
Abstract
Thirty Salmonella enterica subsp. enterica serovar Hadar isolates of avian origin collected between 2007 and 2010 from chicken carcasses in five geographically spread abattoirs in Germany were investigated for plasmid-mediated quinolone resistance determinants. Four isolates were identified by PCR analysis and hybridization experiments to carry qnrB genes. The isolates were indistinguishable by their XbaI macrorestriction patterns and did not exhibit a mutation in the quinolone resistance-determining regions of the DNA gyrase and topoisomerase IV genes. The qnrB genes were found to be located on small plasmids of ∼2.6 kb, which mediated decreased susceptibility only to quinolones. The plasmids were assigned to the same type, pHAD28, and transformation studies into an Escherichia coli recipient strain confirmed their transferability. Sequence analysis of the complete plasmid pHAD28 revealed the presence of a qnrB19 gene. The gene was found on a novel variant of qnrB19-harboring plasmids with high similarity to plasmids pPAB19-3 from E. coli and pPAB19-4 from Salmonella sp. M9397. A presumptive recombination side was detected, suggesting that interplasmid recombination events might have played a role in the development of this plasmid variant.
Introduction
F
However, during recent years, the plasmid-mediated quinolone resistance (PMQR) determinants have been increasingly recognized, although they confer only low levels of tolerance. Nevertheless, the localization on plasmids bears the risk of a wider dissemination and selection of higher resistance levels. 4 The PMQR genes include aac(6′)-Ib-cr, a variant of an aminoglycoside acetyltransferase gene, qepA and oqxAB, encoding quinolone efflux pumps, and the qnr genes, coding for pentapeptide repeat proteins. 1 The Qnr proteins mediate decreased susceptibility by protecting type II DNA topoisomerases from binding by quinolones. 5 So far, five different qnr genes (qnrA, qnrB, qnrC, qnrD, and qnrS) have been discovered, with a number of alleles showing minor variation in nucleotide sequences. 6 In the promotor regions of some qnr genes (e.g., qnrB alleles), LexA-binding sites have been identified and the involvement of the SOS control in gene regulation has been demonstrated.7,8
Since fluoroquinolones are important antibiotics for therapy of systemic and life-threatening infections with Salmonella spp. in humans, resistance or decreased susceptibility of salmonellae to this class of antimicrobials is a matter of concern.6,9 Monitoring of quinolone resistance mechanisms, particularly with regard to the transferable PMQR determinants, is therefore required.4,5
In this study, 30 Salmonella enterica serovar Hadar strains from healthy poultry were investigated for PMQR mechanisms. Four qnrB-carrying isolates were characterized genotypically and the complete nucleotide sequence of a small plasmid was determined to gain insight into the structure of qnrB19-harboring plasmids from Salmonella Hadar.
Materials and Methods
A total of 30 Salmonella enterica subsp. enterica serovar Hadar isolates of avian origin, obtained from a monitoring study on nontyphoidal salmonellae in Germany was included in this study. 10 The isolates were collected from 2007 to 2010 from healthy chickens on five different, geographically spread abattoirs. The minimum inhibitory concentration (MIC) values of ciprofloxacin and nalidixic acid were determined by the broth macrodilution method. Performance of susceptibility testing and interpretation of results were done as per criteria of the Clinical and Laboratory Standards Institute (CLSI) document M100-S24 (breakpoints for resistance: ≥1 μg/ml ciprofloxacin and ≥32 μg/ml nalidixic acid, respectively). 11 In addition, MICs of ampicillin, cefotaxime, chloramphenicol, colistin, gentamicin, streptomycin, tetracycline, and trimethoprim/sulfamethoxazole (1:19) were determined by the agar dilution method. 12 For quality control purposes, Escherichia coli reference strain ATCC 25922 was used in all susceptibility tests.
Genomic DNA of all isolates was extracted by using the Dneasy® Blood and Tissue Kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. The occurrence of PMQR resistance genes qnrA, qnrB, qnrC, qnrD, qnrS, qepA, oqxAB, and aac(6′)-Ib-cr was investigated by PCR analyses in accordance to previously described protocols.6,13,14 A plasmid localization of the genes was confirmed by Southern blotting and hybridization with a specific gene probe labeled by the PCR DIG Probe Synthesis Kit (Roche Diagnostics GmbH, Mannheim, Germany).
Isolates with confirmed PMQR determinants were assessed by PCR and sequence analysis for mutations within the QRDR of the topoisomerase genes gyrA, gyrB, parC, and parE as described earlier. 14 The clonal relatedness of the isolates was investigated by XbaI macrorestriction analysis followed by pulsed-field gel electrophoresis as described elsewhere. 15
Plasmids of qnrB-harboring isolates were extracted with the QIAprep® Spin Miniprep Kit (Qiagen) and transferred into the recipient strain E. coli CS1562 by chemical transformation (CaCl2). Transformants were selected on Luria-Bertani (LB) agar plates containing 0.125 μg/ml ciprofloxacin. 16 Transformants were investigated for their plasmid content, resistance phenotype, and for the absence of mutations in the QRDR of topoisomerase genes. A comparison of plasmids was performed after single and double digestion with the restriction endonucleases EcoRI, ClaI, XbaI, HindIII, PstI (Fermentas, Thermo Fisher Scientific, Darmstadt, Germany), HaeIII, KpnI (New England Biolabs, Frankfurt, Germany), and XhoI (Roche, Mannheim, Germany) on the basis of their restriction patterns.
The nucleotide sequence of the complete plasmid pHAD28 was determined by primer walking (Eurofinsgenomics, Ebersberg, Germany) and sequence analysis was performed with the BLAST program blastn (http://ncbi.nlm.nih.gov/BLAST). The sequence of the qnrB19-carrying plasmid was deposited in the EMBL database under accession no KU674895.
Results and Discussion
The Salmonella Hadar isolates exhibited MIC values in the range of ≤0.0625–1 μg/ml to ciprofloxacin and 4 to >128 μg/ml to nalidixic acid (Table 1). According to CLSI interpretive criteria, all but five isolates were resistant to nalidixic acid, whereas 3 isolates were categorized as ciprofloxacin resistant, 26 isolates as intermediate susceptible and a single isolate as susceptible to ciprofloxacin. As displayed in Table 1, all isolates were classified as cefotaxime and gentamicin susceptible, while a number of isolates were resistant to ampicillin (n = 6), chloramphenicol (n = 4), and trimethoprim/sulfamethoxazole (n = 3). In contrast, all but 4 isolates were resistant to tetracycline and elevated MIC values of streptomycin (≥64 μg/ml) were seen in 27 out of 30 isolates. Notably, a single isolate exhibited an elevated MIC value of ≥8 μg/ml colistin.
Antimicrobial agents are abbreviated as follows: AMP, ampicillin; CHL, chloramphenicol; CIP, ciprofloxacin; CTX, cefotaxime; COL, colistin; GEN, gentamicin; NAL, nalidixic acid; STR, streptomycin; SXT, trimethoprim/sulfamethoxazole (1:19); TET, tetracycline.
Data represent the concentration of trimethoprim.
Isolates were screened for the plasmid-borne resistance genes qnrA, qnrB, qnrC, qnrD, qnrS, qepA, oqxAB, and for aac(6′)-Ib-cr.
MIC, minimum inhibitory concentration.
PCR-directed detection of qnrB genes revealed the presence of a 496 bp amplicon in four isolates, whereas none of the other PMQR genes [qnrA, qnrC, qnrD, qnrS, qepA, oqxAB, and aac(6′)-Ib-cr] could be detected by PCR amplification. Nucleotide sequence analysis of the qnrB amplicons confirmed the presence of an internal segment of the qnrB19 gene showing 100% nucleotide sequence identity to qnrB19 genes, for example, from E. coli and Klebsiella pneumoniae (GenBank accession no. JX298080.1 and CP007733.1, respectively).
The four qnrB19-carrying isolates exhibited MICs of 0.5 μg/ml ciprofloxacin and of 16 μg/ml nalidixic acid and in addition showed reduced susceptibility or even resistance to tetracycline and streptomycin only. None of them exhibited resistance-mediating mutations in the QRDR of the gyrA, gyrB, parC, and parE topoisomerase genes. Although collected from 2007 to 2010, macrorestriction analysis with XbaI revealed indistinguishable fragment patterns of the four isolates and, hence, they were considered as closely related. The isolates carried a single plasmid of ∼2.7 kb and hybridization studies confirmed the plasmid location of the qnrB19 gene.
Since studies reporting the presence of qnrB genes in Salmonella Hadar are rare,17,18 we decided to determine the complete nucleotide sequence of the qnrB-carrying plasmid. For this reason, the plasmids were transformed into E. coli CS1562 (MICs of 0.016 μg/ml to ciprofloxacin and of 0.5 μg/ml to nalidixic acid) and, in comparison to the recipient strain, transformants showed 64-fold higher MICs of nalidixic acid (32 μg/ml) and a 16-fold increase in MICs of ciprofloxacin to 0.25 μg/ml even though there were no mutations in the QRDR of topoisomerase genes. Similar increases in fluoroquinolone/ciprofloxacin MICs have been reported for highly susceptible E. coli recipient strains after transformation of plasmid-borne qnrS and qnrB genes, for example, from Salmonella Infantis or K. pneumoniae.14,19,20
However, while some PMQR genes in Salmonella were detected on plasmids containing extended-spectrum cephalosporin resistance genes or the blaTEM-1 β-lactamase gene, the qnrB19-carrying plasmids from Salmonella Hadar did not mediate decreased susceptibility to additional antimicrobials.14,21 Restriction analysis of single and double-digested plasmid DNA revealed indistinguishable fragment patterns, on the basis of which the four plasmids were assigned to the same type, designated pHAD28.
The complete nucleotide sequence of plasmid pHAD28 (obtained from isolate # 28) was determined by primer walking starting from the qnrB19 gene and the organization of pHAD28 is shown in Fig. 1. Sequence analysis revealed a size of 2,617 bp, slightly smaller than initially estimated.
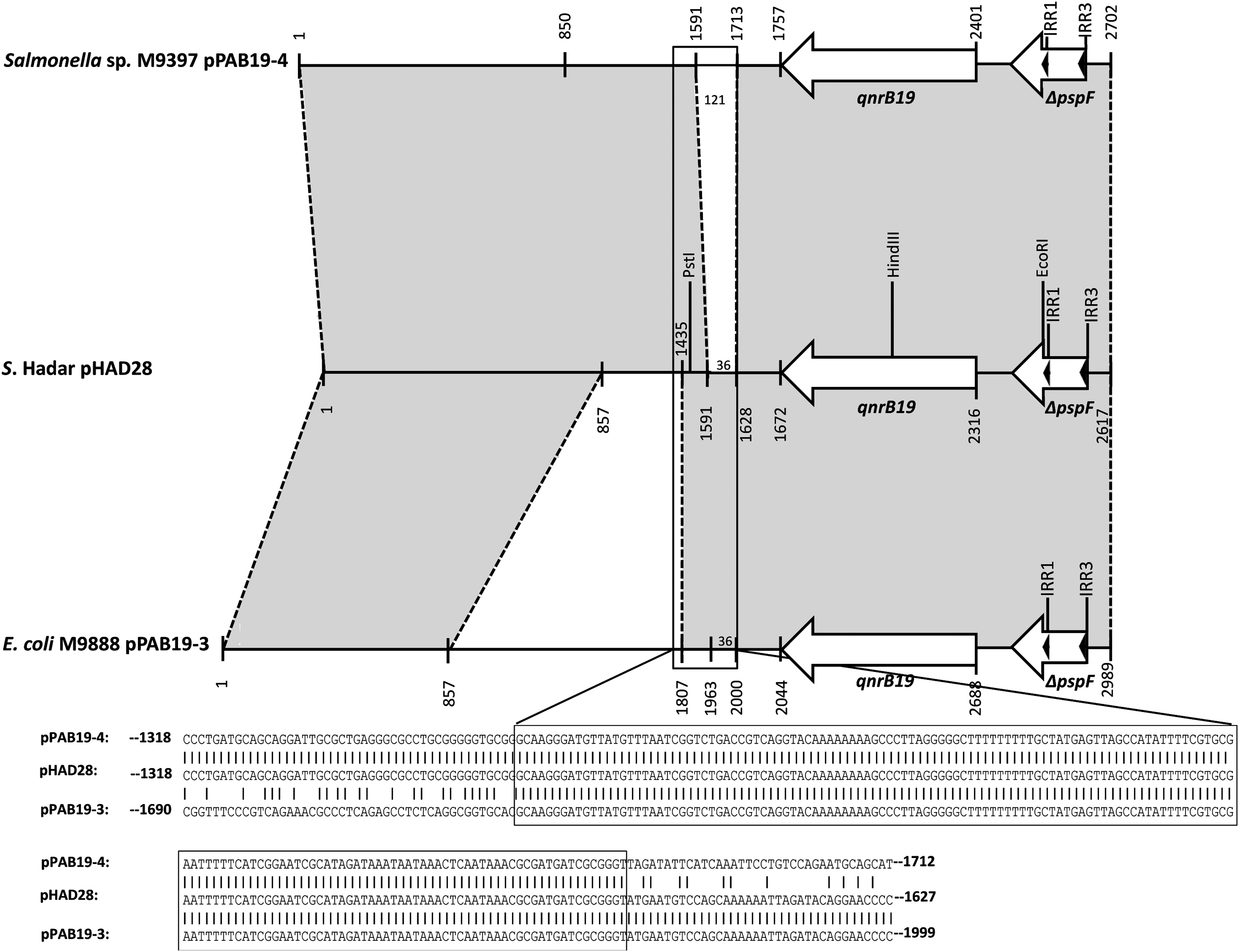
Structural organization and comparison of plasmids pPAB19-4, pHAD28, and pPAB19-3. Gray shadings represent areas of identical nucleotide sequences. A distance scale is given below the maps. The arrows indicate the qnrB19 gene and the truncated pspF reading frame. The inverted repeat-like sequences IRR1 and IRR3 are shown as dark arrowheads. The nucleotide sequence of a putative recombination site (framed in a box) and the adjacent sequences are shown below the map.
The plasmid consisted of two regions with high similarity to other qnrB19-harboring plasmids from Salmonella or E. coli. 22 The first segment (positions 1–1591) showed 100% nucleotide sequence identity to a region from Salmonella sp. M9397 plasmid pPAB19-4 (GenBank accession no. JN995611.1). Overlapping with this region, a second segment with 100% sequence identity to plasmid pPAB19-3 from E. coli M9888 (GenBank accession no. JN985534.1) was detected (positions 1435–2617). This region comprised an open reading frame for the qnrB19 protein (positions 1672–2316) and the 3′end of a truncated pspF reading frame. The pspF gene encodes a transcriptional activator of the psp operon and the truncated gene, including ISEcp1 inverted repeat-like sequences IRR1 and IRR3, was previously identified as part of the conserved genetic environment of qnrB19 plasmids. 22
Between both segments, a 157 bp overlap region was detected (positions 1435–1591) showing 100% sequence identity to plasmid pPAB19-4 (positions 1435–1591) and to positions 1807–1963 of plasmid pPAB19-3. This overlap region includes the 3′ part of a Xer site-specific recombination site, a region that was found to flank a variable DNA region (named as variable region 1) in which genetic variations of qnrB19-carrying plasmids have been described previously. 22 Sequence analysis of the parts flanking the overlap region detected 577 bp immediately upstream, that are unique to pPAB19-4, while immediately downstream, it was followed by 36 bp (positions 1592–1627) that are unique to plasmid pPAB19-3. Therefore, it might be assumed that interplasmid recombination or recombinationally active sequences might have played a role in the development of plasmid pHAD28, with the overlap region representing a putative recombination site (displayed as box in Fig. 1).
This is one of a few descriptions of qnrB19 genes in Salmonella Hadar. The gene was found to be located on a novel variant of a small qnrB19-carrying plasmid, designated pHAD28. Even though the occurrence of plasmid pHAD28 in closely related Salmonella Hadar isolates collected at various times and in different abattoirs in Germany indicated the spread of a single clone rather than a plasmid dissemination, the findings underline the need of further monitoring of PMQR determinants.
Footnotes
Acknowledgment
The authors wish to thank Vera Nöding and Inna Pahl for excellent technical assistance.
Disclosure Statement
No competing financial interests exist.