
Abstract
Fluoroquinolone resistance in bacteria is usually associated with mutations in the topoisomerase regions. We report a novel point mutation in fluoroquinolone-resistant Escherichia coli strains. E. coli isolated from the environment in and around Mangalore, India, were examined for their antimicrobial resistance profile to 12 antibiotics and for the antibiotic resistance genes by polymerase chain reaction. Of the 67 E. coli isolated, 24 (35.8%) were sensitive to all antibiotics and 43 (64.2%) showed resistance to at least one of the 12 antibiotics used in the study. One isolate (EC10) was resistant to nine of the 12 antibiotics used. Resistance to nalidixic acid was the most common (34.32%), followed by nitrofurantoin (26.86%), tetracycline (22.38%), ampicillin (20.89%), cotrimoxazole (13.43%), ciprofloxacin (11.94%), gentamicin (10.44%), piperacillin/tazobactam (7.46%), chloramphenicol (7.46%), and cefotaxime (4.47%). Least resistance was observed for meropenem (1.49%) and none of the isolates showed resistance to imipenem. All the isolates harbored resistance genes corresponding to their antimicrobial resistance. Few quinolone-resistant isolates carried single point mutation (ser83Leu) and some had double point mutation (Ser83Leu and Asp87Asn) in gyrA. A third novel point mutation was also observed at position 50 with the change in the amino acid from tyrosine to cysteine (Tyr50Cys) in gyrA region. The study throws light on a novel point mutation in fluoroquinolone-resistant isolates. While the study helps to understand the risk and occurrence of antibiotic resistance among gram-negative bacteria from the environment, the alarming rate of antibiotic-resistant bacteria is a cause of concern in addressing infections.
Introduction
E
The emergence of resistance to quinolone or fluoroquinolone and frequent reports of resistance to several antibiotics such as tetracycline, nalidixic acid, cefotaxime, chloramphenicol, gentamicin, ampicillin, kanamycin, and trimethoprim/sulfamethoxazole have been of great concern in selecting a therapeutic agent against this organism. 3 Kumar et al., 4 noted that E. coli strains isolated from seafood from India were resistant to multiple antibiotics, thus serving as a reservoir of resistance genes in the aquatic environment. 5 Hence, it is important to study the overall prevalence of quinolone/fluoroquinolone resistance in E. coli existing in different environments to suggest interventional methods to minimize the spread of antimicrobial resistance in human pathogens.
In most of the isolates, quinolone resistance is associated with mutations in the quinolone resistance-determining region (QRDR) of the gene encoding gyrase. Hence, it is important to study these mutations and identify novel point mutations, if any, in the evolution of resistant strains. Most often, quinolone resistance is through mutation in gyrA and rarely in gyrB encoding DNA gyrase, and parC and parE encoding topoisomerase IV in E. coli.6,7 The point mutations in the gyrA gene are commonly observed at position 83, changing the amino acid from serine to leucine/valine/alanine and/or at position 87, changing aspartic acid to asparagine/valine/tyrosine/glycine. 8
The mutations involved in fluoroquinolone resistance have been extensively studied in clinical isolates E. coli in last few decades. However, the prevalence of multiple antimicrobial resistance and information on the association of new point mutations in fluoroquinolone resistance among environmental E. coli isolates are sparse and not well documented. Hence, this study is aimed at identifying the fluoroquinolone resistance due to mutations in the QRDR of gene encoding gyrase in the isolates obtained from fish and fishery products, livestock waste, piggery, and fish farm water. We also characterized the different antibiotic resistance genes in these isolates.
Materials and Methods
Bacterial strains used in the study
Sixty-seven E. coli isolates from fish and fishery products, livestock waste, piggery, and fish farm water from Mangalore were included in the study. Forty-two isolates were from fish and fishery products, 18 from livestock waste, 5 from piggery, and 2 from fish farm water. Samples were preenriched in lactose broth and streaked on eosin methylene blue (EMB) agar plate. Typical colonies from EMB plate were purified and subcultured on Luria Bertani agar (HiMedia Laboratories Pvt. Ltd., India) and subjected to a battery of biochemical tests. All strains were identified by conventional methods 9 and further confirmed by polymerase chain reaction (PCR) using primers binding to uidA encoding beta- glucuronidase enzyme. 10
Antibiotic sensitivity test
An antibiotic sensitivity test was performed for all the isolates using disk diffusion method on Mueller-Hinton agar as described by the Clinical and Laboratory Standards Institute. 11 Depending on the antibiogram profile, the isolates were examined by PCR for the presence of genes encoding resistance. The antibiotics included in the test were nalidixic acid (30 mcg), tetracycline (30 mcg), cotrimoxazole (25 mcg), ciprofloxacin (5 mcg), chloramphenicol (30 mcg), ampicillin (10 mcg), gentamicin (10 mcg), nitrofurantoin (300 mcg), imipenem (10 mcg), meropenem (10 mcg), cefotaxime (30 mcg), and piperacillin/tazobactam (100/10 mcg), and the results were interpreted as being resistant, intermediate, or sensitive according to the CLSI (2012) guidelines.
Detection of antibiotic resistance genes by PCR
Genomic DNA was extracted from the bacterial culture by Cetyl-trimethyl ammonium bromide method. 12 PCR was carried out in a 30 μl reaction mixture containing10X buffer (100 mM Tris-HCl, pH 8.3, 20 mM MgCl2, 500 mM KCl, 0.1% gelatin), 200 mM of dNTPs (dATP, dTTP, dGTP, and dCTP), 10 pmol each of forward and reverse primers, and 1.0 unit of Taq DNA polymerase enzyme. Amplification was carried out in an MJ-Research Thermo Cycler (PTC-200, Bio-Rad). An annealing temperature of 55°C for 60 sec was used for the primer uidA.
The genes conferring resistance to six categories of antimicrobials were included namely aminoglycosides, tetracyclines, quinolones, sulfonamides, phenicols, and penicillins. Several primer pairs were used for the amplification of various antibiotic resistance genes namely tetA, tetB, tetC, tetD, tetE, tetG, 13 tetL, tetM, and tetS 14 for tetracycline-resistant (TCr) isolates; sul1, sul2, and sul3 for sulfonamide; cat1, cat2, cat3, cmlA, cmlB, and floR for chloramphenicol; 13 qnrA, qnrB, and qnrS for quinolones, 15 and blaTEM and blaCTX for ampicillin- and cefotaxime-resistant isolates, respectively. 16
DNA sequencing
The products of amplification were purified by the PCR purification Kit (Qiagen) according to manufacturer's protocol and sequenced (M/s Bio serve Biotechnologies, Hyderabad, India). The generated sequences were analyzed by BLAST programme of NCBI (http://blast.ncbi.nlm.nih.gov) and submitted to the GenBank.
Selection of quinolone-/fluoroquinolone-resistant isolates
Of 67 samples analyzed, 23 (35%) and 8 (12%) isolates from various sources were resistant to nalidixic acid and ciprofloxacin, respectively. Minimum inhibitory concentration (MIC) was determined for these resistant strains using Ezy MIC™ strips (HiMedia Laboratories Pvt. Ltd., India) for nalidixic acid and ciprofloxacin. DNA was extracted from the resistant strains and PCR performed with primers binding to gyrA 17 and gyrB. 18 Amplified fragments were purified by the PCR purification Kit (Qiagen) and sequenced (M/s Bio serve Biotechnologies, Hyderabad, India). Sequences obtained were compared with the sequence of gyrA of quinolone- sensitive E. coli strain TW14359 available in GenBank (Accession No. EU898544).
Results
Identification of bacterial strain
All 67 E. coli isolates from fish and fishery products, livestock waste, piggery, and fish farm water were identified by conventional methods and confirmed by PCR. PCR amplification of uidA yielded a 146 bp product in agarose gel by electrophoresis (Fig. 1). Confirmed E. coli isolates were used for further studies.
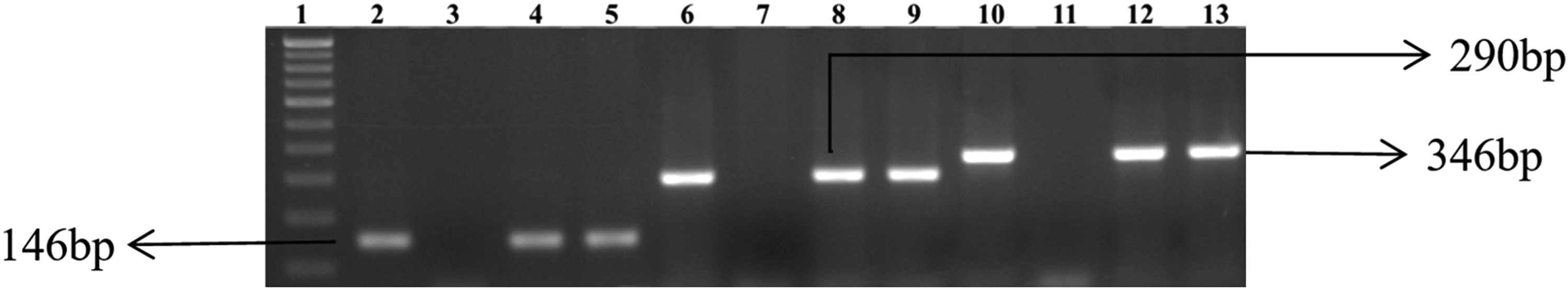
Gel electrophoresis of PCR-amplified products of Escherichia coli isolates. Lane 1: 100 bp DNA ladder; Lane 2: uidA gene, positive control; Lane 3: uidA gene, negative control; Lanes 4 and 5: positive E. coli isolates (positive uidA gene); Lane 6: positive control for gyrA gene; Lane 7: negative control for gyrA; Lane 8 and 9: gyrA gene of isolates; Lane 10: positive control for gyrB gene; Lane 11: negative control for gyrB gene; Lanes 12–13: gyrB gene of isolates. PCR, polymerase chain reaction.
Antimicrobial susceptibility tests
Results of antimicrobial susceptibility test on 67 isolates of E. coli using 12 different antibiotics belonging to 6 different groups showed 24 (35.8%) isolates to be susceptible to all the antibiotics (Table 1) and 43 (64.2%) isolates to be resistant to at least one of the antibiotics used in this study. One strain (EC10) isolated from oysters (wild environment) was resistant to nine antibiotics, which included nalidixic acid, tetracycline, cotrimoxazole, ciprofloxacin, chloramphenicol, ampicillin, gentamicin, nitrofurantoin, and cefotaxime.
Column 1: the number of different resistance pattern exhibited by the isolates; Column 2: different type of resistance pattern; Column 3: total number of isolates shows each different pattern; Column 4: percentage of isolates illustrating each different pattern.
Na, nalidixic acid; Te, tetracycline; Cot, cotrimoxazole; Cip, ciprofloxacin; C, chloramphenicol; Amp, ampicillin; Gen, gentamicin; Nit, nitrofurantoin; Mrp, Meropenem; Ctx, Cefotaxime; Pit, piperacillin/tazobactam.
All the isolates were sensitive to imipenem. Sixteen (23.8%) were resistant to at least one family of antibiotic and 13 (19.4%) to at least 2 antibiotics used in the study. Prevalence of resistance was the highest for quinolones (28, 42.2%) followed by penicillins (19, 28.2%), nitrofurantoin (18, 26.8%), tetracycline (15, 22.3%), trimethoprim/sulfamethoxazole (9, 13.4%), aminoglycoside (7, 10.4%), phenicols (5, 7.46%), and cephalosporins (3, 4.4%). Among the antibiotics tested, a lowest prevalence of resistance was recorded for carbapenems (1, 1.4%).
Detection of antimicrobial resistance genes
The genes associated with antimicrobial resistance were analyzed by PCR. Of 15 isolates showing resistance to antibiotic tetracycline, 8 harbored more than 1 tetracycline resistance gene (1-tetA and tetE; 1-tetB and tetE; 1-tetA and tetS; 2-tetA and tetL; 2-tetC and tetS; 1-tetA; tetC; and tetS), 3 carried single tetA, and 1 harbored tetB (Fig. 2). Three isolates did not carry any of the tetracycline resistance genes although they were phenotypically resistant. None of the isolates were positive for tetD, tetG, and tetM. Of the nine isolates resistant to cotrimoxazole, one each harbored sul1 and sul2, sul1 and sul3, and sul2 and sul3, and one carried only sul2. The remaining five isolates did not harbor any of the tested genes. The two strains (EC10 and EC11) carried class 1 integrons of nucleotide size 1,859 and 1,610 bp (Table 2).
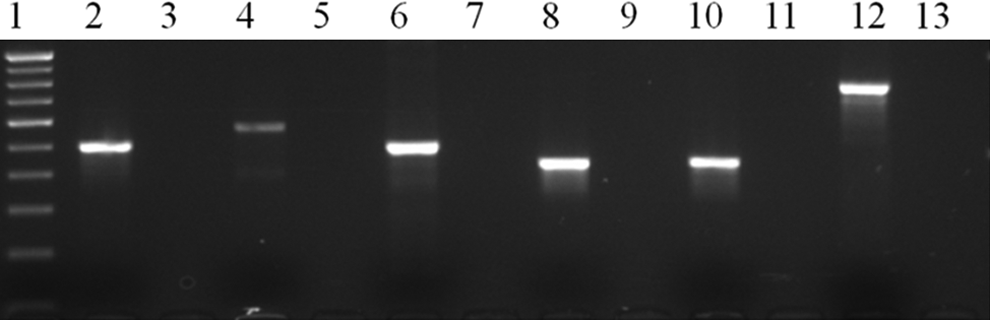
Detection of PCR-amplified products of antibiotic resistance genes: Lane 1: 100 bp DNA ladder; Lane 2: tetA (492 bp); Lane 4: tetB (571 bp); Lane 6: tetE (544 bp); Lane 8: sul1 (425 bp); Lane 10: sul2 (435 bp); Lane 12: sul3 (792 bp); Lane 14: 500 bp DNA ladder, and Lanes 3, 5, 7, 9, 11, and 13: negative controls.
Of 14 ampicillin-resistant isolates, 7 possessed blaTEM and all three cefotaxime-resistant isolates harbored blaCTX. Three of the five chloramphenicol-resistant isolates carried both cat1 and cat2 and 2 had cmlA (Fig. 3). None tested positive for cat3, cmlB, and floR. Four of the 23 nalidixic acid-resistant isolates carried qnrA and qnrB, while three harbored qnrS.
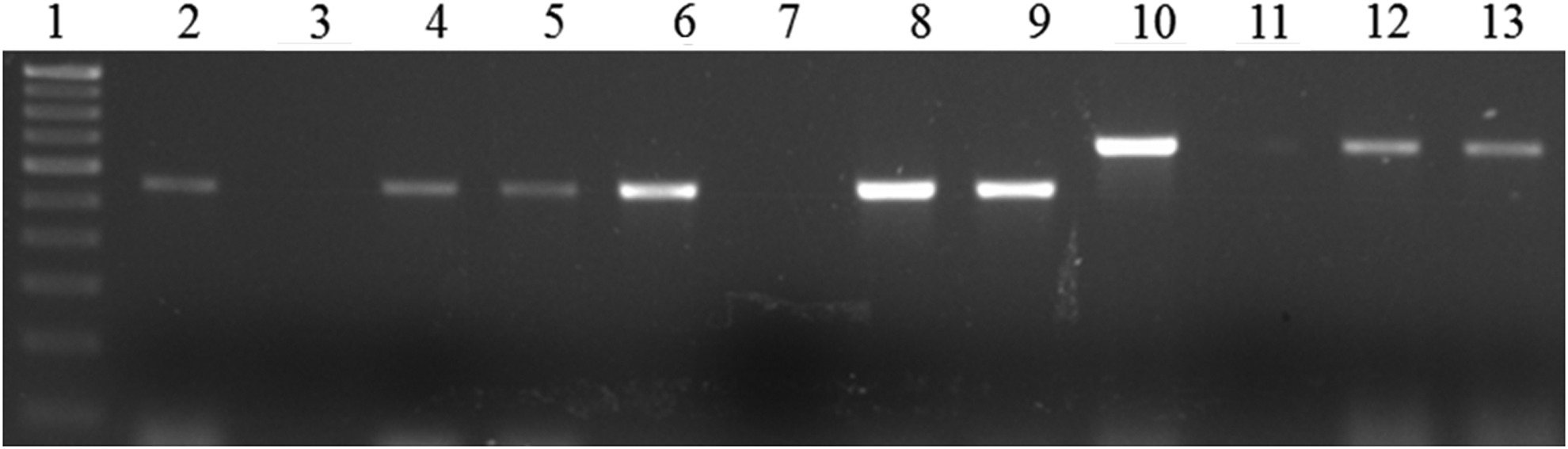
Gel-electrophoresis of PCR-amplified products of chloramphenicol-resistant isolates. Lane 1: 100 bp DNA ladder; Lane 2: cat1 gene, positive control; Lane 3: cat1 gene, negative control; Lanes 4–5: positive isolates (positive cat1 gene); Lane 6: positive control for cat2 gene; Lane 7: negative control for cat2 gene; Lanes 8–9: positive isolates (positive cat2 gene); Lane 10: positive control for cmlA gene; Lane 11: negative control for cmlA gene; Lanes 12–13: positive isolates (positive cmlA gene).
Analysis of nucleotide sequences
The PCR products of antibiotic resistance genes from all the representative isolates of E. coli were purified and sequenced (M/s Bioserve Biotechnologies, Hyderabad, India) followed by bioinformatics analysis. Thus, the novel sequences were submitted to the GenBank and accession numbers were obtained (Table 2).
Detection of mutations in QRDR regions
Isolates resistant to nalidixic acid showed an MIC of more than 256 μg/ml, whereas that of sensitive isolates ranged between 1.5 and 3.0 μg/ml. MIC of isolates resistant to ciprofloxacin ranged between 0.032 and 4.00 μg/ml, while that of sensitive isolates ranged from 0.012 to 0.032 μg/ml. Strains resistant to both nalidixic acid and ciprofloxacin were screened for mutation using primers for the amplification of gyrA and gyrB (Table 3). Nucleotide sequence results of the gyrA of the isolates EC02 and EC23 revealed two transition points, one from C → T causing the replacement of the amino acid serine to leucine at position 83 (ser-83-leu) and the other from G → A resulting in the replacement of aspartic acid to asparagine at position 87 (asp-87-asn). The differences in the nucleotide sequence at single point infer amino acid substitution in the protein synthesized and could explain their resistance to antibiotics.
Results of quinolone resistant isolates (“+” indicates occurence of mutation and presence of qnr genes; “-” indicates no mutation and absence of respective qnr genes).
NA, nalidixic acid; CIP, ciprofloxacin.
A novel transition mutation was also noticed outside the QRDR region of gyrase. Isolate EC 23 depicted a transition from A → G at nucleotide 149, which changed the amino acid at position 50 from tyrosine to cysteine. It showed an MIC of 4.00 μg/ml for ciprofloxacin, whereas the isolate (EC02) with two common mutations (ser-83-leu and asp-87-asn) showed 1.5 μg/ml.
Discussion
Antibiotic resistance in bacteria has become a global issue since it is becoming increasingly difficult to address it both in the environment and in hospitals. The source of resistance is directly linked to its indiscriminate use in various food-producing sectors leading to the emergence of resistant population as a result of selective pressure exerted by exposure to antibiotics. 19 Since E. coli is associated with gut of humans and warm-blooded animals, and ubiquitous, antibiotic resistance in them may reflect the exposure to antibiotics as well as spread from them to other gram-negative organisms in their natural habitat.
Of 12 antibiotics tested in this study, the isolates showed resistance to all except imipenem. Tetracyclines are among the therapeutic agents that have been traditionally used extensively not only as an infection control agent in human and veterinary medicines but also as a growth promoter in animal husbandry and agriculture. Their wide application has led to the equally fast spread of tetracycline resistance that is mediated by four mechanisms: efflux pump, ribosomal protection, enzymatic inactivation, and target modification. 20 It was observed from the results of this study that on an average, 23% of E. coli exhibited resistance to tetracycline.
There have been studies showing that TCr isolates harbor genes responsible for resistance mediated by different mechanisms such as tetA, B, C, D, E, G, H, and L, which code for efflux proteins, tetM, O, S, and W coding for ribosomal protection protein (RPP), tetX coding for a protein that causes enzymatic inactivation, and tetU whose function is still unknown. 21 In this study, eight efflux protein coding genes (tetA, B, C, D, E, G, and L) and two RPP coding genes (tetM and S) were examined in the 15 TCr isolates. Eight isolates harbored more than one tetracycline resistance gene, three possessed single tetA, and one isolate carried tetB. It was interesting that three isolates did not carry any of the tet genes, although they were phenotypically resistant, suggesting the possibility of some other mechanism of resistance in them.
Cotrimoxazole, a sulfa drug comprising one part of trimethoprim and five parts of sulfamethoxazole, generally designated trimethoprim/sulfamethoxazole, and used to treat a broad spectrum of bacterial infections, is used extensively in both veterinary and human medicine. Resistance to cotrimoxazole emerged soon after its initiation into clinical use and has since been on the increase. 22 Sul1, Sul2, and Sul3 are the only three alternative sulfonamide dihydropteroate synthase genes in gram-negative bacteria described to date.
In this study, 9/67 (13.43%) E. coli isolates showed phenotypic resistance. One each harbored sul1 and sul2, sul1 and sul3, sul2 and sul3, and sul2, respectively. Five isolates had none of the genes tested, suggesting the possibility of some other mechanism of resistance such as overexpression of efflux pumps in those isolates. The two strains EC10 and EC11 showed positive for class1 integrons by yielding amplicons of 1,859 and 1,610 bp, respectively. These integrons contained a dihydrofolate reductase gene (dhfrA12 in strain EC10 and dhfrA17 in EC11) and aminoglycoside adenyltransferase (aadA5 in EC11 and aadA2 in EC10) genes.
Chloramphenicol resistance in E. coli has been reported to be associated with a decreased passive accumulation of antibiotic regulated by the multiple antibiotic resistance locus or by an inactivation of the drug by an enzyme acetyltransferase. 23 The enzyme chloramphenicol acetyltransferase transforms chloramphenicol to 3-acetylchloramphenicol by converting the acetyl group of acetyl CoA in chloramphenicol to a primary (C-3) hydroxyl group, which restricts its binding to the bacterial ribosome. 23 Of 67 E. coli isolates studied, 5 (7.46%) showed phenotypic resistance to chloramphenicol. While three of them harbored type1 and type 2 cat, two isolates harbored cmlA. It was interesting that none of the isolates were positive for type 3 cat, cmlB, and floR.
Nitrofurans carry one or more nitro groups on a nitro aromatic or the nitro heterocyclic backbone and exhibit a broad antimicrobial activity against many bacterial infections, but its specific mode of action is complex. 24 Resistance to nitrofurantoin could be due to inhibition of nitrofurantoin reductase. In this study of 67 E. coli isolates, 18 (26.86%) were resistant to nitrofurantoin. Isolates were also examined for resistance to beta-lactam group of antibiotics such as ampicillin, cefotaxime, imipenem, meropenem, and piperacillin/tazobactam.
Resistance was highest to ampicillin (20.89%) followed by piperacillin/tazobactam (7.46%), cefotaxime (4.47%), and meropenem (1.5%). This group of antibiotics posses a unique structural feature, the beta lactam ring, which inhibits bacteria by inactivating enzymes involved in cell wall synthesis. Carbapenems are one of the effective antibiotics and serve as a last resort for infectious diseases caused by gram-negative bacteria such as E. coli and Klebsiella pneumoniae. Shahcheraghi et al., 25 found that all E. coli in their study were susceptible to carbapenem. Likewise, in this study, 98.5% and 100% of the E. coli isolates showed susceptibility to meropenem and imipenem, respectively, suggesting that carbapenems could be the antibiotic of choice to manage E. coli resistant to other common antibiotics.
Gentamicin, a member of the aminoglycoside group, is recognized as a potent protein synthesis inhibitor and has increasingly found application against many bacteria showing resistance to fluoroquinolones and beta-lactam antibiotics. Of 67 isolates in this study, 54 (80.59%) were susceptible to gentamicin. However, resistance in 13 isolates to a potent antibiotic such as gentamicin is a cause of concern.
Quinolones and fluoroquinolones are synthetic bactericidal antimicrobial agents with a broad-spectrum activity, being highly effective against both gram-negative and gram-positive bacteria, thus playing a major role in the treatment of serious bacterial infections. They bind to bacterial topoisomerase II (DNA gyrase) or/and topoisomerase IV and stop DNA replication, 26 and cause chromosomal breaks that lead to rapid cell death. 27 This is the primary action of quinolones that kill the bacterial cells and are thus grouped as strong bactericidal agents. However, their uncontrolled use in the field of medicine and veterinary application has resulted in the development of resistance.
Nalidixic acid is a first-generation quinolone drug and is the first synthetic quinolone. Of 67 E. coli isolated in the study, 23 (34.32%) were resistant to nalidixic acid and 8 (11.94%) to ciprofloxacin, a second-generation fluoroquinolone. 28 However, mutation in genes encoding either DNA gyrase or topoisomerase IV enzyme leads to resistance against these quinolone compounds. 29 Perichon et al. 30 noticed that mutation in the gyrA subunit of DNA gyrase or the parC subunit of topoisomerase IV occurs more frequently than gyrB and parE subunits of DNA gyrase and topoisomerase IV, respectively. In this study, a double point mutation in gyrA of quinolone-resistant E. coli isolated from the environmental sample was observed, which is otherwise common in most of the bacterial isolates from medical samples. No mutation in gyrB sequence was observed in any of the isolates tested.
Mutations at different points in the gyrase gene of E. coli resulting in resistance to quinolones were grouped in a region between nucleotides 199 (Ala67) and 318 (Gln106), denoted as QRDR. 31 Earlier reports on DNA sequence analysis revealed that most of the mutations are in the first half of the gyrA sequence in the QRDR. 32 Even in this study, mutation was observed in gyrA sequence of strain EC02 and EC23 (Table 3) at position 248, resulting in the change in amino acid at the position 83 from serine to leucine. It has been reported that a point mutation in the sequence at the 83rd position can induce amino acid change from serine to leucine/tryptophan/proline/alanine/threonine, but replacement with leucine or tryptophan presents a marked high resistance and selective advantage. 32
After a first step mutation ser-83-leu, an additional mutation at position 87 was also observed and reported in clinical isolates of E. coli associated with high-level resistance to quinolones. 33 In our results too, a double mutation in the gyrA region was observed in an E. coli isolate obtained from the environment. Several mutations at codons 67, 81, 82, 83, 84, 87, and 106 of gyrA that confers resistance have been reported, 34 but double mutation at codon 83 and 87 is most commonly observed in the clinical isolates.
They present a higher MIC for quinolones as has been evidenced in this study. Nalidixic acid-resistant strains EC02 and EC23 showed an MIC of >256 μg/ml, while the MIC of nalidixic acid-sensitive strains ranged between 1.5 and 3.0 μg/ml. The ciprofloxacin-sensitive isolates showed an MIC ranging from 0.012 to 0.032 μg/ml, whereas resistant strains showed an MIC between 0.032 μg/ml (EC02) and 4.0 μg/ml (EC23).
Vila et al., 35 observed mutation ser-83-leu at codon 83, which was constant, whereas at codon 87, the amino acid residues were variable, that is, aspartic acid to tyrosine or asparagine and the same was corroborated by Conrad et al. 33 In respect of changes at position 87 in this study, we noticed transition G → A at nucleotide 259, which ensured change in amino acid from aspartic acid to asparagine (asp-87-asn), resulting in loss of negative charges. Our study is in agreement with earlier studies, which suggest positions 83 and 87 in QRDR of gyrA to be the hot spots for mutation in E. coli where point mutation reduces quinolone–enzyme interaction and acquires resistance. 36
Earlier reports support the argument that resistance to quinolones arises within the QRDR of gyrA, but Friedman et al. 37 observed mutation at position 51, a region outside the QRDR, and reported it as a novel point mutation that decreases susceptibility to fluoroquinolones. In this study, such a mutation was observed in gyrA sequence of strain EC23, in which a nucleotide at 149 shifted from adenosine to guanine (A → G), which caused an amino acid change at codon 50 from tyrosine to cysteine. Mutation at codon 50 in gyrA nucleotide sequence was previously reported by Jaktaji and Mohiti, 38 but the shift in amino acid was from tyrosine to phenylalanine.
The isolate (EC23) that had two common (ser-83-leu and asp-87-asn) and an additional point mutation showed a reduced susceptibility to ciprofloxacin with an MIC of 4.0 mcg/ml and all other isolates (EC02) that reported two mutations showed an MIC of 1.5 mcg/ml. This suggests that the new point mutation in the isolate EC23 has little, but a noticeable effect causing a difference in the ciprofloxacin MIC value. However, the results are only indicative and further studies are required to confirm the involvement of novel mutations in conferring resistance. In addition, in our study, some silent mutations were also noticed at a few points in the gyrA sequence, but this resulted in no substitution of amino acid residue at that particular point. This suggests that not all mutations confer resistance to quinolones/fluoroquinolones.
Quinolone resistance by mutation in QRDR region resulting in overexpression of efflux pumps, has also been reported by Martinez et al. 39 They identified the plasmid-mediated quinolone resistance determinant qnr protein (qnrA1) on a conjugative plasmid isolated from ciprofloxacin-resistant K. pneumonia. 40 Quinolone resistance (qnr) proteins belong to the pentapeptide repeat protein family, 41 which protect the bacterial topoisomerases against the quinolone/fluoroquinolone compounds. Till date, five qnr families of proteins, that is, qnrA, qnrB, qnrS, qnrC, and qnrD have been identified and characterized.42,43 In this study, nalidixic acid- and ciprofloxacin-resistant isolates carried qnrA, qnrB, and qnrS, which impart the acquisition of quinolone resistance through plasmid-mediated determinants.
Conclusion
This study reveals an alarming rate of resistance that is prevalent in E. coli to the different antimicrobials. Furthermore, the detection of resistant determinants confirm that these isolates can be carriers of resistance genes, which could be easily disseminated through horizontal gene transfer. Point mutations observed in QRDR of gyrase of quinolone-resistant E. coli isolates confirm target-based resistance to quinolones. The presence of plasmid-mediated quinolone resistance genes in some of the E. coli isolates indicates the possibility of multiple mechanisms of resistance to a particular antibiotic.
Footnotes
Acknowledgment
Financial support received by the corresponding author from the Indian Council of Medical Research (Grant No. AMR/37/2011-ECD-I) toward this study and the research fellowship to the first author are gratefully acknowledged.
Disclosure Statement
No competing financial interests exist.