
Abstract
Staphylococcus aureus in biofilms is highly resistant to the treatment with antibiotics, to which the planktonic cells are susceptible. This is likely to be due to the biofilm creating a protective barrier that prevents antibiotics from accessing the live pathogens buried in the biofilm. S. aureus biofilms consist of an extracellular matrix comprising, but not limited to, extracellular bacterial DNA (eDNA) and poly-β-1, 6-N-acetyl-
Introduction
B
Production of biofilms by S. aureus is a significant factor involved in colonization and persistence of infections
12
as it protects the bacteria from host immune defense as well as provides a protective barrier, allowing for resistance to antimicrobial therapy.
13
A major structural component of S. aureus biofilms is represented by poly-β-1,6-N-acetyl-
Dispersin B is a 40 kDa soluble glycoside hydrolase produced by the periodontal disease-associated bacterium Actinobacillus actinomycetemcomitans.15,16 Due to its ability to specifically cleave the β-1,6-glycosidic bonds in PNAG, dispersin B efficiently degrades preformed biofilms of Staphylococcus epidermidis, but is only moderately effective in degrading S. aureus biofilms.15,17,18 Although formation of PNAG-independent biofilms by S. aureus has been reported, 19 the majority of isolates contain the ica operon cluster required for production of PNAG and biofilm formation. 20
DNase I detaches biofilms formed by S. aureus, but displays only moderate activity on S. epidermidis biofilms. 14 Dispersin B was reported to sensitize S. epidermidis, but not S. aureus, biofilms to killing by the cationic detergent, cetylpyridinium chloride (CPC), whereas DNase I also sensitized S. aureus biofilms to CPC killing, suggesting differences in the accessibility of PNAG and eDNA in the biofilms of these organisms. 18
We hypothesized that treatment of S. aureus biofilms with more than one biofilm-degrading enzyme may enhance the susceptibility to antibiotics used in staphylococcal infections. We selected tobramycin for this investigation as it is active against many gram-negative pathogens with a narrow-spectrum antimicrobial activity against gram-positive bacterial pathogens, including S. aureus. Tobramycin is commonly used as an inhalant antimicrobial to reduce the bacterial burden of the lungs of cystic fibrosis patients coinfected with S. aureus and P. aeruginosa.21,22
Our study has demonstrated that treatment of preformed S. aureus biofilms with the biofilm-degrading enzymes, DNase I or dispersin B, enhances the bactericidal activity of tobramycin. However, the antimicrobial efficacy of tobramycin was significantly reduced when preformed biofilms were treated with a combination of the two enzymes.
Materials and Methods
Production and purification of recombinant dispersin B
Escherichia coli DH5α was transformed with the plasmid, pDispersin, applying standard procedures. 23 The plasmid consisted of the expression vector pASK-IBA33plus (IBA Lifesciences) containing a synthetic gene encoding dispersin B, as described previously. 24
The E. coli strain containing pDispersin was grown overnight at 37°C in Luria-Bertani broth (Oxoid) containing 300 μg/ml ampicillin on an orbital shaker at 60 rpm. Four 1-L Erlenmeyer culture flasks, each containing 400 ml Terrific broth (Invitrogen) supplemented with 300 μg/ml ampicillin, were inoculated with 50 μl of the E. coli culture and incubated overnight at 30°C on a shaking incubator till A600nm reached ∼2. Induction of recombinant dispersin B expression was achieved by addition of 200 ng/ml anhydrotetracycline (Clontech). The flasks were placed on a shaking incubator at 37°C for 2 hr. The bacteria were harvested by centrifugation (10,000 g, 10 min, 4°C). The supernatant was discarded and the cell pellet resuspended in 16 ml extraction buffer (20 mM Tris–HCl, pH7.5, 500 mM NaCl) containing 1 mM PMSF, 2 mg/ml lysozyme, and 0.1% IGEPAL® (Sigma). The cell suspensions was sonicated three times for 10 sec at low amplitude, using a Diagenode Bioruptor Plus device (10 sec pulse-off time), followed by incubation on ice for 30 min. DNase I was added to a final concentration of 5 μg/ml, and RNase A to a final concentration of 10 μg/ml, followed by incubation at room temperature for 30 min with gentle shaking.
After removal of the cell debris by centrifugation (15,000 g, 30 min, 4°C), the supernatant was filtered through a 0.2-μm filter. The filtered supernatant was passed through a His-Select Nickel Affinity Gel (Sigma-Aldrich) column with 8 ml bed volume. The column was washed with three column volumes of extraction buffer before washing successively with extraction buffer containing 5 and 20 mM imidazole, respectively. Elution of dispersin B was achieved with two bed volumes of extraction buffer containing 100 mM imidazole. Fractions (500 μl) were collected and the A280nm of each fraction measured. Fractions containing the enzyme activity were pooled and concentrated by pervaporation by dialysis against 100 mM phosphate buffer containing 200 mM NaCl, pH 5.9. The dialysate was mixed with an equal volume of 20% glycerol in the same buffer and stored at −20°C until required for use. The protein content was estimated by assuming that a 1 mg/ml solution has an absorption value of 1.25 (http://web.expasy.org/protparam). The dispersin B activity was confirmed using 4-nitrophenyl-N-acetyl-β-
Tests for determining potential contamination of dispersin B with endonuclease or proteolytic activity
The absence of contamination of dispersin B with E. coli endonucleases was determined using a modified DNA-methyl green plate (Difco) test. The modification consisted of using the protocol for standard single radial diffusion test (SRDT), in which the wells were filled with 100 μl of dispersin B at 1.0 mg/ml, DNase I, or RNase-free EcoR1 (NEB Cat No. M0303L) at 2,000 kU/ml as a positive control and incubated at 37°C for 24 hr. The absence of a clear zone around the dispersin B well was interpreted as the absence of DNA-degrading endonucleases (figure not shown).
A second test carried out to ensure the absence of contamination of dispersin B with endonucleases was indirect ELISA, using the protocol described previously. 25 EcoR1 (NEB Cat No. R01015) or DNase I was used as the antigen. Primary antibody used was mouse polyclonal anti-DNase I (ab167580; Abcam) or mouse monoclonal anti-EcoR1 antibody (Cat No. 250334; Abbiotec). The secondary antibody used was goat anti-mouse IgG (H&L) alkaline phosphatase antibody (ab6790; abcam). The substrate used was bromo-chloro-indolyl phosphate (BCIP) and nitro blue tetrazolium (NBT).
The absence of contamination of dispersin B with proteolytic enzyme(s) was determined using proteinase K as a model protease and human serum albumin (HSA) as a model protein antigen. Dispersin B or proteinase K (V3021; Promega) was dialyzed against 0.05 M Tris buffer (pH8.6). HSA (1 mg/ml) dissolved in the same buffer was added to both tubes, which were incubated for 1 hr at 37°C. Untreated HSA was used as a control. This was followed by addition of polyclonal rabbit anti-HSA IgG (Fcγ) to the dispersin B-treated, proteinase K-digested HSA, or the undigested HSA tubes. All tubes were then incubated overnight at 4°C for the formation of an immunoprecipitate.
Bacterial growth inhibition assay
The S. aureus isolate ATCC® 29213™, a strong biofilm producer, was used in this investigation. 6 The strain was grown overnight in Nutrient broth (NB; Oxoid) at 37°C on an orbital shaker at 80 rpm. The cells were washed twice by suspending pelleted cells in cold phosphate-buffered saline (PBS; pH 7.4, NaCl 137 mM, KCl 2.7 mM/L, Na2HPO4 10 mM, KH2PO4 1.8 mM) and centrifugation at 15,000 rpm for 2 min. The cell density was adjusted to an A600nm of 0.132 and the suspension placed on ice. A tobramycin dilution series ranging from 2 to 0.0625 μg/ml was prepared in NB and dispensed in a 96-well microtiter plate (198 μl/well). Two microliters of the S. aureus suspension (0.132 A600nm) was added to each well, followed by incubation at 37°C and 80 rpm for 24 hr. The A600nm was recorded and the sample values corrected by subtraction of the averaged blank values.
DNase I and dispersin B-mediated biofilm degradation
S. aureus biofilms were grown on 96-well plates as previously described. 6 After washing the biofilms twice with PBS, 100 μl PBS containing various concentrations of either DNase I (Sigma Aldrich; D4527, constituted from a freeze-dried vial at 50 and 140 kU/ml) or dispersin B (720, 500, 100, 50, 10 μg/ml) was added to each well. The plates were incubated on an orbital shaker for 2 hr at 37°C and 50 rpm (DNase I treatment) or 80 rpm (dispersin B treatment). The remaining attached biofilms were washed once with PBS and stained for 5 min with 0.2% crystal violet solution in PBS before washing twice with PBS. The crystal violet adsorbed to the residual biofilms was solubilized in 96% methanol and the A630nm recorded on an EnSpire Multimode plate reader (PerkinElmer). The A630nm value obtained for the negative control without biofilm was subtracted from the sample values.
Effect of combinations of biofilm-degrading enzymes with tobramycin on bacterial cell viability
S. aureus ATCC 29213, a strong biofilm-forming strain, was grown overnight in NB supplemented with 1% glucose at 37°C and then diluted 1:200 in the growth medium. The bacterial suspension was dispensed in a 96-well microtiter plate (200 μl/well) and the plate incubated for 24 hr at 37°C on an orbital shaking platform at 60 rpm, followed by incubation at 37°C without shaking for an additional 24 hr. The biofilm formed on the bottom of the wells was washed twice with PBS. Solutions containing DNase I, dispersin B, and tobramycin, alone or in different combinations, were prepared as specified in Table 1 and added to the bacterial suspensions in the wells (100 μl/well) of the microtiter plate. The plate was incubated on an orbital shaker for 2.5 hr at 37°C and 60 rpm. The supernatant was aspirated from the treatment groups and remaining biofilms were scraped, suspended in 200 μl of cold PBS, and spread-plated on nutrient agar (Oxoid), followed by incubation at 37°C overnight. For validating the activity of dispersin B alone, a corresponding experiment was performed with biofilms produced by S. epidermidis.
PBS, phosphate-buffered saline.
Preparation for scanning electron microscopy
An overnight culture of S. aureus ATCC 29213 in NB was pelleted in a 1.5-ml microcentrifuge tube (12,000 rpm, 3 min). The cells were washed twice with 1 ml ice-cold PBS and adjusted with NB to 108 CFU/ml. Aluminum stubs were placed upright in a 24-well plate containing NB with 1% glucose (2 ml/well). To each well, 40 μl of the bacterial suspension was added, and the plate incubated for 24 hr at 37°C without shaking. The stubs were aseptically removed and gently washed with PBS. The stubs were placed into a new 24-well plate containing solutions of DNase I, dispersin B, and tobramycin alone or in different combinations, as specified in Table 1 (2 ml/well). After incubation at 37°C for 2.5 hr, the stubs were removed; their surface rinsed with PBS and allowed to dry at 37°C for 30 min. To each stub surface, 25 μl of 2.5% glutaraldehyde in PBS was added. After incubation for ≥3 hr at 4°C, the stubs were gently washed with water, dehydrated with 70%, 90%, and 100% ethanol, and placed into a desiccator for 3 days. The specimens were coated with 5 nm platinum and analyzed using a Zeiss Neon 40ESB Crossbeam Electron Microscope.
Results
Biofilm-degrading activity of DNase I and dispersin B
Dispersin B used in this study was found to be free of contamination with bacterial endonuclease activity, using EcoR1 or DNase I as model endonucleases, upon testing by SRDT carried out using DNA-methyl green plate, indirect ELISA, and immunoprecipitation of HSA postdigestion with dispersin B, proteinase K (data not shown), and the HSA control tube.
No clear zone was observed around the well loaded with dispersin in the SRDT, with no titer against anti-EcoR1 or anti-DNase antibody in the ELISA. An immunoprecipitate was visible in the tube containing untreated HSA and the tube containing dispersin B+HSA, but not in proteinase K-digested HSA.
In preliminary experiments, we confirmed that DNase I, applied at 140 kU/ml, efficiently degraded preformed S. aureus biofilms grown on 96-well plates, causing a reduction of 65%, as measured by staining with crystal violet, compared with the untreated control (p ≤ 0.015, data not shown). On the other hand, dispersin B, applied at 0.72 mg/ml, also degraded the S. aureus biofilm, although to a lesser extent (51%; p ≤ 0.020) in the crystal violet staining assay.
While recombinant DNase I free of RNase was purchased from Sigma Aldrich (Product No. 4716728001), specificity of dispersin B preparation was confirmed by its ability to release the p-nitrophenolate moiety from the surrogate substrate 4-nitrophenyl-N-acetyl-β-
Combined activity of biofilm-degrading enzymes with tobramycin
In a series of experiments, the effect of the biofilm-degrading enzymes and the antibiotic, tobramycin, alone and in combination, on the viability of S. aureus cells in the biofilm was investigated. In a standard broth microdilution assay, tobramycin partially suppressed the growth of S. aureus (data not shown) in the range of 0.5–1 μg/ml (A600nm < 0.1). Thus, a tobramycin concentration of 0.75 μg/ml was selected for use in the subsequent experiments for the treatment of S. aureus biofilms. Biofilms of S. aureus produced in 96-well plates were incubated with the different enzymes used in this study with or without tobramycin. Total number of viable S. aureus CFUs in each well, including the planktonic cells in the supernatant as well as the biofilm-associated cells, were then determined. Cell scraper was used to dislodge the biofilm-embedded S. aureus. In all treatment groups, the viability of S. aureus was significantly affected, although to varying degrees (Table 2). Tobramycin alone resulted in 40-fold reduction, which is comparable with the effect of dispersin B alone (no significant difference). Significantly higher (p < 0.004) efficacy was observed after treatment with DNase I alone, resulting in 1,285-fold reduction. Remarkably, the combination of DNase I with dispersin B reduced the cell viability less efficiently (p < 0.005) than DNase I alone, indicating an inhibitory effect on the activity of dispersin B for DNase I. Most active were the combinations of tobramycin with DNase I (p < 0.002) and tobramycin with dispersin B (p < 0.004), resulting in 8,780-fold and 7,500-fold reduction, respectively. Unexpectedly, the triple combination of tobramycin, DNase I, and dispersin B displayed comparably significantly less activity (129-fold reduction, p < 0.034).
N/A, not applicable.
Treatment of S. epidermidis biofilms with dispersin B resulted in 12-fold reduction (1.2 × 1012 CFU/ml) compared with the untreated control (1.24 × 1013 CFU/ml) (data not shown). Thus, dispersin B had a similar effect on the viability of S. epidermidis as on S. aureus.
Effect of biofilm-degrading enzymes and tobramycin on biofilm morphology
For studying the effect of the different treatments on the biofilm morphology, S. aureus biofilms were grown on aluminum stubs, which were then incubated with DNase I or dispersin B and tobramycin before being processed for scanning electron microscopy. The untreated control biofilms displayed the characteristic features, including sequential layering and the presence of well-developed channels that permit nutrients to flow through the biofilm (Fig. 1a). The biofilm after tobramycin (Fig. 1b) treatment was characterized by incomplete coverage of the stub surface, but still consisted of several cell layers. Compared with the control, there was a largely reduced number of cells attached to the aluminum stub with more extended areas of complete dispersal, which was observed after all treatments containing DNase I and/or dispersin B (Fig. 1c, d, respectively). The most efficient treatment for removal of the biofilm was the tobramycin-dispersin B (Fig. 1g), followed by tobramycin-DNase I, combination (Fig. 1f). This confirms total significant reduction in CFUs/ml for both treatments with tobramycin with 7,500-and 8,780-fold reductions, respectively. The triple combination treatment of tobramycin, DNase I, and dispersin B displayed low dispersal cell numbers (Fig. 1h), confirming the 129-fold reduction (Table 2).
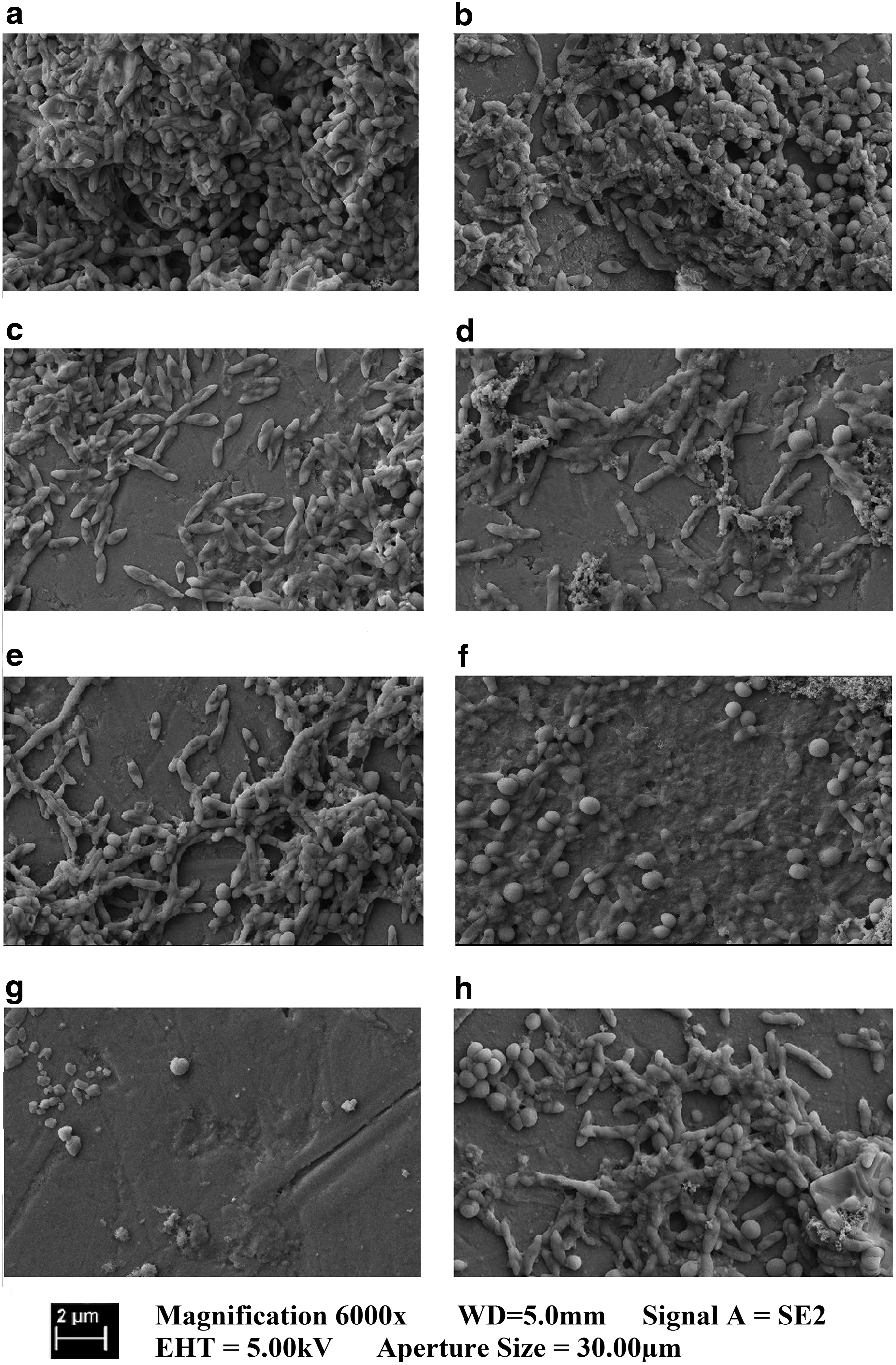
Scanning electron micrographs (magnification 6,000-fold) showing Staphylococcus aureus biofilms treated with buffer as a control (
Discussion
Previous studies have demonstrated the efficacy of DNase I in degrading the biofilms of various gram-positive and gram-negative pathogens and its ability to enhance the bactericidal activity of several antibiotics. 27 However, no investigation thus far has studied the effect of a combination of biofilm-degrading enzymes such as DNase and dispersin B in combination with an antibiotic.
The hypothesis underpinning our investigation was that the combination of two biofilm-degrading enzymes would result in an improved dispersal of biofilms and antimicrobial efficacy of antibiotics in comparison with that obtained by the treatment of biofilms with the enzymes individually. This hypothesis was not validated by the results obtained in this study involving dispersal of S. aureus biofilms by a combination of dispersin B and DNase I.
Commercially available DNase I, previously demonstrated to degrade eDNA in staphylococcal biofilms, 24 and recombinant dispersin B known to cleave PNAG were used in this investigation. 24 Tobramycin, a relatively broad-spectrum antibiotic, 28 used in the treatment of patients with chronic ear infections, 29 coinfected with S. pneumoniae and nontypeable Haemophilus influenzae, and cystic fibrosis patients frequently coinfected with S. aureus and P. aeruginosa, was used as a model antibiotic in this investigation.
Using S. aureus as a biofilm-forming pathogen and tobramycin, a clinically relevant antibiotic, our study has demonstrated that the use of multiple biofilm-degrading enzymes in combination with each other may not necessarily result in a synergistic dispersal effect as hypothesized, but may reduce the overall antimicrobial efficacy of an antibiotic. The mechanism of reduction of antimicrobial efficacy of DNase I, biofilm dispersal molecule, by dispersin B, although highly unlikely, may be due to impurities yet to be identified. However, impurities do not appear to comprise endonucleases or proteolytic enzymes. Whether this inhibition is species specific or due to steric hindrance warrants further investigations.
Regardless, it is apparent from this study that combinations of different biofilm-degrading enzymes or compounds in combination with antibiotics may require careful assessment of their ability to enhance the efficacy of different antimicrobials in vitro before their use in disinfection, covalent coating of medical devices, and/or incorporation in potential pharmaceutical excipients targeted for use in vivo.
Footnotes
Acknowledgments
Thanks are due to Curtin University for the award of an Australian Postgraduate Award to C.B.W. and the School of Biomedical Sciences for purchase of consumables for execution of this work. Financial support for Scanning Electron Microscopy by the Research Performance Index awarded to T.M. by Curtin University is also sincerely acknowledged. The authors also acknowledge financial support provided by the Hessen State Ministry of Higher Education, Research and the Arts, including a generous grant for the LOEWE research center, Insect Biotechnology and Bioresources, to A.V.
Authors' Contributions
C.B.W. and T.M. were involved in conception, approach, experimental design, data interpretation, and manuscript writing; J.R. provided the method for scanning electron microscopy (SEM) of S. aureus; N.C-.T. and C.B.W. performed SEM and assisted in interpretation; C.B.W., K.W., and D.U. performed cloning and expression of plasmid; J.G-.T. and P.C. edited the manuscript; A.G., A.V., and J.W. provided the plasmid encoding dispersin and edited the manuscript; C.B.W. and T.M. performed final editing of the manuscript; and T.M. submitted to journal.
Disclosure Statement
No competing financial interests exist.