of Grohs et al. showed the FA/linezolid combination to lack synergism against MRSA and MSSA, yet it prevented the selection of resistant mutants. 60 However, Østergaard et al. reported antagonism between FA and methicillin in an experimental meningitis model against S. aureus. 61
This study is the first investigation of the role of fusB and fusC in FA resistance among Egyptian MRSA- and methicillin-resistant CoNS isolates. We are reporting a moderate rate of FA resistance in these isolates. Yet, we are showing that exposure of the isolates to FA monotherapy, which sometimes occurs in Egypt, can trigger the development of resistance. We are suggesting a number of FA combinations with CN, RIF, or SAM to increase FA effectiveness; however, clinical studies are still needed to confirm the therapeutic potential of the proposed combinations.
Footnotes
Acknowledgment
This work was conducted on the premises of and supported by the Department of Microbiology, Faculty of Pharmacy, Alexandria University.
Disclosure Statement
No competing financial interests exist.