
Abstract
Aim:
The aim of this study was to characterize two carbapenem-resistant Klebsiella pneumoniae isolates recovered from urine samples in a patient hospitalized at Annaba University hospital (Algeria) in 2014.
Results:
Two K. pneumoniae isolates were studied because they proved resistant to almost all antibiotics tested with a high level resistance to ertapenem (minimum inhibitory concentration = 32 mg/L). The results of modified Hodge test and combined disk test (ROSCO Diagnostica, Taastrup, Denmark) were positive. The two isolates harbored the blaNDM-1 gene and one was also positive for blaCTX-M-15. Screening of aminoglycoside-modifying enzymes and plasmid-mediated quinolone resistance contents detected aac(6′)-Ib-cr, aac(3′)-II, qnrB2, and oqxAB in both isolates. Multilocus sequence typing demonstrated that the two isolates belonged to sequence type 147. However, repetitive sequence-based PCR and pulsed-field gel electrophoresis showed that they were not clonally related. The blaNDM-1 gene and all other resistant genes were contained on an IncR plasmid of c.a. 85 kb.
Conclusions:
This study comprises the first identification of NDM-1-producing K. pneumoniae in Algeria. We thus confirm the concerning worldwide dissemination of this carbapenemase that involves the emergence of the IncR plasmid and the success of the ST147 clonal complex harboring it.
Introduction
T
In Algeria, few data concerning the emergence and diffusion of this resistance mechanism are available, although several reports have described the presence of NDM-1 in Acinetobacter baumannii in Algerian hospitals.7–9 Recently, autochthonous infections because of NDM-5-producing Escherichia coli isolates have also been described in Algeria. 10 In this study, we report the first detection of two blaNDM-1-producing K. pneumoniae clinical isolates in Algeria.
Materials and Methods
Case selection and clinical data
Data collection and report of the case were approved by the Ibn Roch University Hospital ethical committee (Annaba, Algeria).
In November 2013, a 32-year-old patient was hospitalized in Annaba University hospital (Algeria). He had Guillain-Barré syndrome characterized by febrile paralysis of the peripheral nervous system caused by respiratory complications. In his medical history, we noted two prior hospitalizations in Algeria (Hospital of El Oued, southeast near the Tunisian border) and in Tunisia (Hospital Soukra, Tunis) during the past year. During the hospitalization in Annaba hospital, a bronchoalveolar lavage sample was obtained and A. baumannii and Pseudomonas aeruginosa were isolated. In December 2013 the patient was readmitted to the intensive care unit in Annaba University hospital. The clinical status of the patient worsened necessitating intubation, ventilation, and tracheotomy. He remained bedridden for 2 months. He was treated with different antibiotics: piperacillin–tazobactam, colistin sulfate, and imipenem. In February 2014, a urinary tract infection was diagnosed and the urine samples examined the same day revealed the presence of two K. pneumoniae isolates (with two different aspects on agar medium) resistant to carbapenems from the same sample. He then received three daily doses of colimycin 2,000,000 UI for 3 weeks. No stool carriage assay of carbapenemase-producing strains was performed. Patient follow-up showed that he was still alive at that point after discharge.
Identification and susceptibility testing
Bacterial identification was determined using the Vitek® MS system (bioMérieux, Marcy l'Etoile, France). Susceptibility to antimicrobial agents was tested by the disk diffusion method (Bio-Rad, Marnes La Coquette, France) on Mueller–Hinton agar according to recommendations of EUCAST-SFM (www.sfm-microbiologie.org). The following antibiotics were tested: ampicillin, amoxicillin–clavulanic acid, ticarcillin, ticarcillin–clavulanic acid, piperacillin, piperacillin–tazobactam, cefalexin, cefixime, cefotaxime, ceftazidime, cefepime, cefoxitin, aztreonam, amikacin, netilmicin, gentamicin, tobramycin, nalidixic acid, ofloxacin, ciprofloxacin, cotrimoxazole, tetracycline, chloramphenicol, tigecycline, and fosfomycin. In addition, minimum inhibitory concentration (MIC) of carbapenems (ertapenem, imipenem, meropenem, and doripenem) was determined by the E-test method (bioMérieux). MIC of colistin was determined using microbroth dilution (Umic®; Biocentric, France). The MICs were interpreted as specified by the EUCAST-SFM criteria. The nonsusceptibility to the carbapenems and colistin was assessed on the following MICs: ertapenem >0.5 mg/L, imipenem >2 mg/L, meropenem >2 mg/L, doripenem >1 mg/L, and colistin >2 mg/L.
Phenotypic detection of carbapenemase
Detection of carbapenemase producers was investigated in the two isolates using a modified Hodge test 11 and KPC, MBL, and OXA-48 Confirm® kit (Rosco Diagnostica, Taastrup, Denmark) following manufacturer's recommendations. 12
Characterization of β-lactamase-encoding genes
Plasmid or chromosomal DNA was extracted from the two isolates using the EZ1 DNA Tissue kit on the BioRobot EZ1 extraction platform (Qiagen, Courtaboeuf, France). Genes encoding the most clinically prevalent carbapenemases (blaKPC, blaOXA-48-like, blaVIM, blaIMP, and blaNDM), extended-spectrum β-lactamases (ESBLs; blaTEM, blaSHV, and blaCTX-M), and penicillinases (blaTEM and blaSHV) were detected by PCR using specific primers and confirmed by sequencing the PCR products, as described previously.13–16 A multiplex PCR was used for the detection of the six plasmid-carried blaAmpC genes. 17 For each PCR tested, we used positive controls from our own collection.
Characterization of aminoglycoside-modifying enzymes and plasmid-mediated quinolone resistance
The presence of genes encoding the aminoglycoside-modifying enzymes [aac(3)-I, aac(3)-II, aac(3)-IV, aac(6′)-Ib, ant(2"), aph(3′)-Ia, and aph(3′)-VI], the 16S rRNA methyltransferases (armA, npmA, rmtA, rmtB, rmtC, rmtD, rmtE, and rmtF), and the plasmid-mediated quinolone resistance [qnrA, qnrB, qnrS, qepA, oqxAB, and aac(6′)-Ib-cr] was studied as previously described.18–22 For each PCR tested, we used positive controls from our own collection.
Plasmid characterization
Plasmid DNA extraction of K. pneumoniae isolates was performed by the alkaline lysis method using Qiagen Midiprep kit® following the manufacturer's protocol.
Plasmids were transformed into recipient E. coli TOP10-competent cells. Transformants were selected on Mueller–Hinton agar with meropenem 0.5 μg/ml, and surviving colonies were identified with the Vitek MS system. The resistance phenotype and gene content of transformants were assessed by the disk diffusion method, E-test, and PCR and compared with those of parental cells.
Plasmids from the studied strains were assigned to incompatibility groups by plasmid relaxase gene typing (PRaseT) performed on total DNA using previously described primers and conditions. 23 This method includes six different multiplex PCRs and a simplex PCR using primers targeting the most frequently encountered relaxase gene in Enterobacteriaceae. Nonrelaxase IncR replicon was screened for by an additional PCR assay. 24
Analysis of clonality
The genetic relationship between the two carbapenem-resistant isolates was evaluated by DiversiLab® system (bioMérieux), following the manufacturer's instructions. This semiautomated fingerprinting tool is based on repetitive sequence-based PCR (rep-PCR). Isolates with identical strain patterns were considered indistinguishable if the similarity percentage was ≥95%.
Multilocus sequence typing (MLST) analysis was performed using the Pasteur Institute's MLST scheme (bigsdb.web.pasteur.fr). Seven housekeeping genes (gapA, infB, mdh, pgi, phoE, rpoB, and tonB) were amplified according to this protocol.
A macrorestriction analysis of chromosomal DNA using XbaI enzyme was performed according to a previous published procedure and analyzed with GelCompar II software (Applied Maths, Sint-Martens-Latem, Belgium) as previously described. 25 Isolates were considered to be within a cluster if the coefficient of similarity was ≥80%. 26
Results
The main characteristics of the two K. pneumoniae isolates are shown in Table 1. The isolates were resistant to all β-lactams tested except aztreonam for both isolates and imipenem (MIC = 2 μg/ml) for the isolate 19M3. They also displayed a high level of resistance to aminoglycosides and fluoroquinolones. They presented susceptibility only to fosfomycin, tigecycline, and colistin (MIC = 0.25 μg/ml). Carbapenemase activity was observed using the modified Hodge test and was significantly inhibited by dipicolinic acid. PCR detection confirmed that the two K. pneumoniae isolates harbored the blaNDM gene. Sequencing analysis showed that the isolates carried β-lactamase NDM-1. Neither ESBL-encoding nor ampC genes were detected in 19M3. 19V harbored the blaCTX-M-15 gene. The two isolates also presented qnrB2, oqxAB, aac(6′)-Ib-cr, and aac(3′)-II genes that confer resistance to quinolone and aminoglycosides. No methyltransferase gene was present. Rep-PCR and pulsed-field gel electrophoresis (PFGE) revealed that the two isolates were nonclonal (Fig. 1). However, MLST classified the two isolates into the same sequence type clone ST147.
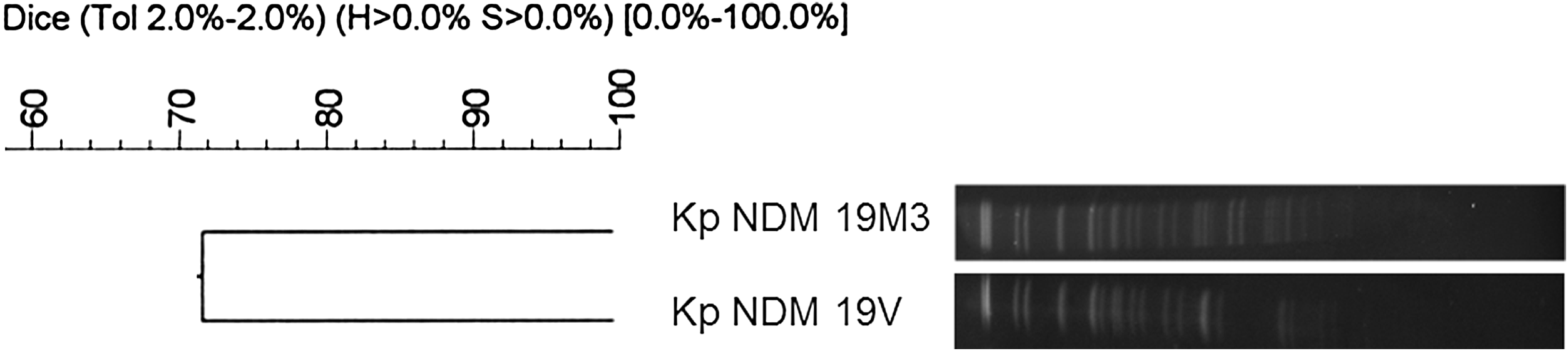
Clonality analysis (PFGE) of clinical NDM-1-producing Klebsiella pneumoniae isolated in a patient hospitalized at Annaba University Hospital (Algeria). Isolates with identical strain patterns were considered clonal if the similarity percentage was ≥80%. PFGE, pulsed-field gel electrophoresis.
AMC, amoxicillin–clavulanic acid; AMK, amikacin; AMP, ampicillin; ATM, aztreonam; CAZ, ceftazidime; CFM, cefixime; CFX, cefalexin; CHL, chloramphenicol; CIP, ciprofloxacin; CTX, cefotaxime; DOR, doripenem; ETP, ertapenem, FEP, cefepime; FOX, cefoxitin; GEN, gentamicin; IPM, imipenem; MIC, minimum inhibitory concentration; MEM, meropenem; NAL, nalidixic acid; NET, netilmicine; NIT, nitrofurantoine; OFX, ofloxacin; PIP, piperacillin; PMQR, plasmid-mediated quinolone resistance; SXT, cotrimoxazole; TET, tetracycline; TIC, ticarcillin; TIM, ticarcillin/clavulanic acid; TOB, tobramycin.
For both strains, only one plasmid of ∼85 kb was isolated after plasmid extraction. To study the transferability of the resistance phenotype, electroporation experiments were performed. The assays to transfer plasmid isolated from 19M3 were unsuccessful, despite repeated attempts, but the plasmid isolated from 19V was successfully transferred into E. coli. A transformant was obtained exhibiting resistance to all β-lactams except aztreonam, including carbapenems (MICs of imipenem, ertapenem, meropenem, and doripenem were 6, 32, 4, and 8 μg/ml, respectively). The E. coli transformant acquired a c.a. 85 kb plasmid. PCR amplification confirmed that the plasmid present in transformant harbored the blaNDM-1 gene together with blaCTX-M-15, qnrB, oqxAB, aac(6′)-Ib-cr, and aac(3′)-II. The E. coli transformant was thus only susceptible to fosfomycin, colistin, and tigecycline. PCR-based replicon typing method showed that the plasmid present in transformant and parenteral isolates belonged to the IncR incompatibility group.
Discussion
This report documents for the first time the production of NDM-1 carbapenemase in two K. pneumoniae strains isolated from a patient hospitalized in Algeria. There is no known reservoir of this gene in Algeria. Several hypotheses can be suggested. This carbapenemase has already been described alone or in association with OXA-48 in Tunisia,27,28 where our patient was previously hospitalized. Travelers contribute significantly to the diffusion of bacteria and resistance genes. In our case, it is impossible to know whether the resistant plasmid was acquired in Tunisia or in nosocomial transmission in Ibn Roch Hospital. Indeed, the medical history of hospitalization in the Algerian hospital (the patient was hospitalized for 2 months) could also represent a major risk factor for multidrug-resistant bacteria acquisition. Various mobile genetic elements (e.g., integrons, transposons, and/or insertion sequences) participate in horizontal transfer of the blaNDM gene between different species, notably Acinetobacter spp. to enterobacteria. 29 As recently observed, the NDM-1 enzyme was present in the hospital environment in Algeria, representing an important source/reservoir of nosocomial infections. 7 However, the low local surveillance data impede full assessment of the situation there. The emergence and diffusion of NDM-1-producing strains have been associated with Asia (particularly the Indian subcontinent) and the Balkan region,1,30 which represent the main reservoirs of such pathogens. Interestingly, our isolates belonged to ST147, a worldwide emerging clone of K. pneumoniae strain, diffused throughout North America, European countries, India, Australia, and China with various β-lactamases.3,31–36 In Greece, the clone ST147 has been mainly associated with production of VIM and KPC carbapenemases,37,38 although the emergence of the IncR plasmid encoding VIM MβLs in this clone has been previously described. 37 Finally, ST147 NDM-1-producing K. pneumoniae was recently described in Tunisia, which borders Algeria. 28
Although carbapenemase-encoding enterobacteria frequently carry several plasmids, the results of electroporation clearly suggested that the different resistance genes observed in our strains were transferred through a unique IncR plasmid that also confers resistance to most β-lactams, all aminoglycosides, and fluoroquinolones. Previous studies have described the difficulty in transferring the IncR plasmid.39–41 Electroporation represents the best method to transfer this plasmid because it lacks a transfer system and a relaxase. 39 We encountered difficulties in electroporating this plasmid, with only the plasmid isolated from 19V successfully electroporated into only one bacterial colony after several attempts. Although IncR is difficult to transfer and mobilize under laboratory conditions, it plays an important role in the spread of multidrug resistance genes in enterobacteria. Its role in the transmission of resistance has been attributed to transposition events or plasmid recombination, leading to multireplicons (associations with IncA/C, IncF, IncFIIk, or nontypeable plasmid), contributing to the high plasticity observed in bacterial plasmids.40,41 blaNDM-1 gene was frequently located on IncF-, IncA/C-, IncX3-, IncL/M-, IncN2-, or IncH1B-type plasmids.27,28,32,41,42 IncR blaNDM-1 carrying plasmids have previously been reported in Czech Republic, Poland, the Caribbean, and Singapore.43–47 Most NDM-1 producers are extremely drug resistant: they typically coproduce ESBLs or acquired AmpC enzymes, thus conferring resistance to all β-lactams; they also harbor 16S rRNA methyltransferases, often ArmA or RmtC, which confer resistance to all clinically useful aminoglycosides. This characteristic was previously described in Egypt and Singapore with strains coharboring blaNDM-1, blaCTX-M-15, and rmtF genes.46,47 This was not the case in our isolates in which rmtF (and other methyltransferases) was not detected. This emerging family of plasmids could potentially become widely disseminated because of its quinolone and ESBL resistance as we observed in this study. In a country where the OXA-48-enzyme has become endemic and where the blaNDM gene is increasingly reported in A. baumannii clinical isolates, this emergence is of interest and suggests that the epidemiology of carbapenemase-encoding genes varies around the world, particularly in the Mediterranean region. Microbiologists and clinicians should now be aware of this threat and prepare to implement the necessary control measures to prevent nosocomial infections and a possible spread into the wider population.
The diffusion of NDM-1-producing Enterobacteriaceae continues to advance worldwide. 48 Although recently the Middle East has been considered as an additional and important reservoir for NDM producers, the situation in Maghreb should also raise concerns.
Footnotes
Acknowledgments
We thank Agnes Masnou (Laboratoire de Bactériologie, UFR Pharmacie, Université de Montpellier, Montpellier) for her help in the PFGE experiment and Sarah Kabani for her assistance in preparing and editing the article.
Disclosure Statement
No competing financial interests exist.