
Abstract
Since the implementation of routine PCV13 immunization in Japan, nonvaccine serotypes (NVTs) have been increasing among clinical isolates of Streptococcus pneumoniae. In this study, susceptibility to 18 antibiotics was tested for all the 231 isolates with NVTs, which were collected from children <16 years of age in northern Japan in 2014 (July–November). High resistance rates were observed for macrolides (>90.9%), tetracycline (91.3%), and clindamycin (75.3%), while penicillin (PEN) nonsusceptibility (PNSP; MIC ≥0.12 μg/ml) was detected in 42.9% of the pneumococci [39.4%; PEN-intermediate S. pneumoniae (PISP), 3.5%; PEN-resistant S. pneumoniae (PRSP)]. All serotype 15A isolates were PRSP (MIC, ≥2 μg/ml) or PISP, and PNSP was prevalent in also serotypes 23A (96.9%), 6C (41%), and 35B (33.3%). Overall, 42.0% of the isolates showed multidrug resistance (MDR). Sequence types (STs) determined for 20 PNSP isolates with NVTs were ST63 (15A), STs 242 or 5832 (6C), STs 338 or 5242 (23A), and ST558 (35B). All the PNSP isolates possessed tet(M), and erm(B) or mefA(A/E), and 70% of them were gPRSP having three altered genes pbp1a, pbp2x, and pbp2b. Among alterations in transpeptidase-coding region of penicillin-binding proteins (PBPs), two substitutions of T371S in the STMK motif and TSQF574–577NTGY in PBP1a were common to all PRSP isolates. The present study showed the spread of PNSP in NVTs 15A, 23A, 6C, and 35B, and the emergence of the MDR international clone Sweden15A-ST63 in northern Japan.
Introduction
P
An increase in NVTs in pneumococcal diseases is associated with the rising prevalence of nonsusceptibility to penicillin (PEN) and erythromycin, 5 and the emergence of NVTs with multidrug resistance (MDR) has become an international concern. Increase in the prevalence of the NVTs with resistance to several antibiotic classes, especially β-lactams, was observed in the United States6,7 and Canada. 8
The prevalence of penicillin-nonsusceptible pneumococci (PNSP) combined with resistance to more antibiotic classes has been recorded and updated since 1997 on the website of pneumococcal molecular epidemiology network (PMEN) (http://web1.sph.emory.edu/PMEN/), where the classification of resistant clones and antibiotic-susceptible clones worldwide is also shown. Resistance to β-lactams in pneumococci is related to sequence polymorphism in penicillin-binding proteins (PBPs). 9 Of the six pbp genes encoding PBPs, high-level resistance to penicillin is mainly due to mutations of the three pbp genes, pbp1a, pbp2x, and pbp2b, which reduce their affinity to β-lactams. 10 However, information regarding pbp gene sequences of penicillin-nonsusceptible NVT pneumococcus is limited.
In Japan, estimated vaccination rates increased 50–60% in the PCV7 voluntary immunization period in 2011 to 89.2% in the PCV13 routine immunization period in 2014,11,12 accordingly, a significant increase in NVTs was also seen. In our previous study in 2011 and 2013–2014,13,14 the number of NVT isolates, particularly in serotypes 15A, 23A, 11A, 10A, and 35B, increased from 39.7% to 75.1% (p < 0.001) among noninvasive or colonization isolates from children. Furthermore, Chiba et al. reported the increase of penicillin-resistant Streptococcus pneumoniae (PRSP) with NVTs 15A and 35B among cases of invasive pneumococcal disease (IPD) in Japanese children. 11 However, drug resistance of the NVTs in pneumococci and their genetic traits responsible for resistance have not yet been well elucidated. The purpose of the present study was to investigate the prevalence of antimicrobial drug resistance and genetic characteristics in the PNSP isolates of non-PCV13 type S. pneumoniae associated with noninvasive infections and/or colonization from children in northern Japan.
Materials and Methods
Pneumococcal strains
In our previous study, the distribution of serotypes and macrolide resistance genotypes of 2057 noninvasive isolates of S. pneumoniae from April 2013 to November 2014 was described. 14 In this study, in a PCV13 routine immunization period between July and December 2014, a total of 293 nonduplicate isolates from noninvasive infections or colonization (i.e., isolates from nonsterile specimens) were derived from outpatients of children (<16 years of age) who visited various hospitals and clinics throughout Hokkaido, the northern main island of Japan. Among them, all the isolates of non-PCV13 serotypes (231 isolates) were analyzed in the present study. The NVT isolates were assigned to 12 serotypes, including the common types 6C (16.9%), 15A (16%), 23A (13.9%), 15C (11.7%), 10A (11.3%), and 35B (9.1%), which accounted for 78.9% of all the NVT isolates. All isolates were stored in Microbank (Pro-Lab Diagnostics, Richmond Hill, Canada) at −80°C until analysis.
Serotyping and discrimination of serotypes 15A from 15F by PCR
In our previous studies, multiplex polymerase chain reactions (M-PCRs) were used to determine serotypes, including 15A, as described by Pai et al. in 2006. 15 However, thereafter, serotype 15A determined by the method 15 was revealed to contain 15F as well as 15A, which was alerted by the CDC website (www.cdc.gov/streplab/pcr.html). Between serotypes 15A and 15F, there is a difference in length with approximately 130 bp sequence in wciZ gene (GenBank accession numbers CR931663 for serotype 15A and CR931666 for serotype 15F). 16 In the present study, serotypes 15A and 15F were discriminated by PCR using newly designed primers 15bc-n1 (5′-TGGAGAGAAGGAATATGCAG-3′) and 15bc-n2 (5′-CTATTCCAAAAAATGGTATTCCCA-3′) producing different sizes of products (165 bp and approximately 290 bp for 15A and 15F, respectively, based on the earlier reference sequences). Using this method, all the isolates previously assigned to serotype 15A were confirmed to have a genetic trait of 15A.
Antimicrobial susceptibility testing
Susceptibilities to penicillins [PEN, ampicillin (AMP)], ampicillin–sulbactam (SAM), macrolides [erythromycin (ERY), clarithromycin (CLR), azithromycin (AZM)], clindamycin (CLI), cephalosporins [cefepime (FEP), cefuroxime (CXM), ceftriaxone (CRO)], carbapenems [imipenem (IPM), meropenem (MEM)], quinolones [levofloxacin (LVX), moxifloxacin (MXF), gatifloxacin (GAT)], vancomycin (VAN), tetracycline (TET), trimethoprim–sulfamethoxazole (SXT) were measured by the broth microdilution method using “Dry Plate, ‘Eiken’ HW04 (Eiken Chemical Co., Tokyo, Japan)” for all isolates. By this method, the minimal inhibitory concentration (MIC) was measured within a limited range of concentration. S. pneumoniae ATCC49619 was used as a quality control strain for the testing. Susceptibility class (susceptible [S], intermediate [I], or resistant [R]) was determined according to the Clinical and Laboratory Standards Institute (CLSI) M100-S24, 17 in which breakpoints for oral administration [S, I, R (μg/ml): ≤0.06, 0.12–1, ≥2, respectively] were applied to PEN. Because breakpoints for AMP and SAM are not provided by the CLSI, the European Committee on Antimicrobial Susceptibility Testing (EUCAST) Clinical Breakpoint Table v. 6.0, valid from January 1, 2016, 18 was used to interpret susceptibilities. Nonsusceptibility to PEN (MIC, ≥0.12 μg/ml), combined with resistance to two or more non-β-lactam antimicrobial classes, were defined as MDR.2,6
Detection of resistance genes and assignment of PBP genotypes
The prevalence of macrolide resistance genes [erm(B) and mef(A/E)] for S. pneumoniae isolates had been investigated in our previous study. 14 In the present study, a tetracycline resistance gene tet(M) was detected by PCR as described elsewhere. 19 The presence of the three unaltered pbp genes (pbp1a, pbp2x, and pbp2b) was examined by M-PCR as described previously.20,21 According to the previously published scheme,20,21 genotype of penicillin resistance (PBP genotypes) was represented as gPRSP (three pbp gene alterations) and gPISP (one or two pbp gene alterations). gPISP was expressed with the altered PBP gene(s) in a parenthesis, for example, gPISP (pbp2b).
MLST and sequence analysis of pbp1a, pbp2x, and pbp2b genes
To investigate the molecular characteristics of PNSP, we selected the representative 20 isolates of dominant serotypes, including all PRSP isolates and PISP with different resistance patterns and/or MDR [PRSP; 15A (eight isolates) and PISP; 15A, 35B, 23A, 6C (three isolates each)] for analysis by sequence types (STs) through the MLST scheme and sequences of the transpeptidase (TP) region of PBP1a, PBP2x, and PBP2b. The MLST protocol with primers for amplification and sequencing described in the S. pneumoniae MLST website (http://pubmlst.org/) was used, and the STs were also assigned at the database. The obtained STs of isolates, together with serotypes, were compared with PMEN international clones (www.sph.emory.edu/PMEN). Partial pbp1a, pbp2b, and pbp2x gene sequences encoding TP-domains (1119, 1053, and 1398 bp, respectively) were determined by PCR and direct sequencing as previously described.22,23 The PCR products were sequenced on an automated sequencer (Applied Biosystems PRISM 3130) using the Applied Biosystems Big Dye® Terminator v1.1 Cycle Sequencing Kit. The obtained sequence data were analyzed with BLAST (http://blast.ncbi.nlm.nih.gov/Blast), and alignment of sequences was performed by ClustalW program (http://clustalw.ddbj.nig.ac.jp) and Molecular Evolutionary Genetic Analysis software (MEGA5.05), comparing with those of reference strain R6 (GenBank accession numbers M90527 for pbp1a, X16367 for pbp2x, and X16022 for pbp2b).
Statistical analysis
Statistical analysis was performed by SPSS 19.0 (IBM). A two-tailed chi-square test or Fisher's accurate probability methods (for small group sizes) were used to compare proportions of antimicrobial susceptibility in each serotype. A p < 0.05 was considered statistically significant.
Nucleotide sequence accession numbers
The pbp1a, pbp2x, and pbp2b gene sequences containing TP-domain-encoding regions of 13 PNSP isolates were deposited to GenBank under accession nos. KU749446–KU749484.
Results
Antimicrobial susceptibility
The percentages of intermediate resistance (I) and resistance (R) of all isolates with non-PCV13 serotypes are summarized in Table 1. All isolates were susceptible to LVX, MXF, GAT, and VCM (data not shown). PISP and PRSP were detected in 39.4% and 3.5% of the isolates, respectively. All the serotype 15A isolates were nonsusceptible to PEN (PRSP, 21.6%; PISP, 78.4%), and PISP was found in serotypes 23A (96.9%), 6C (41.0%), 35B (33.3%), 15C (18.5%), 15B (8.3%), 11A (6.3%), and 10A (3.8%).
All tested isolates were susceptible to LVX, MXF, GAT, and VCM.
CLSI does not provide MIC breakpoints for AMP and SAM, therefore, EUCAST breakpoints 18 (AMP; S ≤ 0.5 μg/ml, R > 2 μg/ml) were used. SAM susceptibility inferred from the MIC of AMP.
MDR was defined as nonsusceptibility to PEN (MIC ≥0.12 μg/ml) combined with resistance to 2 or more antibiotic classes.2,6
AMP, ampicillin; AZM, azithromycin; CLI, clindamycin; CLR, clarithromycin; CRO, ceftriaxone; CXM, cefuroxime; ERY, erythromycin; FEP, cefepime; IPM, imipenem; MDR, multidrug resistance; MEM, meropenem; MIC, minimum inhibitory concentration; PEN, penicillin; SAM, ampicillin–sulbactam; SXT, trimethoprim–sulfamethoxazole; TET, tetracycline.
A high rate of nonsusceptible isolates to macrolides (ERY, CLR, and AZM), TET, and CLI (≥92.7%, 92.6%, and 75.3%, respectively) was observed. Of note, serotypes 15A, 23A, 15C, and 35B were 100% resistant to macrolides and TET. Among serotypes 15A and 35B, considerable isolates showed intermediate susceptibility to IPM (51.4% and 19.0%, respectively), MEM (32.4% and 23.8%, respectively), and CXM (48.6% and 23.8%, respectively). All the PNSP of serotype 35B were nonsusceptible to CXM. In addition, all the serotypes 15A and 35B were nonsusceptible to SXT. Overall, MDR was observed in 42.0% (97/231) of NVT isolates, containing dominant types 15A (n = 37) and 23A (n = 31), followed by 6C (n = 14), 35B (n = 7), and 15C (n = 5).
MLST and resistance gene genotypes in PNSP strains
Among the PNSP isolates of serotypes 15A, 35B, 23A, and 6C, representative 20 isolates were selected for further genetic analysis. In terms of MLST, 90% (18/20) of these isolates revealed to belong to PMEN clones or their single-locus variants (SLVs) (Table 2). All the serotype 15A isolates were classified into ST63, that is, PMEN clone Sweden15A-25 (international clone), and serotype 35B isolates were assigned to ST558, which is an SLV in ddl gene of Utah35B-24 clone ST377. Among the serotype 23A isolates, one isolate was assigned to ST338 (Colombia23F-26), while two isolates to ST5242, an SLV of ST338. Among serotype 6C, one isolate was assigned to ST242 (Taiwan23F-15), while remaining two isolates to ST5832.
ddl gene locus number in SLV of PMEN clones is underlined.
International clone name with clone number in the list in PMEN website (www.sph.emory.edu/PMEN) was assigned by serotype and ST of each strain.
gPISP(pbp2x+2b), SRP1988, SRP2038, SRP1987, and SRP2124; gPISP (pbp1a+2b), SRP2130; gPISP (pbp2b), SRP1921.
PMEN, pneumococcal molecular epidemiology network; PNSP, penicillin-nonsusceptible pneumococci; SLV, single-locus variant; ST
PBP gene genotypes and genes responsible for macrolides and tetracycline resistance are shown in Table 2. Serotypes 15A, 6C, and 35B with PEN-MIC of 0.5–2 μg/ml were assigned to gPRSP, while serotype 23A isolates with PEN-MIC of 0.5–1 μg/ml were assigned to gPISP (pbp2x+2b and pbp2b). Remaining serotypes 15A and 6C isolates (PEN-MIC, 0.12 μg/ml) were typed as gPISP (pbp1a+2b and pbp2x+2b, respectively). All these isolates possessed tetracycline resistance determinant tet(M), and either macrolide resistance gene erm(B) or mef(A/E). Most of isolates with erm(B) showed high-level resistance to ERY (MIC, ≥128 μg/ml).
Sequence analysis of pbp1a, pbp2x, and pbp2b genes
The representative 20 isolates were further genetically analyzed for the TP-regions encoded by pbp1a, pbp2x, and pbp2b (Figs. 1–3). In PBP1a (Fig. 1), a mutation T371S in the 370STMK373 motif was found in all the serotype 15A PRSP isolates and PISP isolates with serotypes 15A, 6C/ST242, and 35B. On the contrary, an amino acid substitution of P432T in 428SRNVP432 motif was observed in seven of eight isolates of PRSP and 25% of PISP isolates. Four consecutive substitutions, TSQY to NTGY at positions 574–577, were also found in all the PRSP isolates and 75% of PISP isolates. Amino acid substitutions in TP regions of PBP2x were T338A (17/20) and M339F (8/20) in the 337STMK340 motif, H394L (3/20) in the 394HSSN397 motif, and L546V (17/20) in the 546LKSGT550 motif (Fig. 2). In addition, a mutation of STMK337–340SAFK was found in eight PNSP (PEN; MIC of ≥0.12 μg/ml) isolates with nonsusceptibility to cephems (CXM; MIC of ≥8 μg/ml, FEP and CRO; MIC of ≥2 μg/ml), except for two isolates (Table 2 and Fig. 2). In contrast, a mutation of H394L was observed only in three isolates in serotypes 6C and 23A, which had no mutations in other two motifs. In the PBP2b (Fig. 3), all isolates possessed only one mutation T445A in 442SSNT445 motif. Other motifs had no amino acid substitutions in any isolates.
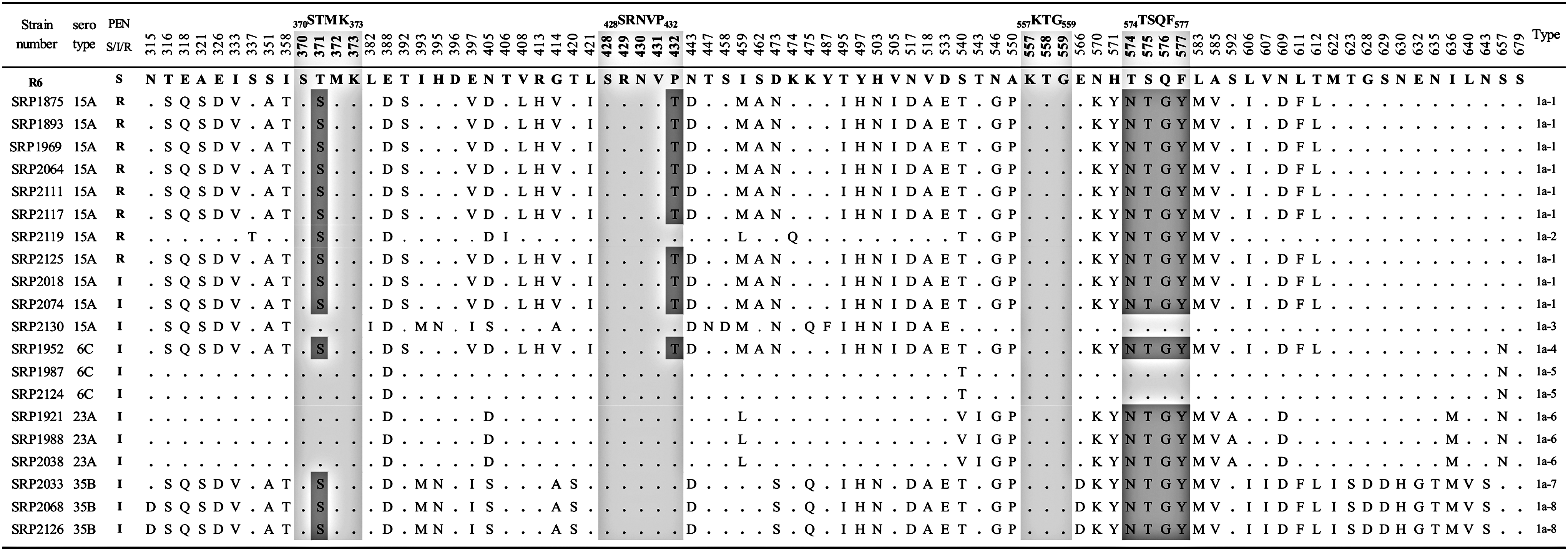
Alignment of the amino acid sequence (amino acid position) of the PBP1a transpeptidase domain region of PNSP isolates. Amino acids were compared with those of the penicillin-susceptible Streptococcus pneumoniae R6 strain in boldface. Amino acids identical to those of R6 reference strain (GenBank accession no. M90527) are represented by dots. The penicillin-binding motifs are shown in light gray. Substitutions at positions 371, 432, and 574–577 in the motif are shown in dark gray. MIC against PEN (μg/ml); ≤0.06, 0.12–1, ≥2, respectively. MIC, minimum inhibitory concentration; PEN, penicillin; PNSP, penicillin-nonsusceptible pneumococci. I, intermediate; R, resistant; S, susceptible.
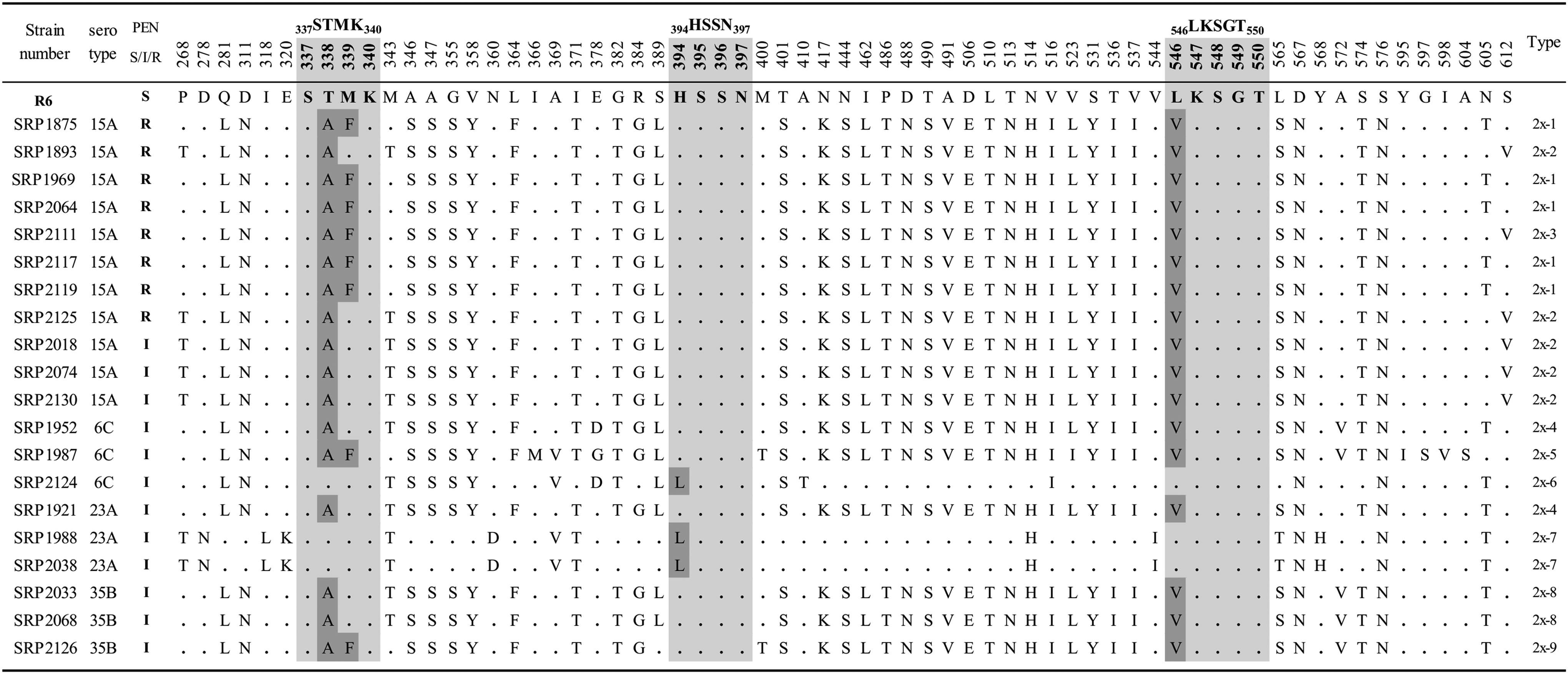
Alignment of the amino acid sequence (amino acid position) of the PBP2x transpeptidase domain region of PNSP isolates. Amino acids were compared with those of the penicillin-susceptible S. pneumoniae R6 strain in boldface. Amino acids identical to those of R6 reference strain (GenBank accession no. X16367) are represented by dots. The penicillin-binding motifs are shown in light gray. Substitutions at positions 338, 339, 394, and 546 in the motif are shown in dark gray. S, I, R: MIC against PEN (μg/ml); ≤0.06, 0.12–1, ≥2, respectively.
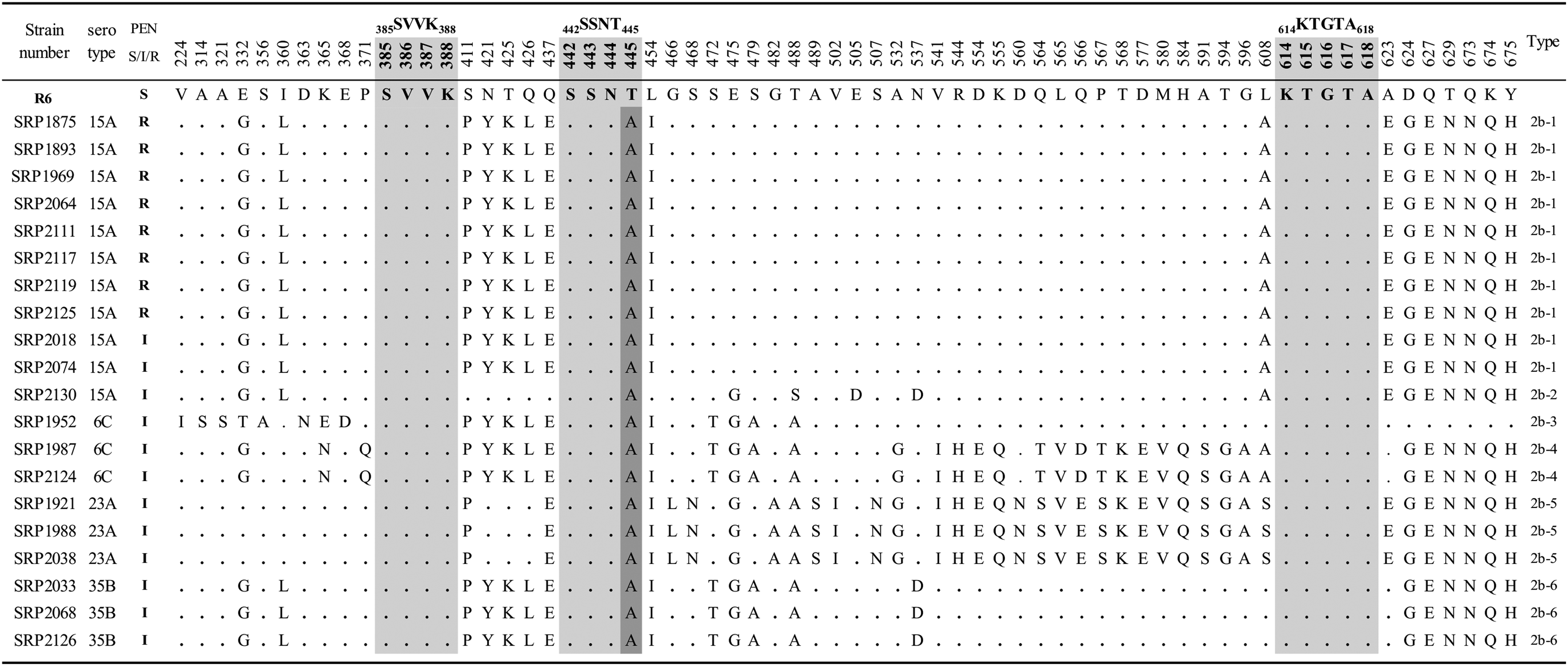
Alignment of the amino acid sequence (amino acid position) of the PBP2b transpeptidase domain region of PNSP isolates. Amino acids were compared with those of the penicillin-susceptible S. pneumoniae R6 strain in boldface. Amino acids identical to those of R6 reference strain (GenBank accession no. X16022) are represented by dots. The penicillin-binding motifs are shown in light gray. Substitutions at position 445 in the motif are shown in dark gray. S, I, R: MIC against PEN (μg/ml); ≤0.06, 0.12–1, ≥2, respectively.
Discussion
A nationwide study of Japanese pediatric patients revealed an increase of NVTs, from 2012 (51.6%) to 2014 (71.4%), and high rates of PNSP particularly in serotypes 15A and 35B (97.8% and 68.8%, respectively). 4 Similarly, in our previous study, in northern Japan, detections rates of NVTs (e.g., 15A, 23A, 11A, and 35B) among non-IPD pediatric isolates in November 2013–2014 (75.1%) were higher than those in 2011 (pre-PCV13 era) (39.7%).13,14 In the present study, all serotype 15A isolates in 2014 were found to have MDR, showing nonsusceptibility to PEN, SXT and resistance to macrolides and TET. Similarly, reduced antimicrobial susceptibility, as well as MDR, was noted also for NVTs 6C, 23A, and 35B. Notably, the high prevalence of nonsusceptible isolates to IPM and MEM was detected in only serotypes 15A and 35B. Similar to our finding, an increase of serotype 15A isolates, which are nonsusceptible to MEM, was reported in Japan in 2012–2014. 4
Tetracycline resistance is often associated with ERY resistance, 24 and their resistance genes, both tet(M) and erm(B), are often related to the MDR in pneumococci. 25 In the present study, all PRSP serotype 15A/ST63 isolates showed MDR having both tet(M) and erm(B) as seen in the surveillance study in Portugal. 26 In Japan, the most consumed antimicrobial was penicillin from nationwide surveillance at hospitals in 2010, 27 and the resistance or nonsusceptibility rate to penicillin was 42.2% in 2013 from a recent global surveillance report by WHO. 28 In contrast, macrolides account for 30% of all prescribed oral antibiotics. 29 Thus, a rapid increase in the prevalence of NVTs with antimicrobial drug resistance, especially isolates with MDR, is considered a serious public health concern in Japan. In addition, for adults ≥65 years of age, the 23-valent pneumococcal polysaccharide vaccine (PPSV23) and PCV13 are used routinely and voluntarily, respectively. However, the prevalent PNSP serotypes 6C, 15A, 23A, and 35B are not included in both PCV13 and PPSV23. Therefore, it is suggested that these PNSP serotypes, including those with MDR, may gradually spread in all age groups.
According to surveillance reports from both the United States and Canada, the increased prevalence of NVT clones in pneumococcal diseases has already been identified.6,8 In the United States, the high rate of MDR in Sweden15A-25 was documented in 2004–2005, 6 as well as a high rate of PRSP/PNSP in 35B.6,7 In Canada, the spread of MDR international clone Sweden15A-ST63 and PNSP-ST558 was observed in the post-PCV13 period in 2010. 8 ST558, an SLV in ddl gene of Utah35B-ST377, was also identified in the United Kingdom. 30 In recent studies from Japan, serotype 15A/ST63 PRSP or PISP in children4,31 and serotype 35B/ST558 with PISP in children 4 and adults 32 have also been detected. Our present study in northern Japan for non-IPD isolates also revealed the presence of ST63 and ST558 in MDR 15A and PNSP 35B isolates, respectively, suggesting clonal expansion of these drug-resistant serotypes.
A direct association between resistance to β-lactams and pbp gene alterations in pneumococcus has been described. 33 In the present study, gPRSP (pbp1a+2x+2b) isolates with PEN-MIC 0.5–2 μg/ml were identified in serotypes 15A/ST63, 6C/ST242, and 35B/ST558, while gPISP with PEN-MIC 0.12–1 μg/ml isolates was found in serotypes 6C/ST5832 and 23A (STs 338 and 5242). Our results of PBP gene genotypes in NVTs and their STs were similar to a recent report from Japan in IPD in adults. 32 Thus, it is suggested that these lineages of PNSP clones may be widely spread in both IPD and non-IPD among all age groups in Japan.
Sequence analysis of TP-domain regions in pbp genes revealed some specific mutation patterns in PNSP isolates. Regarding three motifs in TP-domain of PBP2x, only three PISP isolates harbored solely H394L mutation in HSSN motif, while all other isolates harbored T338A and L546T substitution in STMK and LKSGT motifs, respectively, as reported previously. 22 Similarly to findings in other studies,34–36 we found double substitutions T338A and M339F in STMK motif among eight PNSP (PEN; MIC of ≥0.12 μg/ml) isolates with nonsusceptibility to cephalosporins (CXM; MIC of ≥8 μg/ml, FEP and CRO; MIC of ≥2 μg/ml). It was demonstrated that change to SAFK from STMK conferred high-level resistance to cephalosporins [cefotaxime; MIC of ≥2 μg/ml] 36 and CXM (MICs of 4 and 16 μg/ml). 36 The other study 35 also revealed high-level β-lactam resistance due to four new PBP2x substitutions, such as I371T, R384G, M400T, and N605T in detail. Likewise, in the present study, SAFK in isolates highly resistant to cephems and the four substitutions in PNSP isolates were detected. In addition, other uncommon mutations L510T and T513N were observed in all PRSP isolates, in agreement with a recent study, 9 suggesting involvement of these mutations in PBP2x for resistance to penicillin. Previously reported pbp1a substitutions include T371S in the 370STMK373 motif, 37 P432T in the 428SRNVP432 motif, 38 and TSQY to NTGY at positions 574–577.23,33,39 Peculiarly, a mutation of P432T in 428SRNVP432 motif was found to confer high-level penicillin resistance (MIC of 8 μg/ml). 38 In our present study, all the PRSP and some PISP had these mutations in PBP1a at high rate. In addition, in the TP-domain of PBP2b, only T445A substitution was present in all PNSP isolates. The PBP2B mutations of TP-domain region were reported to mediate low-level resistance to penicillin. 40 Consequently, our results showed that resistance to penicillin is associated with several mutations in TP-region in pbp genes, mainly in pbp1a and pbp2x.
In conclusion, our present study elucidated the presence of isolates with MDR in several NVTs in northern Japan. Particularly, serotype 15A isolates were revealed to be Sweden15A-ST63 clone disseminating internationally. Therefore, continuous epidemiological surveillance of antimicrobial resistance in S. pneumoniae isolates, especially those with NVTs, is necessary.
Footnotes
Acknowledgment
This work was supported by a Grant-in-Aid for Scientific Research (KAKENHI) (No. 26893212 and 16K09101) from the Japan Society for Promotion of Science (JSPS).
Disclosure Statement
The authors of this article have no commercial associations that might create a conflict of interest in connection with the submitted article.