
Abstract
Epidemiological studies have demonstrated that the human immunodeficiency virus (HIV)-1 drug-resistant rate among injecting drug users is higher than that in other HIV-1-positive populations, which is generally believed to be largely due to clinical nonadherence. Little is known, however, about whether heroin abuse has a direct impact on the generation of HIV-1 drug-resistant mutations. In this study, we investigated the impacts of morphine, the active metabolite of heroin, on HIV-1 infection/replication and HIV-1 drug-resistant mutations through an in vitro HIV-1-CD4+ T cell system under selective pressure from two typical antiviral drugs, Lamivudine and Nevirapine. We found that morphine treatment of MT4 cells (a CD4+ T cell line) significantly increased HIV-1 III B (a T-tropic viral strain) infection and replication in MT4 cells, and the effect of morphine on HIV-1 was mediated through an opioid receptor. More importantly, our results showed that morphine treatment not only induced more drug-resistant mutations under selective pressure from antiretroviral drugs but also shortened the mutations' generation time, compared with the control groups that were treated with antiretroviral drugs alone. Although the in vivo relevance remains to be determined, these findings provide direct in vitro evidence to support the possibility that heroin abuse itself can act as an independent factor contributing to the generation of HIV-1 drug resistance during clinical antiretroviral therapy. Therapeutic guidelines should consider this issue for heroin users with HIV infection.
Introduction
H
Despite the success of antiviral therapy, drug resistance caused by HAART has become a major factor contributing to antiretroviral therapy failure and an obstacle to HIV prevention and control, and is thus an ongoing concern worldwide. A report that summarized 17 studies of acquired HIV-1 drug resistance in Southeast Asia found that the resistance rates of NRTI and NNRTI ranged from 52 to 92% and 43% to 100%, respectively, among those with virological failure. 3 The prevalence of transmitted HIV-1 drug resistance for any class of antiretroviral drug has been estimated at around 8–20% of the untreated population.4–10 In China, a recent study found that the HIV-1 drug resistance rate was 10.79%. 11
Injection of illicit drugs is an important high-risk factor for acquiring HIV-1 infection and contributes to the spread of the virus worldwide. 12 A number of earlier studies showed that intravenous use of opiates influences the outcome of HIV infection.12–14 Injecting drug users (IDUs) frequently abuse heroin, the most commonly abused opiate. Heroin injection increases the risk of acquiring HIV-1 infection 15 and progression to AIDS.13,16 Although the mechanism by which heroin promotes HIV infection and AIDS progression is still debatable, there is overwhelming in vitro evidence that morphine, the active metabolite of heroin, enhances the susceptibility of the immune cells to HIV infection as well as HIV replication in cells.17–21
A number of studies have revealed that the rate of HIV-1 drug resistance among IDUs is higher than that in other HIV-1-positive populations. A previous study by Salomon et al. reported that high-level resistance to antiretroviral drugs was observed in individuals newly infected by injecting drug use. 22 A series of studies by Kozal et al. indicated that antiretroviral resistance occurs in 10–15% of newly infected individuals and 30–50% of untreated IDUs.23–25 Gill et al. reported that IDUs had a 70% higher risk of resistance than nondrug users. 26 Kowalski et al.'s study found that primary PI-resistant mutations were more frequent among IDUs (24%) than non-IDUs (8%). 27 Pando's study also showed that the frequency of primary resistant mutations in female IDUs (33.3%) was much higher than that (8.4%) in the whole study groups. 28 A recent study in Belgium showed a particularly high prevalence of drug-resistant mutations among IDUs (29.4%) compared with 7.8% among all HIV-1 patients. 29
On the other hand, a few other studies indicated that there was no significant difference in HIV-1 drug resistance between IDUs and other populations, such as Weinstock's study among drug-naive HIV-1-infected persons in 10 US cities, 4 Moore's study in HIV-infected persons in care, 30 Paraskevis et al.'s investigation on newly diagnosed HIV-1 patients in Greece, 31 and Chang et al.'s research in untreated HIV-1 patients in Taiwan. 32 Thus, the influence of heroin addiction on HIV-1 drug resistance remains debatable.
Because of the extreme complexity of opiate addiction and/or HIV infection, it is extremely difficult to compare different clinical and epidemiological findings when studying the impacts of heroin abuse on HIV-1 drug resistance. Laboratory in vitro studies will yield relatively reliable data. Therefore, this study was conducted to evaluate the effect of morphine (the active metabolite of heroin) on the generation of HIV-1 drug-resistant mutations induced by antiretroviral drugs through an in vitro HIV-1-CD4+ T (HIV-1 III B-MT4) cell system. We selected two typical clinical antiviral drugs, including an NRTI (Lamivudine, 3TC) and an NNRTI (Nevirapine, NVP), to treat HIV-1-infected cells and observed the impacts of morphine on the occurrence of HIV-1 drug-resistant mutations. The findings will provide a better understanding about the effects of morphine on HIV drug-resistant mutations as well as viral replication, and will facilitate a rational basis for practical guidance toward the reduction of heroin abuse among IDUs.
Materials and Methods
Reagents
Morphine sulfate was obtained from the Department of Anesthesia, the first affiliated hospital of Guangxi Medical University. 3TC, NVP, and Naltrexone were purchased from Sigma Company. 3TC and NVP were dissolved in RPIM 1640 media at the concentration of 3.2 μM, filtered through a 0.22-μm filter (Millipore), and stored at −80°C.
Cell culture and HIV-1 strain
MT4 cell line (a human T4 cell line) 33 and a wild-type HIV-1 III B strain (HIV-1subtype B, a laboratory strain) 34 were kindly provided by Dr. Jinyun Li of the AIDS Research Centre, Chinese Academy of Military Medical Sciences. MT4 cells were maintained in 10% RPMI 1640 media (Corning Corp.) containing 10% fetal bovine serum. The viral stock preparation followed the standard procedure of HIV-1 viral stock preparation. 35 To exclude the possibility of preexisting drug-resistant mutations in the viral preparation, we have sequenced our viral stock and submitted the sequence to HIV DATABASE, showing that the wild viral stock contains no drug-resistant mutations (data not shown). The viral stock's 50% tissue culture infective dose (TCID50) is 20,000/ml.
Growth kinetic assay
Growth kinetic assay on HIV-1 III B virus was measured in MT4 cells as described previously 36 with a slight modification. Briefly, MT4 cells were plated in 48-well plates at a density of 1 × 106 cells/well in 0.5 ml of RPMI 1640 medium, and then 50, 100, 200, and 400 TCID50 of HIV-1 III B viruses were added, respectively. After incubation for 2 hours at 37°C, cells were washed twice with phosphate-buffered saline and resuspended in 10% RPMI 1640 media. The cultures were split every 3–4 days by replacing 50% of the media with the same volume of fresh media. Triplicate cultures were tested for each group, and viral growth was determined by HIV-1 p24 levels in culture supernatants on day 3–9 postinfection. The levels of HIV-1 p24 antigen were determined by enzyme-linked immunosorbent assay (PerkinElmer).
Infection of MT4 cells with HIV-1 III B strain
MT4 cells were infected with cell-free HIV-1 III B (200 TCID50/106 cells) for 2 hours at 37°C. The cells were then washed thrice with plain RPMI 1640 media (Corning Corp.) to remove unabsorbed virus, and then fresh media containing 10% fetal bovine serum were added to the cell cultures.
Morphine, naltrexone, and 3TC/NVP treatments
To observe the effect of morphine on HIV-1 infection and replication in MT4 cells, we incubated MT4 cells with or without morphine at different concentrations (10−12–10−6 M) for 24 hours before HIV-1 infection. The concentrations of morphine we used were comparable with the levels found in the drug user's plasmas or urine samples of heroin-using subjects.37,38 To investigate whether naltrexone antagonizes the activity of morphine, we used naltrexone (10−8 M) to treat MT4 cells for 1 hour before morphine treatment. The morphine- and/or naltrexone-treated cells were then infected with the same stock of HIV-1 III B strain (200 TCID50/106 cells). After washing the cells with plain RPMI 1640 media, fresh media containing morphine and/or naltrexone were added to the cell cultures. Cells and culture supernatants were collected for RT-PCR analysis and HIV-1 p24 assay at day 3–9 postinfection. Untreated cells and culture supernatants served as controls.
To determine the effect of morphine on HIV-1 drug-resistant mutations, cells were treated with a combination of morphine and antiretroviral drugs (3TC or NVP). Thus, MT4 cells were treated with or without morphine (10−6, 10−8, 10−10 M) for 24 hours, and then treated with 3TC 0.16 μM or NVP 0.32 μM for 2 hours. The concentrations of 3TC or NVP used in this study were based on an IC50 test, and were similar to those described in previous studies.34,39 The treated cells were infected with the same stock of HIV-1 III B strain (200 TCID50/106 cells) for 2 hours at 37°C in the presence or absence of the reagents described above. After washing the cells with plain RPMI media, fresh medium containing morphine and/or 3TC/NVP was added to cell cultures. The final washing medium was tested for HIV-1 p24 antigen to confirm that it was free of residual inoculum. Untreated cells served as controls. Culture supernatants were collected 7 days after infection. Of the supernatants collected from each well, 200 μl was used for HIV-1 RNA extraction, and 100 μl was used as parental virus to infect naive MT4 cells to generate progeny viruses. The infection lasted for 2 hours at 37°C. The cells were then washed thrice with plain RPMI media, and fresh media containing morphine and/or 3TC/NVP were added to the cell cultures. The reinfection procedure was repeated 12 times. Correspondingly, the supernatants collected during the different passages were described as passage 1 to passage 12, respectively.
Viral RNA extraction and RT-PCR
HIV-1 RNA was extracted from a 200 μl supernatant using a High Pure Viral RNA Kit (Roche Corp.) following the protocol provided with the kit. The cDNA (1,315 bp) of the HIV-1 pol gene was obtained by nested RT-PCR. Briefly, the isolated RNA was subjected to RT-PCR (Life Technologies Superscript One-Step RT-PCR for long templates) using the primers MAW-F26 and DO2-R1 (Table 1) with one cycle of 50°C for 30 minutes and 94°C for 2 minutes, and then 30 cycles of 94°C for 30 seconds, 55°C for 30 seconds, and 72°C for 3 minutes, followed by a final extension at 72°C for 7 minutes. The second-round PCR used the primers PRO-F1 and RTO-R2 (Table 1), with an amplification protocol of one cycle of 94°C for 2 minutes, followed by 30 cycles of 94°C for 30 seconds, 63°C for 30 seconds, and 72°C for 2.5 minutes, followed by one final cycle of 72°C for 7 minutes. The amplified cDNA products were harvested and confirmed by 1% agarose gel electrophoresis.
Sequencing
After confirmation and purification, the amplified products were sequenced by the Huada Genomics Company with the primers SEQ-F1, SEQ-R3, SEQ-F4, SEQ-R6, and SEQ-F7 (Table 1).
Determination of drug-resistant mutations
The sequences of the products were submitted to the Stanford HIV Drug Resistance Database (http://hivdb.stanford.edu). Resistant mutations and the extent of resistance to antiviral drugs 3TC and NVP were determined by comparing the sequences using this database.
P24 test
The supernatants of HIV-1-infected cell cultures were tested using an HIV-1 p24 antigen assay from Perkin Elmer, according to the manufacturer's recommendations.
Results
Virus growth kinetics
To obtain an appropriate viral inoculum concentration for the experiments, viral growth kinetics assay was performed first. As shown in Fig. 1A, infections using 50 TCID50 and 100 TCID50 resulted in a relatively low level of HIV-1 replication, whereas infection using 400 TCID50 was too strong to reflect the effect of morphine treatment. Therefore, we chose an appropriate viral inoculum (200 TCID50) to do the subsequent experiments.
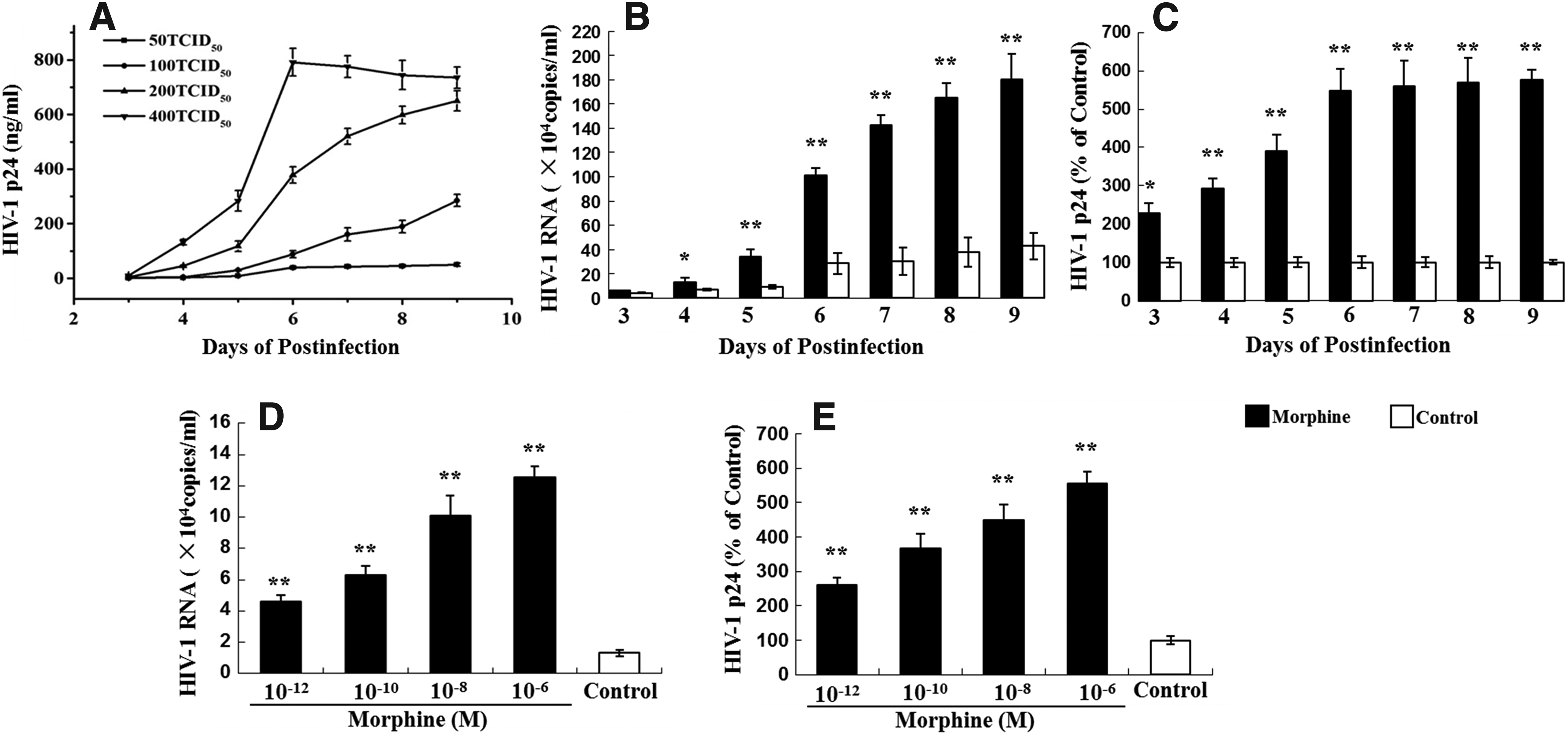
Viral growth kinetics, dose-dependent and time-course effects of morphine on HIV III B infection/replication in MT4 cells.
Morphine enhances HIV-1 replication of MT4 cells
We then determined the effect of morphine on HIV infection of MT4 cells. As shown in Fig. 1, morphine treatment resulted in an increased expression of HIV RNA (Fig. 1B, D) and viral p24 protein (Fig. 1C, E). This morphine-mediated increase in HIV RNA and viral protein expression was statistically significant (p < 0.05). These effects of morphine on HIV were dose and time dependent (Fig. 1B–E). The highest increase in HIV RNA expression was observed at a dose of 10−6 M morphine (Fig. 1D) at day 9 after infection (Fig. 1B). The enhancing effect of morphine could be abrogated by pretreatment with naltrexone, either at the HIV RNA level or at the viral protein level (Fig. 2).
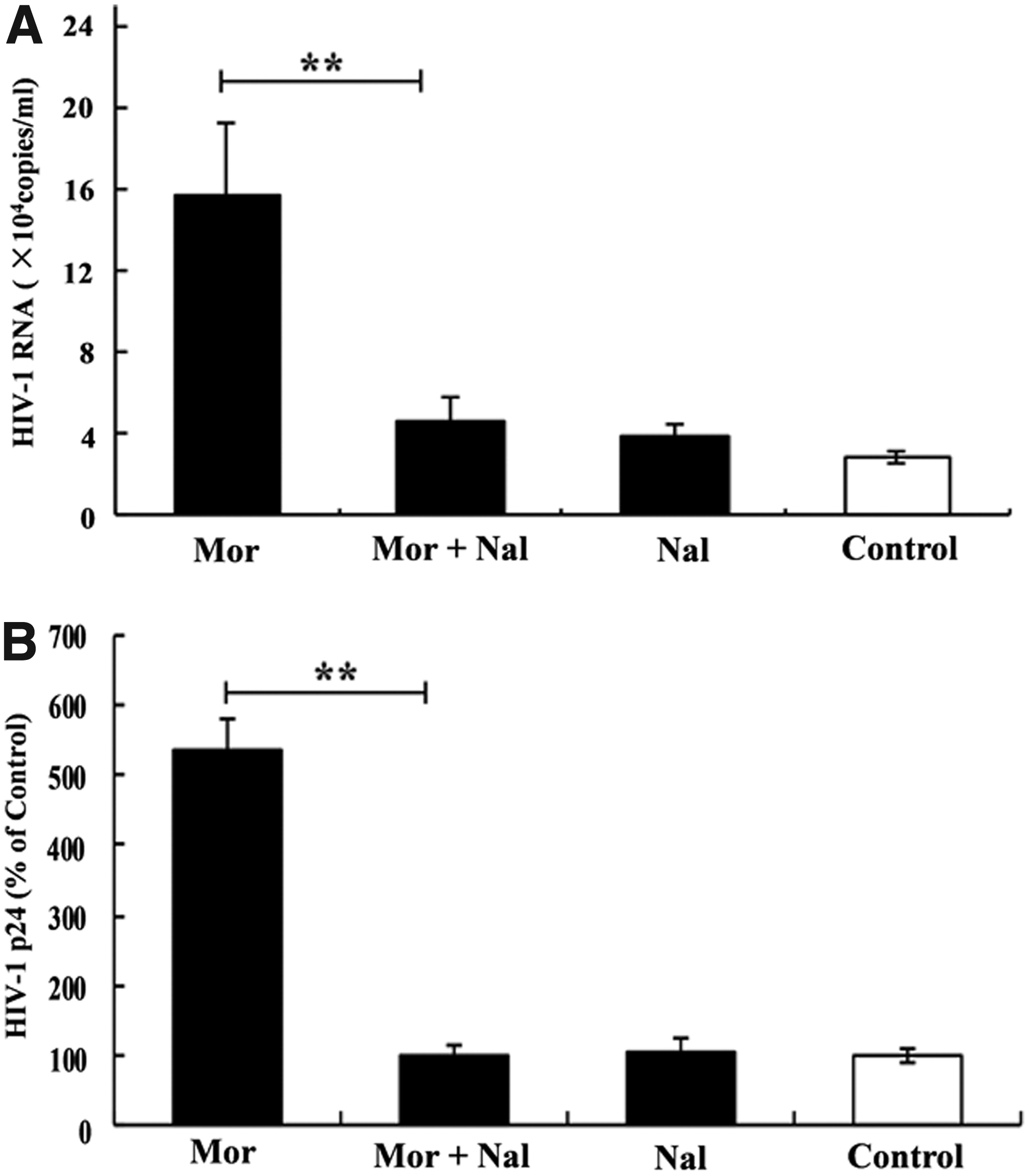
Effect of the opiate antagonist on morphine-mediated upregulation of HIV III B infection.
Effect of morphine on HIV-1 drug-resistant mutations induced by 3TC
After we determined that morphine could augment HIV replication in MT4 cells in vitro, we considered the possibility that morphine treatment might enhance 3TC or NVP induction of HIV-1 drug-resistant mutations in MT4 cells. Thus, we investigated the effects of morphine on HIV-1 drug-resistant mutations induced by 3TC or NVP. The effects of morphine on HIV-1 drug-resistant mutations induced by 3TC are shown in Table 2. Morphine/3TC treatment induced and generated more drug-resistant mutations than the 3TC treatment only. The morphine (10−6 M)/3TC treatment induced HIV-1 to produce V118I and M184I mutations and finally to generate V118I and M184I at the 12th passage (Table 2), whereas 3TC alone only induced M184I mutations (Table 2).
3TC, lamivudine; —, no mutation detected.
The effect of morphine on HIV-1 drug-resistant mutations was dose dependent. As the morphine concentration increased (10−10, 10−8, 10−6 M), more mutations were observed and generated (Table 2). In addition, in morphine alone (without 3TC) and control (without morphine and 3TC treatment) groups, no drug-resistant mutations were observed (Table 2).
We also compared the time of first appearance and the time of fixation of important mutations against that caused by 3TC, including M184I and V118I. As shown in Table 3, the morphine/3TC treatment induced and fixed the mutations at an earlier time compared with 3TC alone. For example, under the condition of 10−6 M morphine treatment, the first time of M184I and V118I appeared and the time of their fixation was at passage 3 (Table 3). However, in the 3TC alone group, the first time of M184I appeared and the time of their fixation was at passage 7 (Table 3).
—, no mutation detected.
Effect of morphine on HIV-1 drug-resistant mutations induced by NVP
The effects of morphine on HIV-1 drug-resistant mutations induced by NVP are shown in Table 4. Morphine/NVP treatment induced and generated similar drug-resistant mutations to NVP alone. The morphine/NVP (morphine 10−6 M) treatment induced HIV-1 to produce G190A, Y181C, and Y188C mutations and finally to generate Y181C and Y188C at the ninth passage (Table 4). NVP alone also induced Y181C and Y188C mutations and finally fixed the mutations at the 12th passage (Table 4). However, there was a significant difference between morphine/NVP and NVP-alone groups in the time of first appearance and the time of fixation of these mutations (Table 5). For example, under the condition of 10−6 M morphine treatment, Y181C and Y188C first appeared at passage 6 and 4, respectively, and the time of fixation of both was at passage 7. In the NVP-alone group, Y181C and Y188C first appeared at passage 6, and the time of fixation of both was at passage 12.
NVP, nevirapine; —, no mutation detected.
—, no mutation detected.
The effect of morphine on time of first appearance of mutations was dose dependent. As morphine concentration increased (10−10, 10−8, 10−6 M), the mutations were observed and generated at earlier passages (Table 4). In addition, no drug-resistant mutation was observed in the morphine-alone and control groups (without morphine or NVP treatment) (Table 4). Furthermore, we observed quite a few mixed strains in the morphine and/or NVP treatments. Thus mutated and wild strains coexisted in the same passage, for example, Y181CY, Y188CY, and G190AG (Table 4).
Discussion
In this study, we examined the impact of morphine on HIV-1 infection of MT4 cells. We demonstrated that when exposed to morphine, MT4 cells were more susceptible to HIV-1 infection than unexposed cells, as demonstrated by the increased HIV RNA and p24 levels in exposed cell cultures (Fig. 1). The action of morphine on HIV-1 appeared to be mediated through an opioid receptor, as an opioid receptor antagonist blocked the effect of morphine (Fig. 2). The results are consistent with those of previous studies on the effect of morphine on HIV-1 infection and replication in a variety of immune cells, including peripheral blood mononuclear cells (PBMCs), 17 neonatal monocyte-derived macrophages, 19 human blood mononuclear phagocytes, 18 CD8+ T cells, 40 and human macrophages. 21
Because CD4+ T cells represent the cellular reservoir in which HIV is harbored predominantly in the body and contribute to most of the viral replication detected in blood plasma,41,42 our finding that morphine enhances HIV-1 infection of MT4 cells, a CD4+ T-cell line, provides the strongest in vitro evidence to date to support the enhancing effect of morphine on HIV replication. In addition, an animal study also observed that chronic morphine exposure enhanced viral replication and accelerated the onset of AIDS in simian immunodeficient virus-infected Indian rhesus macaques. 43
As an RNA retrovirus, HIV shows a high degree of genetic variability due to virally encoded reverse transcriptases that do not possess proofreading capability. The estimated mutation rate for HIV is ∼10−4–10−5 mutations per nucleotide and cycle of replication.44,45 In addition, HIV replication is very rapid and up to 1010 virions can be produced daily in an untreated HIV-infected individual.46,47 Therefore, the generated viral quasispecies are composed of many HIV variants, including drug-resistant strains, and the chances of developing mutations involved in drug resistance are very high.
Although the mechanism(s) by which morphine enhances HIV-1 infection and replication is still debatable, including inducing the expression of key HIV entry coreceptors CCR5 and CXCR4, morphine-mediated inhibition of the IFN signaling pathways, inhibition of apolipoprotein B mRNA-editing enzyme catalytic polypeptide-like 3 (APOBEC3), or anti-HIV microRNAs' expression,17–21 there is overwhelming in vitro evidence that morphine enhances the susceptibility of the immune cells to HIV infection and HIV replication in cells.17–21 In this sense, we suppose that the enhancement of HIV replication by morphine may result in the increase of HIV-1-resistant mutations, because higher levels of HIV-1 replication means more HIV transcriptional errors, which in turn lead to more HIV-1-resistant mutations.
In our investigation on the effect of morphine on the generation of HIV-1 drug-resistant mutations in the context of antiretroviral drug selective pressure, our results showed that morphine treatment not only induced more resistant mutations under selective pressure but also shortened the generation time (Tables 2–5). These findings are extremely important in the context of understanding the epidemiological antiretroviral resistance in IDUs, indicating that heroin (morphine) abuse is an independent factor contributing to the generation of HIV-1 drug-resistant mutations, despite lower adherence to HAART, which is a commonly accepted factor contributing to the higher antiretroviral resistant rate in IDUs than in other HIV-1-positive populations.
Aside from enhancement of HIV infection and replication by morphine, other molecular mechanism(s) cannot be ruled out, for example, the impact of morphine on cellular APOBEC. Human APOBEC enzymes with the cytidine deaminase activity have been shown to cause G-to-A hypermutagenesis and are directly related to HIV-1 nucleotide mutations. 48 Whether, and how, morphine acts on cellular APOBEC would be an interesting topic for future study.
Previous studies have found that all resistant mutations are negative for the ability to replicate.49,50 For example, the codon 184 mutation can decrease the replication capacity of the virus by 3–10%50,51 and even up to 48–57%, 52 compared with the wild type. Consistent with these results, our study found that a typical mutation of NVP, G190A, appeared in only two passages and was then lost, even under the selective pressure of NVP (Table 4). However, the fixation of resistant mutations requires only 5–6 passages for NVP treatment under the condition of optimized morphine inducement. This result, combined with our finding that many more drug-resistant mutations were generated by morphine treatment than by antiretroviral drug use only (Tables 2 and 4), implies that continuing to take heroin during clinical antiretroviral therapy would greatly enhance the occurrence of resistant strains as well as shorten their generation time.
A confusing result in our experiment is that among two mutations, V118I and M184I, which appeared in the morphine plus 3TC treatment group, only M184I mutation was reported to be selected by 3TC. Why does V118I, a non-NRTI-resistant mutation, appear simultaneously along with M184I mutation? We could not provide a reasonable explanation.
To confirm our results, we repeated the experiments and the results are presented as Supplementary Data (Supplementary Tables S1–S3; Supplementary Data are available online at www.liebertpub.com/mdr). In the repeated experiments, the similar results were observed in morphine plus 3TC or NVP treatment group (Supplementary Tables S1–S3). Although the results of repeated experiments were not the same to those of the original one, they support the conclusion that morphine treatment not only induced more drug-resistant mutations under selective pressure from antiretroviral drugs but also shortened the mutations' generation time. One possible reason for this phenomenon is that higher levels of HIV-1 RNA replication induced by morphine increase base pairing errors, which lead to simultaneous occurrence of various non-NRTI mutations along with M184I mutation.
Taken together, our study provides compelling experimental evidence that morphine increases HIV-1 replication/infection and drug-resistant mutations in vitro. Although additional mechanism(s) might be involved in the effect of morphine on resistant mutations, the increase in HIV-1 infection and replication likely account for much of the morphine-mediated increase in resistant mutations in HIV-1-infected MT4 cells. Determining the clinical relevance of these in vitro findings as well as exploring other mechanisms will be the subject of further research.
Footnotes
Acknowledgments
The study was supported by National Natural Science Foundation of China (NSFC, 81171624, 81360259, 31360033, and 81460511), Guangxi Scientific and Technological Development Project (Gui Ke Gong 14124003-1), Key Project of Guangxi Universities Scientific Research (2013ZD012), Guangxi University “100-Talent” Program and Guangxi University innovation team and outstanding scholars program (Gui Jiao Ren 2014[7]), Guangxi Scientific and Technological Project (2013GXNSFBA01966), and Youth Science Foundation of Guangxi Medical University (GXMUYSF201344).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.