
Abstract
Metronidazole (MTZ) is a first-line drug for the treatment of human infections caused by anaerobic bacteria. Although most Clostridium perfringens isolates are susceptible to metronidazole, a number of strains with decreased susceptibility have been obtained in clinical and environmental surveys. However, little is known yet about the toxinotype and genetic diversity of metronidazole-resistant C. perfringens strains. We tested for susceptibility to metronidazole and assessed the toxigenic status and amplified fragment length polymorphism (AFLP) diversity of a collection of 80 C. perfringens isolates of animal origin. In vitro susceptibility testing by the standard agar dilution technique revealed that 22 isolates (27.5% of total) were susceptible to metronidazole, 54 (67.5%) intermediate susceptible, and four (5%) resistant. All isolates belonged to toxinotype A, and two, 41 and 17, of them yielded a positive PCR result for presence of the gene encoding for enterotoxin (cpe), the consensus form of β2 toxin (cpb2) and the atypical form of β2 toxin, respectively. AFLP fingerprinting classified isolates into 65 genotypes indicating a high molecular diversity among isolates in this study, and this diversity was reflected among isolates from each susceptibility category. In conclusion, the results of this study revealed extensive genetic diversity among C. perfringens type A isolates regardless of their susceptibility to metronidazole.
Introduction
T
Antibiotic resistance in anaerobes is an emerging problem of unknown magnitude, as in vitro susceptibility testing of these is not performed routinely.10,11 Metronidazole is still regarded as a first-line drug for the treatment of anaerobe infections in humans and animals10,12,13 and is also widely used for treatment of diverse parasitic infections. 14 Although most isolates of C. perfringens characterized so far seem to be susceptible to metronidazole,15–19 decreased susceptibility to this antimicrobial agent has been reported for some clinical and environmental isolates from different geographic regions,20–23 which suggests that this phenotype might be more frequent than previously thought. However, some of the aforementioned studies only tested a limited number of isolates for which toxinotyping information was not always available. Furthermore, little is known yet about the genetic characteristics of metronidazole-resistant strains.
In this study we assessed the in vitro susceptibility to metronidazole and the toxinotype and genetic diversity, as determined by amplified fragment length polymorphism (AFLP) fingerprinting, of a collection of C. perfringens isolates from diverse animal hosts.
Materials and Methods
Isolates
A total of 80 Clostridium perfringens isolates from the culture collection of our research group (COVEMI, Department of Animal Health, Universidad Complutense de Madrid, Spain) were analyzed (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/mdr). Isolates were recovered from different animal species distributed as follows: pigs (n = 41, 38 intensively raised- and 3 Iberian pigs), dogs (n = 20), oriental cockroaches (Blatta orientalis, n = 15), and other species (n = 4; cat, Dorcas gazelle, plains zebra, and wild boar, one isolate each).
Metronidazole susceptibility testing
In vitro susceptibility to metronidazole was determined by the agar dilution method, following the guidelines of the Clinical Laboratory and Standards Institute (CLSI).23,24 All isolates were tested at least twice on different days, and drug-free plates were included as growth controls. Bacteroides thetaiotaomicron ATCC 29741 was included as quality control in each test run. Minimum inhibitory concentration (MIC) results were used for classification of isolates as susceptible (MIC ≤ 8 mg/L), intermediate susceptible (8 mg/L < MIC < 32 mg/L), or resistant (MIC ≥ 32 mg/L) to metronidazole.23,24
Toxinotyping
The genes encoding for C. perfringens major toxins (cpa, cpb, etx, and iap, for toxins α, β, ɛ, and ι, respectively), enterotoxin (cpe), and β2 toxin (cpb2) were detected using two multiplex PCR protocols, as detailed in a previous publication. 23 In addition, the triplex PCR assay developed by Jost et al. 25 was used to confirm the presence of a consensus or atypical form of the cpb2 gene. In all cases, PCR amplicons were analyzed by electrophoresis on 1.5% agarose gels (Sigma-Aldrich, Madrid, Spain).
AFLP genotyping
Total bacterial DNA (150 ng) was digested using two sets of restriction endonucleases, which included a six-cutter (HindIII in set 1 and EcoRI in set 2; Thermo Fisher Scientific, Waltham, MA) and a four-cutter one (MseI in both sets; Thermo Fisher Scientific), following the recommendations of the manufacturer. After digestion, 5 μl aliquots of the digested products were added to 15 μl ligation mixtures containing 0.1 μM of HindIII or EcoRI adapter (in sets 1 and 2, respectively; Integrated DNA Technologies, Leuven, Belgium), 1 μM of MseI adapter (in both sets; Integrated DNA Technologies) (Supplementary Table S2), 2 μl of 50% (w/v) polyethylene glycol (Thermo Fisher Scientific), and 2 U of T4 DNA ligase (Thermo Fisher Scientific) in 1× T4 DNA ligation buffer (Thermo Fisher Scientific). Ligations were performed for 12 hr at 25°C before enzyme inactivation at 70°C for 10 min.
Five microliters of the 10-fold-diluted ligation products were then used for selective amplification in a total volume of 20 μl under the following conditions: 1× PCR buffer (Biotools, Madrid, Spain), 2.5 mM MgCl2 (Biotools), 62.5 μM each deoxynucleoside triphosphate (Roche Diagnostics GmbH, Mannheim, Germany), 1 μM HindIII-C or EcoRI-0 primer 5′-labeled with 6-carboxyfluorescein (in sets 1 or 2, respectively), 1 μM MseI-G primer (in both sets) (Integrated DNA Technologies; Supplementary Table S2), and 1.5 U of HotSplit DNA polymerase (Biotools). Amplifications were carried out in a GeneAmp PCR System 9700 thermocycler (Applied Biosystems, Foster City, CA) and consisted of 5 min at 94°C, followed by 40 cycles of 20 sec at 94°C, 2 min at 52°C, and 2 min at 72°C, and a final extension at 72°C for 10 min.
The products resulting from the selective amplification step were diluted 1/10 in nuclease-free water (Promega, Madison, WI) and analyzed by capillary electrophoresis using the GeneScan 1200 LIZ size standard (Applied Biosystems). Raw electropherograms were visually checked using Peak Scanner software (Applied Biosystems), and AFLP peaks were scored as absence/presence (0/1) data. Only peaks with a size of 150–1,200 bp and ≥10 % of the height of the highest peak of the individual run were considered, and double peaks were counted only if they were separated by more than 1.5 bp. All AFLP reactions were performed twice on different days for each isolate, and only reproducible peaks were kept in further analyses.
Data analysis
Hierarchical analysis of molecular variance (AMOVA) 26 was used to estimate the partitioning of genotypic variation among and within groups of isolates of different origin and/or metronidazole susceptibility category. Significance of the genetic differentiation was tested using 9,999 permutations. These analyses were performed on the AFLP fingerprinting data using GenAlEx 6.501 software. 27 In addition, a binary 0/1 matrix was created based on the absence/presence of DNA peaks, and a dendrogram of AFLP patterns was built with PAST v.3.11 software 28 using Pearson's correlation coefficients and the unweighted pair group method with arithmetic mean (UPGMA) clustering algorithm. Isolates clustering with <86% similarity were considered to represent different AFLP genotypes, as recommended in previous studies dealing with AFLP typing of other clostridia.29–31
Fisher's exact test and Pearson's chi-square test were used for statistical analysis of categorical data where appropriate. In all cases, two-tailed p-values of <0.05 were considered to be statistically significant.
Results
MIC to metronidazole for the isolates analyzed in this study ranged from 4 to 64 mg/L, and 22 isolates (27.5% of total) were classified as susceptible to this antibiotic, 54 (67.5%) as intermediate susceptible, and 4 (5%) as resistant (Fig. 1 and Supplementary Table S1). Metronidazole-resistant isolates had been obtained from dogs (two isolates), a plains zebra, and a Dorcas gazelle (one isolate each).
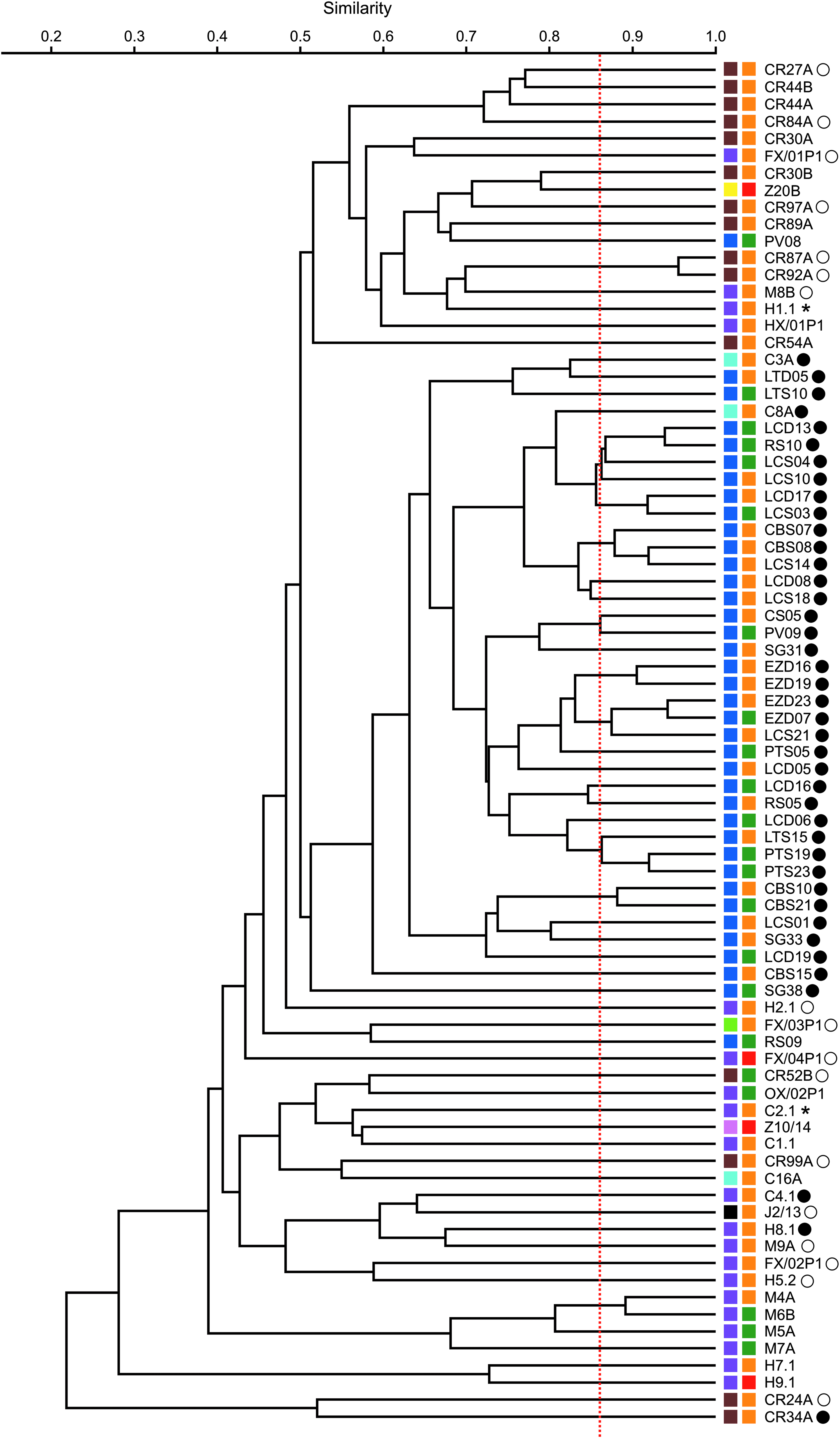
Dendrogram of AFLP profiles obtained for the Clostridium perfringens isolates characterized in this study. The dendrogram was created by UPGMA clustering using Pearson's correlation coefficients. Individual AFLP genotypes are distinguished at ≥86% similarity (red dotted vertical line). Presence of the enterotoxin-encoding gene (cpe) and the consensus or atypical form of the β2 toxin-encoding gene (cpb2) is indicated next to isolates' names by asterisks, filled circles, and open circles, respectively (see also Supplementary Table S1). Color legend for host origin (first column of filled squares): black, wild boar; brown, cockroach; dark blue, intensively-raised pig; light blue, Iberian pig; light green, cat; pink, plains zebra; violet, dog; and yellow, Dorcas gazelle. Color legend for metronidazole susceptibility (second column of filled squares): green, susceptible (MIC ≤ 8 mg/L); orange, intermediate susceptible (8 mg/L < MIC < 32 mg/L); and red, resistant (MIC ≥ 32 mg/L). AFLP, amplified fragment length polymorphism; UPGMA, unweighted pair group method with arithmetic mean.
Multiplex PCR targeting the genes encoding C. perfringens major toxins revealed that all studied isolates had the cpa gene but not other major toxin encoding genes and, thus, belonged to toxinotype A. Only two isolates, both of canine origin and displaying intermediate susceptibility to metronidazole (MIC = 16 mg/L), yielded a positive PCR result for presence of the enterotoxin-encoding gene (cpe) (Fig. 1 and Supplementary Table S1). In addition, 41 isolates (15 susceptible and 26 intermediate susceptible to metronidazole) were positive for presence of the consensus form of the cpb2 gene, and 17 isolates (one susceptible, 15 intermediate susceptible, and 1 resistant to metronidazole) were found to carry an atypical form of the same gene by the triplex PCR procedure described by Jost et al. 25
Notably, no metronidazole-resistant isolate had the gene encoding for the consensus form of β2 toxin. However, there was no association between absence of consensus cpb2 and decreased susceptibility to metronidazole (i.e., MIC > 8 mg/L) either when all studied isolates were considered (n = 80, p = 0.081) or after clone correcting the data set (i.e., removing repeated AFLP genotypes, except when differences in cpb2 status and/or metronidazole susceptibility category were noted; n = 72, p = 0.118).
AFLP fingerprinting resulted in a total of 80 different peak profiles, all of them represented by a single isolate. Regardless of the criterion used for definition of groups in AMOVA analyses, most genetic variation (≥77%) was found at the within-group level (Supplementary Table S3). Significant genetic variation was found between isolates susceptible to metronidazole (MIC ≤ 8 mg/L) and those displaying decreased susceptibility (MIC > 8 mg/L, thus including intermediate susceptible and resistant isolates), and also among isolates from different hosts (p = 0.011 and <0.001, respectively; Supplementary Table S3). However, genetic differentiation was not significant when three metronidazole susceptibility categories (susceptible, intermediate susceptible, and resistant) were considered in the analysis (p = 0.131; Supplementary Table S3).
Furthermore, based on the UPGMA dendrogram (Fig. 1), isolates were classified into 65 AFLP genotypes. Notably, seven AFLP genotypes included isolates belonging to different metronidazole susceptibility categories (intermediate and susceptible, in all cases), whereas the four metronidazole-resistant isolates belonged to different AFLP genotypes.
Discussion
Despite the importance of C. perfringens as a human and animal pathogen, antimicrobial susceptibility testing of isolates from this species is not performed routinely, and thus, resistance rates might be underreported. Furthermore, although there is an increasing interest in studying the molecular epidemiology of C. perfringens in the context of outbreaks of food poisoning in humans32–34 and enteritis in food animals,35–38 detailed molecular comparison of isolates displaying differences in their antimicrobial susceptibility profiles is uncommon (but see, e.g., Álvarez-Pérez et al. 23 and Chalmers et al. 35 for some exceptions to this statement). In this study, we determined the toxigenic status and AFLP genotype diversity of a collection of C. perfringens isolates of animal origin, as well as their in vitro susceptibility to metronidazole.
All isolates characterized in this study belonged to toxinotype A. This toxinotype is highly prevalent in diverse animal species, including pigs and dogs,1,4 which together accounted for 76.3% of our collection of isolates. Only two canine isolates with intermediate susceptibility to metronidazole yielded a positive PCR result for the cpe gene, thus confirming the low prevalence of the enterotoxin-encoding gene among animal isolates. 39 In addition, the consensus and atypical forms of the cpb2 gene were detected in 51.3% and 21.3% of isolates, respectively, but their prevalence varied widely depending on the origin of these. Notably, although we recently found a marginally significant association between absence of consensus cpb2 and decreased susceptibility to metronidazole for waterborne C. perfringens type A strains, 23 such an association was not supported by the data obtained in the present study.
AFLP fingerprinting revealed that the studied isolates were genetically diverse, and high genetic variation was found even among isolates from the same host. Nevertheless, significant genetic variation was found for isolates from different hosts, and most isolates of porcine origin clustered together in the UPGMA dendrogram. Similarly, high intragroup diversity of AFLP profiles but with some degree of genetic differentiation was also detected in the present study for metronidazole susceptible isolates and those displaying decreased susceptibility.
Although we acknowledge that the small number of metronidazole-resistant isolates in our strain collection hinders the assessment of genetic differentiation among isolates belonging to different susceptibility categories, it is noteworthy that the four metronidazole-resistant isolates belonged to different AFLP genotypes that were scattered throughout the UPGMA dendrogram. Similarly, high genetic variability has also been observed among C. perfringens type A strains with resistance to bacitracin.35,40
In conclusion, the results of this study revealed extensive genetic diversity among C. perfringens type A isolates displaying elevated MIC values to metronidazole. Future research should explore if other toxinotypes follow a similar pattern for metronidazole and/or other major antimicrobials.
Footnotes
Acknowledgments
This work was funded by project grant AGL2013-46116-R from the Spanish Ministry of Economy and Competitiveness. Sergio Álvarez-Pérez acknowledges a “Juan de la Cierva” postdoctoral contract [JCI-2012-12396]. The funders had no role in study design, data collection, and interpretation or the decision to submit the work for publication. The authors thank the staff of the Genomics Service at Universidad Complutense de Madrid for providing excellent technical assistance. The constructive comments from two anonymous reviewers are also gratefully acknowledged.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.