
Abstract
Salmonella Enteritidis strains that are resistant to nalidixic acid and exhibit reduced susceptibility to fluoroquinolones have been increasing worldwide. In Brazil, few studies have been conducted to elucidate the quinolone resistance mechanisms of S. Enteritidis strains. This study analyzed the profile of gyrA, gyrB, parC, and parE mutations and plasmid-mediated quinolone resistance (PMQR) mechanisms in S. Enteritidis NalR strains isolated in Brazil. Moreover, the minimum inhibitory concentrations (MICs) of ciprofloxacin were evaluated in 84 NalR strains and compared with 20 NalS strains. The mutation profiles of the gyrA gene were accessed by high-resolution melting analysis and gyrB, parC, and parE by quinolone resistance-determining region sequencing. The MICs of ciprofloxacin were accessed with Etest®. The strains were divided into five gyrA melting profiles. The NalR strains exhibited the following amino acid substitutions: Ser97→Pro, Ser83→Phe, Asp87→Asn, or Asp87→Tyr. The average MICs of ciprofloxacin was 0.006 μg/ml in the NalS and 0.09 μg/ml in the NalR strains. No points of mutation were observed in the genes gyrB, parC, and parE. The qnrB gene was found in two strains. In conclusion, the reduced susceptibility to ciprofloxacin observed in NalR strains may cause treatment failures once this drug is commonly used to treat Salmonella infections. Moreover, this reduced susceptibility in these Brazilian strains was provided by target alteration of gene gyrA and not by mobile elements, such as resistance plasmids.
Introduction
G
In the United States, Salmonella is estimated to cause every year 1 million foodborne illnesses with 19,000 hospitalizations and 380 deaths. Recent reports show that in 2014 serovar Enteritidis was the most isolated, representing 19% of the Salmonella isolates in that country. 4 In European countries, among the 82,694 confirmed salmonellosis cases in 2013, serovar Enteritidis was the most isolated, representing 39.5% of the reports. 5
The use of antimicrobials is not recommended in cases of gastroenteritis, which are usually self-limiting. However, in some cases, the infection might lead to serious infections, and the antibiotic therapy might be necessary.6,7
Nalidixic acid was the first quinolone and was widely used in the early 1960s to treat Gram-negative bacterial urinary tract infections. Further modifications in the chemical structure of these drugs, such as the addition of a fluorine atom, increased the efficacy of this group.6,8–11 The drug of choice for the treatment of Salmonella infections is typically ciprofloxacin due to its broad-spectrum antimicrobial activity. In the veterinary field, fluoroquinolones have been used for the treatment of infections or as growth promoters in food production animals.7,12
The extensive use of these antimicrobials has led to increasing numbers of non-typhi Salmonella strains that are resistant to quinolones and exhibited reduced susceptibility to fluoroquinolones.10,13–19 This reduced susceptibility can lead to treatment failures in some cases.13,15
It has been reported that quinolone resistance among Salmonella serovars has been increasing over the years in several countries, such as the United States, European countries, Southeast Asia, among others. 20 In the United States, around 3% of all non-typhoidal Salmonella isolated between 2009 and 2011 presented some level of resistance to ciprofloxacin. 21
Quinolones target the bacterial DNA gyrase; this enzyme is a type II topoisomerase that is essential for bacterial DNA replication. 22 This enzyme consists of 2A and 2B subunits encoded by gyrA and gyrB genes, respectively. The second target of the fluoroquinolones is the topoisomerase IV enzyme, which is codified by the genes parC and parE.7,8,20,22
The mechanisms linked to quinolone resistance in Salmonella include changes in the binding site of the antimicrobial to the enzyme. These changes are related to mutations in the quinolone resistance-determining region (QRDR) of the target genes. These points of mutation can lead to amino acid substitutions that result in different levels of quinolone resistance.7,19,23,24
Although four target genes exist, gyrA is the main gene involved in quinolone resistance, and a single point of mutation in the QRDR of this gene typically results in resistance to nalidixic acid and reduced susceptibility to fluoroquinolones. Two or more mutations in the gyrA gene or in the other topoisomerase genes confer resistance to ciprofloxacin. The QRDR of DNA gyrase A in S. enterica is localized between amino acids 67 and 122.23,25
Other resistance mechanisms include the qepA, oqxA, and oqxB genes that codes an efflux pump that does not permit the intracellular accumulation of the antimicrobial agent, the actions of the plasmid gene qnr that codes a group of pentapeptide proteins that bind to DNA gyrase and prevent the action of quinolones, and the action of the aminoglycoside acetyltransferase that is coded by gene aac(6′)-Ib-cr and can reduce susceptibility to ciprofloxacin.7,19,22,26–28
Recent studies from our research group regarding S. Enteritidis strains isolated from humans, food, and chickens in Brazil revealed a high level of nalidixic acid resistance, particularly in poultry-related strains.29,30 However, the levels of resistance to fluoroquinolones and the mechanisms of quinolone resistance were not evaluated in those studies. In Brazil, the only study of quinolone resistance mechanisms was conducted with Salmonella strains from several serovars that were isolated only in the Paraná State over the period of 8 years from 1999 to 2007. 31
Thus, the objective of this study was to analyze the profile of gyrA mutations and investigate the presence of some plasmid-mediated quinolone resistance (PMQR) genes that are related to quinolone resistance, such as qnr genes, qepA, and aac(6′)-Ib-cr, in nalidixic acid-resistant (NalR) S. Enteritidis strains isolated in Brazil. The studied strains were isolated from humans, food, chickens, and farm environments in five Brazilian States in four regions over a 44-year period. Moreover, the minimum inhibitory concentrations (MICs) of ciprofloxacin for NalR S. Enteritidis strains were evaluated and compared with those of nalidixic acid-susceptible strains (NalS). These data were analyzed to evaluate the prevalent points of mutation in the gyrA gene, and to verify the presence of other mechanisms of quinolone resistance in S. Enteritidis strains from Brazil.
Materials and Methods
Bacterial strains
A total of 84 NalR Salmonella Enteritidis strains were studied (Table 1). These strains were composed of the nalidixic acid-resistant strains that were identified among 188 S. Enteritidis strains that were studied in previous works by our research group.29,30 Additionally, four NalR strains isolated in 1969, 2012, and 2013 were included in this study (Table 1), which were identified among 26 new strains received by our laboratory and isolated between 1968 and 1993 and in 2012 and 2013 (unpublished data). The strains were isolated from human feces (24), food (16), chickens (29), and farm environments (15) (Table 1). Moreover, 20 nalidixic acid-sensitive (NalS) S. Enteritidis strains that were representative of the four sources and years of isolation were used for comparisons with the resistant strains (Table 1). Those strains were provided by two Salmonella spp. References Laboratories in Brazil that were the Adolfo Lutz Institute of Ribeirao Preto (IAL-RP) and Oswaldo Cruz Foundation (FIOCRUZ) and by AVIPA (Avicultura Integral e Patologia S/A).
The 26 strains with reduced susceptibility to ciprofloxacin presented MICs between 0.12 and 0.5 μg/ml and were isolated from the four sources in the States of São Paulo, Mato Grosso, Pernambuco, and Paraná in 1996, 1999, 2000, 2001, 2004, 2005, 2006, 2008, 2009, and 2010.
Strains isolated from food include mayonnaise, sandwich, cake, pork, raw chicken, papaya, raw fish, and beef.
MICs, minimum inhibitory concentrations.
Detection of mutations in the QRDR of gyrA, gyrB, parC, and parE
Mutations in the QRDR of the gyrA gene were searched in the 84 NalR and 20 NalS strains of S. Enteritidis (Table 1). Initially, the different mutation profiles were assessed through high-resolution melting analysis (HRMA) using the general procedure described by Souza and Falcão 32 with a few modifications. The primers used were identical to those presented by Slinger et al. 33 and amplified a 89-bp fragment of the QRDR in the gyrA gene of S. enterica (Table 2).
HRMA, high-resolution melting analysis.
Briefly, HRMA is considered to be a fast, cost-effective, sensitive, specific, and high-throughput method for the interrogation of single-nucleotide polymorphisms (SNPs), hypervariable repeat regions in polymerase chain reaction (PCR) products, and also for the discovery of new SNPs. 34 It consists of a one-step, closed-tube, post-PCR assay that detects sequence variations within specific loci through accurate monitoring of the reduction in the fluorescence of a PCR product stained with a double-stranded specific fluorescent dye to monitor the transition from unmelted to melted DNA. In contrast to traditional melting analysis, the information of the HRMA method is contained in the shape of the melting curve, rather than just in the calculated melting temperature. 35
Heteroduplexes were generated to separate the profiles that exhibited mutation points with complementary bases (e.g., A-T and T-A) through the addition of an exogenous DNA sample with a known sequence.
Representative strains of each group generated by HRMA were sequenced to analyze the type of mutation in each profile. Moreover, representative strains of each group comprising 20 NalR and 10 NalS strains had their genes gyrB, parC, and parE sequenced to check possible mutation points in the QRDR of these genes. The general procedure for sequencing was the same as that described by Souza et al. 36 and used the primers gyrA1, stgyrA2, SgyrB.1, SgyrB.2, parC.Sal, parC.Sal2, SparE.1, and SparE.2 (Table 2).19,33,37 Each forward and reverse strand was sequenced at least two times.
Sequence analysis
The resulting DNA sequence of the QRDR of each strain described in the previous section was checked for quality by visual inspection with the aid of the ChromasPro version 2.33 software (Technelysium Pty. Ltd.) and trimmed to remove the low-quality nucleotide sequences from the ends. This process generated sequences with the same number of nucleotides that were consequently of the same size.
Detection of PMQR genes
The genomic DNA of the 84 studied strains (Table 1) was extracted as described by Campioni and Falcão. 38 The primers used in the reaction, the annealing temperatures, and the references for the primers used are described in Table 2.10,39–42 The genes under study were the qnrA, qnrB, qnrC, qnrD, and qnrS genes, which are code for a group of pentapeptide proteins, the qepA, oqxA, and oqxB, which are code for efflux pumps, and the aac(6′)-Ib-cr, which codes for an aminoglycoside acetyltransferase.
The applied reaction conditions included an initial 5-min denaturation at 94°C, followed by 30 cycles consisting of denaturation at 94°C for 45 sec, the application of the annealing temperature of each primer for 45 sec, and extension at 72°C for 1 min. After the last reaction, a final extension step of 72°C for 15 min was applied.
To evaluate the reproducibility of the experiments, the PCR reactions were repeated twice for some strains. The strains Klebsiella pneumonia KP15 and Escherichia coli 16-00 were used as positive controls for the genes qnr. The strain E. coli 78-01 and Salmonella Typhimurium 138.12 were used as positive control for the genes aac(6′)-Ib-cr and oqxA/B, respectively. Reactions without DNA as a template were used as negative controls. The PCR products were analyzed through agarose gel electrophoresis and visualized by ultraviolet light after staining the gel with ethidium bromide (1.0 μg/ml).
Determination and analysis of the MIC of ciprofloxacin
MIC of the ciprofloxacin for the 84 NalR and 20 NalS S. Enteritidis strains studied were determined using Etest® and the interpretation following the guidelines of the Clinical and Laboratory Standards Institute. 43 Strains with MIC ≤0.06 μg/ml were considered sensitive and ≥1 μg/ml resistant.
Results
Detection of the mutations in the QRDR of gyrA
The resulting melting curves for the 84 NalR strains and the 20 NalS strains allocated the strains into four different melting profiles (Fig. 1). All 20 (100%) of the NalS strains were grouped in the same profile named A, and the NalR strains comprised the other three profiles. The resistant profile B comprised only one strain, whereas the profile C comprised 15 (17.8%) strains. Profiles D and E were initially grouped in the same melting curve because they presented mutations of complementary bases (A-T and T-A). Thus, a reaction involving the formation of heteroduplexes was necessary to divide these profiles (Fig. 2). After the reaction, the profile D presented 13 (15.4%) strains, and profile E presented 55 (65.4%) strains. The profiles found did not reflect either the source of the strains or the year of isolation.
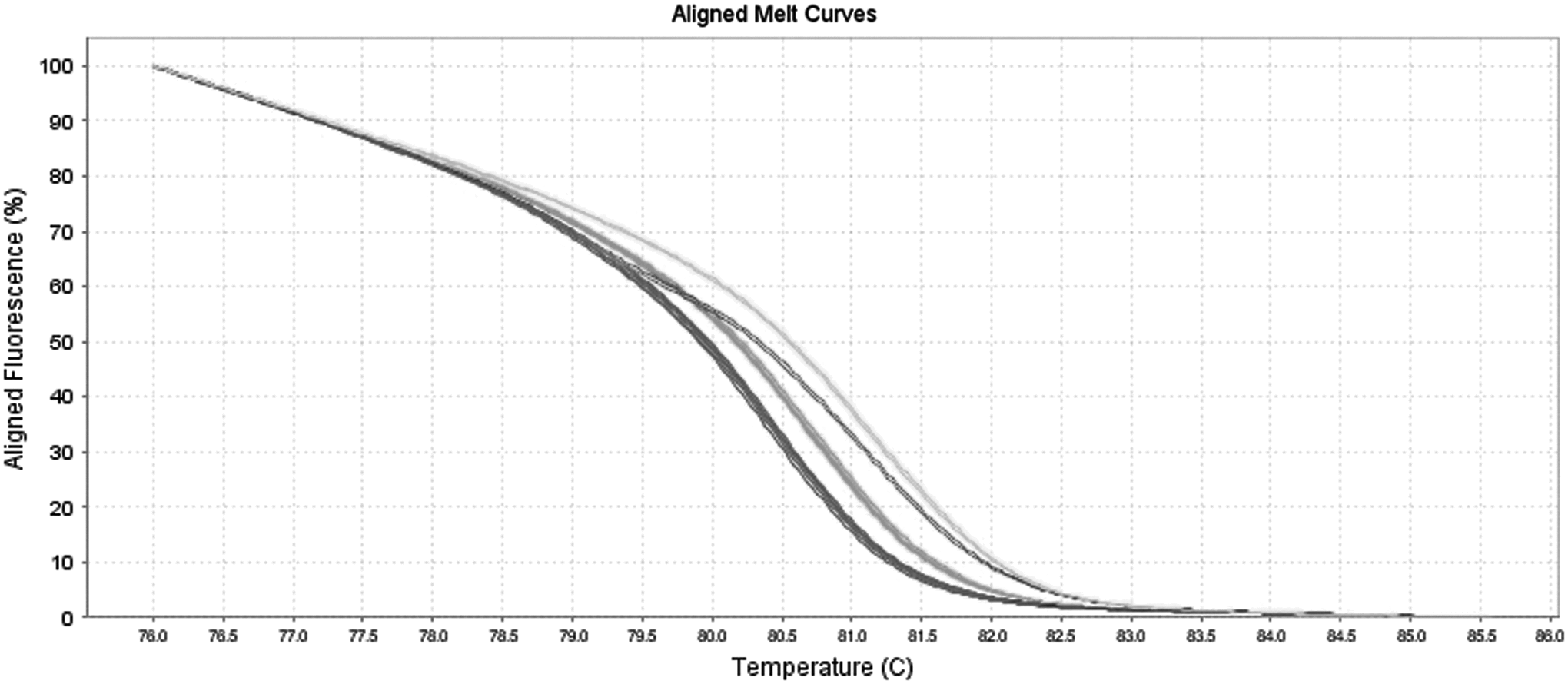
Melting profiles of the 84 nalidixic acid-resistant and 20 nalidixic acid-sensitive Salmonella Enteritidis strains. Black curve: profile A sensitive strains; dashed black curve: profile B resistant strains; gray curve: profile C resistant strains; and dashed gray curve: profile D and E resistant strains.
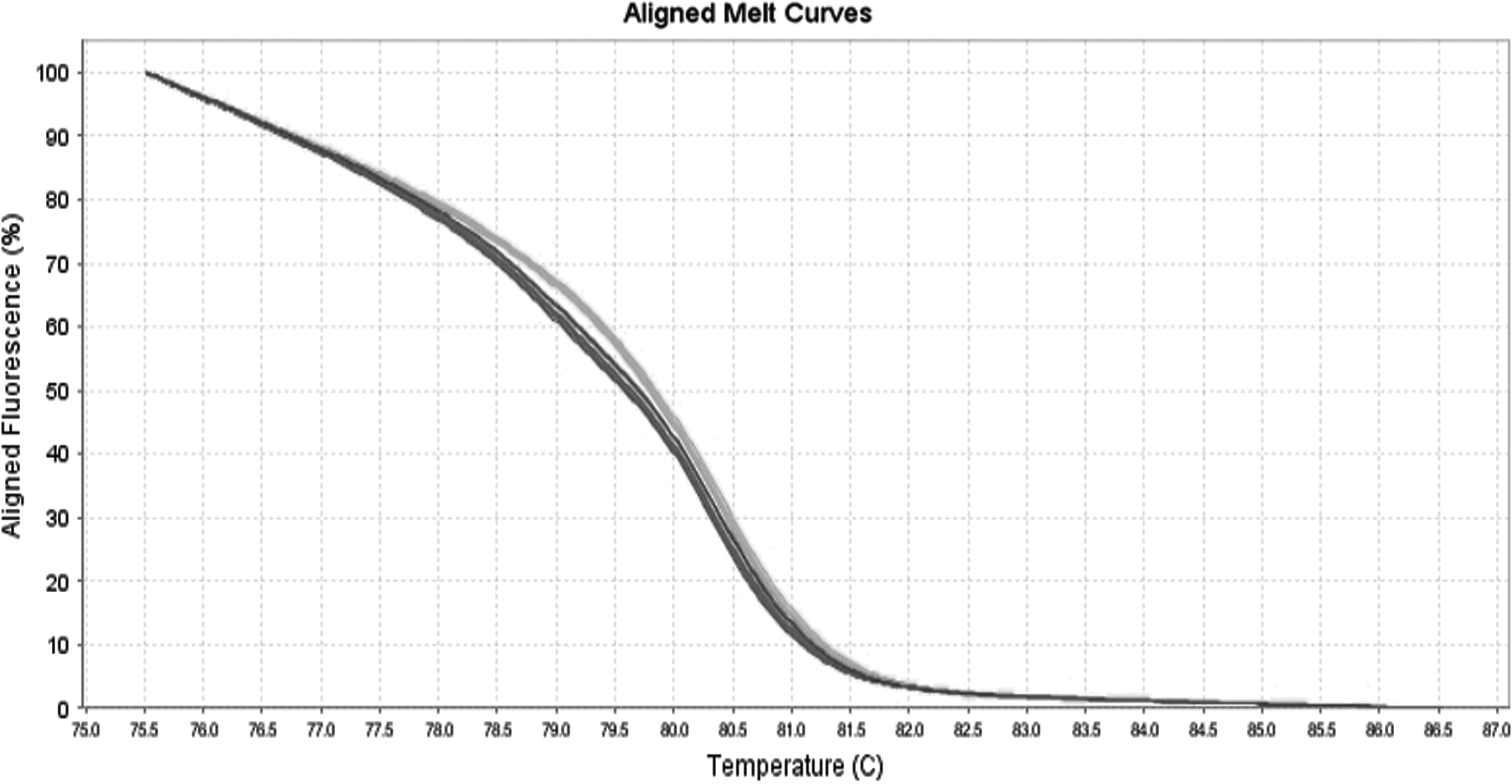
Melting profiles of the D and E profiles after separation through the production of heteroduplexes. Gray curve: profile D resistant strains; and dashed gray curve: profile E resistant strains.
Representative strains of each group were sequenced and exhibited point mutations in the QRDRs of the gene gyrA in NalR strains relative to the NalS strains. No points of mutation were observed in the genes gyrB, parC, and parE. The points of mutation in the gene gyrA were observed in the codons serine 83 (Ser83), asparagine 87 (Asp87), or serine97 (Ser97) of the sensitive strains.
Profile B presented a single T to C base point mutation in the trio of bases that codes amino acid serine 97 that resulted in a change to proline. Profile C presented a single C to T point mutation in the trio of bases that codes amino acid serine 83 and resulted in a change to phenylalanine. Profile D presented a single G to A point mutation in the trio of bases that codes the aspartic acid 87 and resulted in a change to asparagine. Finally, profile E presented a single G to T point mutation in the trio of bases that codes the amino acid aspartic acid 87 that resulted in a change to tyrosine.
Two (2.5%) NalR strains that carried the qnrB gene were both in profile E and did not exhibit differences in susceptibility compared with other strains of the group.
Detection of PMQR genes
None of the 84 NalR and the 20 NalS strains carried the qnrA, qnrC, qnrD, qnrS, qepA, oqxA, oqxB, or aac(6′)-Ib-cr genes. Only two strains (2.5%) carried the qnrB gene. These strains (107/09 and 191/10) were isolated from chickens in 2009 and 2010.
Determination and analysis of the MIC of ciprofloxacin
A comparison of the MICs of the 84 NalR strains according to the different point mutations and the MICs of the 20 NalS strains is presented in Table 3. All the 20 (100%) NalS strains and 58 (69.0%) NalR strains studied were sensitive to ciprofloxacin (MIC ≤0.06 μg/ml), whereas 26 (30.9%) NalR strains presented as intermediate to this drug (MIC 0.12 − 0.5 μg/ml). Among the HRMA-resistant profiles, the C (Ser83→Phe) profile exhibited the greatest reduction in susceptibility.
Asn, aspartic acid; Asp, asparagine; Phe, phenylalanine; Pro, proline; Ser, serine; Tyr, tyrosine.
Discussion
Resistance to quinolones has been increasing in Salmonella in recent years due to the extensive use of fluoroquinolones in both human and veterinary medicine.7,12 This fact is concerning because treatment failure might occur once nalidixic acid-resistant strains exhibit reduced susceptibility to fluoroquinolones.10,18,20
In the published studies from our research group based on Salmonella Enteritidis strains isolated in Brazil between 1986 and 2010, nalidixic acid resistance was observed only in the strains isolated after 1996.29,30 However, a recent analysis with additional strains isolated from 1968 to 1993 and 2012 to 2013 revealed that a single strain isolated in 1969 from humans was resistant to nalidixic acid in addition to three strains isolated in 2012 and 2013 (unpublished data). Thus, our study comprises NALR strains isolated over a 44-year period in Brazil and includes strains isolated close to the introduction of nalidixic acid into clinical use in 1962. 8
The MICs of the fluoroquinolone, ciprofloxacin, for the 84 Salmonella Enteritidis NalR strains and the 20 NalS strains were determined. The MICs of the strains for ciprofloxacin were all <1 μg/ml, however, even when these MICs were compared in NalS and NalR strains, the average was 0.006 μg/ml with a range of 0.002 – 0.008 μg/ml for NalS strains, whereas for NalR strains was 0.09 μg/ml with a range of 0.047 – 0.19 μg/ml (Table 3). These results indicate a reduction in the susceptibility of the NalR strains to ciprofloxacin compared with the NalS strains. Moreover, among the 84 NalR strains, 26 presented an intermediated MIC for ciprofloxacin.
Similar results have been found in studies performed in other countries, such as Spain, Korea, and the United Kingdom, involving Salmonella strains belonging to various serovars, including Enteritidis.10,44–47 In Brazil, Souza et al. examined strains isolated only in Parana State from 1999 to 2006 and also found reduced susceptibility to ciprofloxacin in the NalR strains compared with the NalS strains. 48
The main mechanism of quinolone resistance is thought to be due to target alterations of DNA gyrase and topoisomerase IV. However, a single point mutation in the QRDR of the gyrA usually results in resistance to nalidixic acid. Additional point mutations in gyrA, gyrB, parC, or parE genes typically result in high levels of resistance to these drugs. 20
In the present study, only gyrA gene presented points of mutation in the QRDR, which may explain the fact that those strains presented only reduced susceptibility to ciprofloxacin and not resistance. Four different gyrA point mutations were found in the gyrA gene among the NalR S. Enteritidis strains relative to the NalS strains, and these point mutations resulted in amino acid substitutions in the QRDR of this gene. The most frequent amino acid substitution was Asp87→Tyr, which was present in 55 of the 84 strains studied. Similar results were reported by Escribano et al. 45 who found this mutation in 18 of 20 NalR S. Enteritidis strains isolated in Spain. In contrast, Ferrari et al. studied Salmonella strains isolated in Parana State, Brazil and found that the most common point mutation in the gyrA gene was Asp87→Asn, which was present in 20 of the 64 S. Enteritidis strains studied. 31 The same frequency was found by Tamang et al., in a study of Salmonella strains isolated in Korea; these authors found the Asp87→Asn mutation in 17 of the 53 strains studied. 10 In the present study, the Asp87→Asn mutation was the third-most common mutation among the NalR strains (13 of 84 strains).
Regarding the mutation that produced the greatest decrease in ciprofloxacin susceptibility in the present study, the Ser83→Phe mutation was in 15 of the 84 strains, and the strains with this mutation exhibited an average MIC of 0.138 μg/ml. This mutation was also the same mutation that provided the greatest decrease in ciprofloxacin susceptibility among the strains studied by Tamang et al. that found this mutation only in human strains, which presented an average MIC of 0.25 μg/ml. 10 Furthermore, the other point mutations found in the work mentioned above, that is, Asp87→Asn, Asp87→Gly, and Asp87→Tyr, resulted in similar MICs of ciprofloxacin (0.12–0.25 μg/ml). 10
Other genes, such as those that code efflux pumps, aminoglycoside acetyltransferase, and the plasmid genes qnr, can provide quinolone resistance independently of mutations in the target genes.19,22,27,28 In the present study, the only PMQR gene found was qnrB, which was present in two of the 84 studied strains. However, these strains also carried the Asp87→Tyr mutation in the gyrA gene, and their MICs did not differ from those of the other strains in the same group that did not carry qnrB, which might suggest that the mechanism that caused quinolone resistance in these cases may be due to the target alteration of the gyrA gene and not qnrB. However, further conjugation experiments should be performed to show that the transfer of plasmid harboring qnrB gene does not change the MIC of the recipient strain.
Similarly, Tamang et al. failed to detect the abovementioned qnr genes in S. Enteritidis strains isolated in Korea. 10 However, other researchers have identified PMQR genes that cause resistance in Salmonella Enteritidis strains. Ferrari et al. detected the qnrA1 gene in one strain of S. Enteritidis that was isolated in Brazil and was resistant to NAL without mutations in the QRDR. 31 This report was the first of this gene in Salmonella strains from Brazil. 31 Wasyl et al. found eight S. Enteritidis strains isolated in Poland bearing the genes qnrB or qnrS. 47 Zhang et al. found 15 S. Enteritidis strains isolated in China that carried the aac(6′)-Ib-cr gene. 27
In conclusion, the reduced susceptibility to ciprofloxacin observed in NalR strains may cause treatment failures once this drug is commonly used to treat Salmonella infections. Moreover, this reduced susceptibility in these Brazilian strains was provided by target alteration of gene gyrA and not by mobile elements, such as resistance plasmids.
Footnotes
Acknowledgments
The authors thank São Paulo Research Foundation - FAPESP (Proc. 2008/57478-1) for financial support. During the course of this work, F.C. was supported by a scholarship from FAPESP (proc. 2009/09998-9). The authors thank J.D.D. Pitout (University of Calgary, Calgary, Canada) for kindly providing the control strains Kp15 and E. coli 16-00 and Dr. Monique Ribeiro Tiba Casas (Adolfo Lutz Institute, São Paulo, Brazil) for kindly providing the control strain Salmonella Typhimurium 138.12.
Disclosure Statement
No competing financial interests exist.