
Abstract
Staphylococcal infections are often hard to treat due to increasing resistance, especially to β-lactams. Previous studies described the synergy between common antibiotics and isoflavonoids; however, little is yet known about the combinatory effects of antibiotics with products of human isoflavone metabolism. In this study, demethyltexasin (DT), a human body metabolite of soybean isoflavones, was evaluated for its possible antistaphylococcal combinatory effect with amoxicillin and oxacillin. For comparison, common therapeutically used combination of amoxicillin/clavulanic acid was tested. DT showed strong synergistic interactions against most of Staphylococcus aureus strains when combined with amoxicillin (sum of fractional inhibitory concentrations [ΣFIC] 0.257–0.461) and oxacillin (ΣFIC 0.109–0.484). When oxacillin was combined with DT, resistance to this antibiotic was overcome in many cases. Moreover, antibiotic/DT combinations were effective mainly against methicillin-resistant S. aureus (MRSA); however, the commonly used drug amoxicillin/clavulanic acid was effective only against sensitive strains. Our results indicated DT as a compound able to act synergistically with β-lactams. In addition, some combinations are effective against MRSA and decrease staphylococcal resistance. To the best of our knowledge this is the first report of the antimicrobial synergistic effects of isoflavone human body metabolite with common antibiotics. DT seems to be a possible candidate for further research focused on antistaphylococcal drug development, especially against antibiotic-resistant strains.
Introduction
S
Plants serve as a great source of secondary metabolites in drug discovery. Some of the most widely investigated natural substances are isoflavonoids, which have many times been reported for its various beneficial activities; moreover, there are several studies describing its boosting effect on antibiotics.8,9 It is known that most flavonoids must undergo intestinal transformation by microbiota and enzymes to be incorporated into the blood stream. In addition, this transformation gives rise to diverse beneficial properties, including better antimicrobial activity. 10
Demethyltexasin (DT; 6,7,4’-trihydroxyisoflavone; Fig. 1) originates through hepatic metabolism in the human body as a major hydroxylated metabolite of the well-known legume isoflavones, daidzein and genistein. It has also been detected in many traditional Asian fermented soybean products such a Tempeh or Doenjang, where DT is a product of isoflavone aglycone fermentation process caused mainly by fungi of the genus Rhizopus and by a few bacterial species, such as Brevibacterium epidermidis, Micrococcus luteus, and Microbacterium arborescens. 11 Recently, DT has become of growing scientific interest, mostly because of its remarkable antioxidant and anticancer effects.12,13 In our previous study focused on the antibacterial activity of selected isoflavones, we identified DT as a compound with potential to inhibit S. aureus growth. Moreover, DT showed stronger antimicrobial activity than its precursor genistein. 14
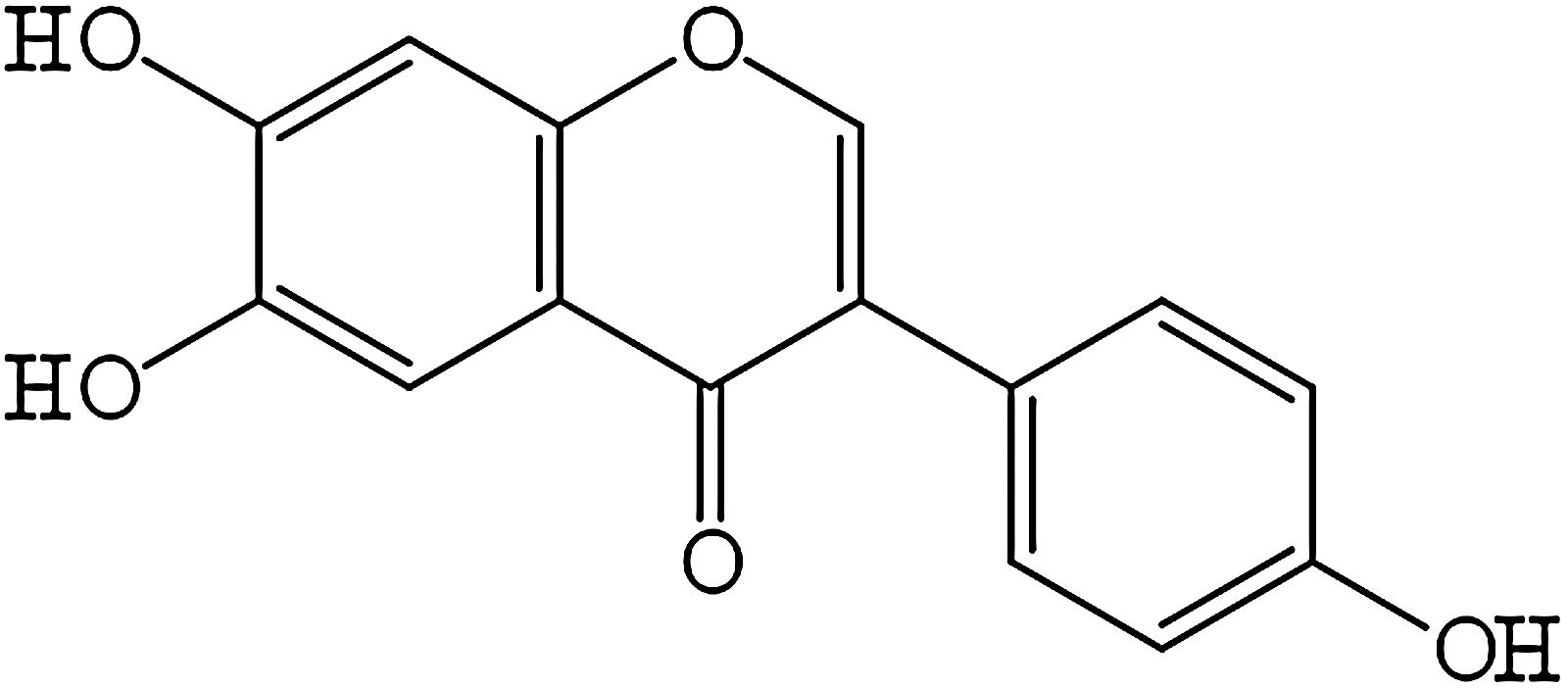
Skeletal formula of demethyltexasin.
In view of the fact that many S. aureus strains are now resistant to antibiotics as well as to some specific compounds capable of lowering bacterial resistance, there is a need to identify novel compounds able to act synergistically with common drugs. Such substances can be, for example, isoflavonoids and their metabolites. Therefore, in the present study we evaluated the in vitro combinatory effect of DT with β-lactam antibiotics (namely amoxicillin and oxacillin) through the checkerboard microdilution method against various sensitive, as well as resistant S. aureus strains. For comparison we also performed experiments with the common pharmacologically applied prophylaxis against MRSA, amoxicillin combined with clavulanic acid.
Materials and Methods
Chemicals
The DT was purchased from Indofine (Hillsborough, USA), whereas the amoxicillin, clavulanic acid (salt potassium clavulanate), and oxacillin came from Sigma-Aldrich (Prague, CZ). Analytical grade solvents (acetonitrile, dimethyl sulfoxide, and ethanol), formic acid, trifluoroacetic acid (TFA), and α-cyano-4-hydroxycinnamic acid (HCCA) were also obtained from Sigma-Aldrich.
Bacterial strains
In this study, fourteen S. aureus strains, including antibiotic-resistant and sensitive forms were used. American Type Culture Collection (ATCC) standard strains 25923, 29213, 33591, 33592, 43300, and BAA 976 were purchased from Oxoid (Basingstoke, UK) on ready-to-use bacteriological Culti-Loops, and clinical isolates (SA1-8) were obtained from The Motol University Hospital (Prague, CZ). Cation-adjusted Mueller–Hinton broth (Oxoid, Basingstoke, UK) equilibrated to pH 7.6 with a Trizma® base (Sigma-Aldrich) was used as cultivation medium.
The identification of clinical isolates was performed by Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS). Briefly, the bacteria were regrown for 24 hr on a Colombia agar with 5% horse blood (Oxoid, Basingstoke, UK). An isolated colony was harvested to 300 μl deionized water with 900 μl of ethanol. The suspension was mixed and centrifuged (2 min at 15,000 rpm), supernatant was discharged, and the pellets were dried. A dry pellet was suspended in 30 μl of 70% formic acid with 30 μl of acetonitrile, mixed, and centrifuged again. Subsequently, 1 μl of the supernatant was deposited on a ground steel target plate (Bruker Daltonics, Bremen, DE) and allowed to dry at room temperature.
The dried sample was afterward overlaid with 1 μl of MALDI matrix (HCCA dissolved in 250 μl of 50:47.5:2.5 acetonitrile:water:TFA solution), dried again, and analyzed by Autoflex Speed MALDI-TOF MS (Bruker Daltonics) operating in a linear positive ion mode. Mass spectra were acquired in the range 2,000–20,137 Da, using a smartbeam frequency of 1,000 Hz, pulsed ion extraction at 220 ns, and a 0.01 kV lens. The voltages for the first and second ion sources were 19.38 and 18.18 kV, respectively. Bacteria were identified using the MALDI Biotyper 3.1 database. Cut-off values higher than two were regarded as relevant for species identification. The mass spectra of each strain were measured in duplicate.
Minimum inhibitory concentration determination and evaluation of combined antimicrobial effect
Individual minimum inhibitory concentrations (MICs) of amoxicillin, clavulanic acid, DT, and oxacillin were determined by the broth microdilution method as described by the Clinical and Laboratory Standards Institute, 15 as modified according to Cos et al. 16 in their recommendations proposed for the effective assessment of the anti-infective potential of natural products, whereas the antistaphylococcal combinatory effect of amoxicillin/clavulanic acid, amoxicillin/DT, and oxacillin/DT were evaluated by the checkerboard method based on fractional inhibitory concentrations (FICs), 17 both performed on 96-well microtiter plates.
In combinations, eight two-fold serial dilutions of antibiotic (amoxicillin or oxacillin) from horizontal rows of the microtiter plate were subsequently crossdiluted vertically by eight two-fold serial dilutions of the test compound (DT or clavulanic acid). Microplates so arranged can be used to screen 64 different combinations of concentrations. The initial concentrations used in the combinations for both clavulanic acid and DT were 256 mg/L, whereas for amoxicillin and oxacillin various starting concentrations were used depending on the staphylococcal strain's susceptibility to the antibiotics tested.
Assay microplate preparation and serial dilution were performed through the automated pipetting platform Freedom EVO 100 equipped with a four-channel liquid handling arm (Tecan, Männedorf, CH). Plates were inoculated by bacterial suspension (final density 5 × 105 cfu/ml) and incubated at 37°C for 24 hr. The bacterial growth was then measured spectrophotometrically using a Multimode Reader Cytation 3 (BioTek Instruments, Winooski, VT) at 405 nm.
MICs were expressed as the lowest concentrations that inhibited bacterial growth by ≥80% compared with that of the agent-free growth control. The MICs presented in this work are the average of MICs obtained from three independent experiments performed in triplicate.
The combined effects of the antibiotics with the tested compound (A and B) were then determined based on the value of ΣFIC, which was calculated according to the following equation: ΣFIC = FICA+FICB, where FICA = MICA(in the presence of B)/MICA(alone), and FICB = MICB(in the presence of A)/MICB(alone). In view of the fact that it is a widely accepted norm in MIC testing that variation in a single result places an MIC in a three-dilution range (mode ±1 dilution), the possibilities for reproducibility errors in an MIC checkerboard are considerable. Within the limits of experimental error, ΣFIC really indicates only “synergy,” “no interaction,” and “antagonism” between agents. 18
Therefore, in this study, the antimicrobial combinatory effect was interpreted according to Odds 18 as follows: a synergistic effect if ΣFIC ≤ 0.5; no interaction if ΣFIC > 0.5–4, and antagonistic if ΣFIC > 4. S. aureus 29213 was used as a control strain for antibiotic susceptibility testing and strains were identified as MRSA when oxacillin MIC was ≥ 4 mg/L. 15
Solvents used as the negative control did not inhibit any strain tested.
Results
In this study, DT demonstrated significant potentiating antistaphylococcal activity of β-lactam antibiotics especially against MRSA. Data summarized in Table 1 show many synergistic interactions together with no antagonism occurrence.
Bold values: synergy (ΣFIC ≤ 0.5).
Sensitive strain (oxacillin MIC < 4 mg/L).
Methicillin-resistant S. aureus strain (oxacillin MIC ≥4 mg/L).
AMX, amoxicillin; CLA, clavulanic acid; DT, demethyltexasin; OXA, oxacillin; MIC, minimum inhibitory concentration (expressed as an average from three independent tests set in triplicate); ΣFIC, sum of fractional inhibitory concentrations.
DT combined with oxacillin possessed synergy against ten out of fourteen S. aureus strains with a range of ΣFIC 0.109–0.484. Moreover, all of these ten strains were MRSA (oxacillin MICs ranging from 16 to 341 mg/L) and their resistance to oxacillin was overcome in most of them when the DT/oxacillin combination was used. The strongest synergy (ΣFIC 0.109) was obtained against the clinical isolate SA3 at a DT concentration of 8 mg/L, when a 64-fold oxacillin MIC decrease was achieved (from 64 to 1 mg/L). The highest oxacillin MIC reduction was observed for MRSA 33591, where a DT concentration of 32 mg/L caused a 170-fold oxacillin MIC reduction (from 341.333 to 2 mg/L) with a ΣFIC value of 0.381.
In view of the fact that amoxicillin combined with clavulanic acid is a commonly used therapeutic combination, especially as a prophylaxis against the development of MRSA infections, we decided to combine amoxicillin also with DT against four S. aureus strains. Similarly, as in the case of oxacillin, these results showed many synergistic interactions between DT and amoxicillin for all three tested MRSAs (ΣFIC 0.257–0.461); however, against the sensitive strain there was no interaction. The highest amoxicillin MIC decrease (a 43-fold reduction) was obtained for MRSA ATCC 43300 in a DT concentration of 32 mg/L (ΣFIC 0.398). On the other hand, while the combination of amoxicillin/clavulanic acid displayed a synergy against sensitive S. aureus (ΣFIC 0.319–0.396), there was only one synergistic effect against MRSA (SA6) causing only a two-fold amoxicillin MIC reduction (ΣFIC 0.495).
Discussion
The enhancing effect of isoflavonoids on the activity of antibiotics has been reported several times over the last few years. The most often tested compound is biochanin A, which has shown synergy with ciprofloxacin against eleven MRSA clinical isolates (ΣFIC 0.13–0.5) and one standard S. aureus ATCC 25923 (ΣFIC 0.5). 19 Wang et al. 20 described the antistaphylococcal combinatory effect of genistein together with norfloxacin and of diosmetin with ciprofloxacin, norfloxacin, and streptomycin.
In our study, DT also provided strong synergistic effects against all MRSA strains tested. Moreover, it was able to cause up to a 170-fold reduction in oxacillin MICs. Our results can be compared with the review by Cushnie and Lamb, 21 where they indicated that the most potent synergistic flavonoids/antibiotic antistaphylococcal combinations were able to cause from a 16 to a 1,024-fold reduction in antibiotic MICs. Therefore, any combination DT causing up to a 170-fold MIC reduction can be noted as a compound with promising potential for further pharmacological research focused on the development of new pharmaceuticals able to overcome microbial drug resistance. In addition, our results suggest that not only plant isoflavones, but also their metabolic products are able to synergistically interact with conventional antibiotics.
Several modes of action have been proposed for the flavonoids' antimicrobial, synergistic, and antibiotic resistance-modulating activity; however, the exact mechanisms found in the isoflavonoid group are still not clear. It has been shown that there is a significant correlation between their antistaphylococcal effect and the presence of several functional groups at particular positions in isoflavonoid structure. Their hydroxyl groups have obvious affinity to proteins and their presence at the position C7 and/or C5 is important. 22 DT has one of its hydroxyl groups at the C7 position; moreover, it is also present at the C4 position, which is thought to increase antimicrobial effectiveness. 23
In the case of synergy with β-lactams, certain principles can be in play: the reduction of D-alanylation of teichoic acid in the bacterial cell wall resulting in the deactivation of modified penicillin-binding proteins 2a (PBP2a, responsible for resistance due to low affinity to β-lactams), intercalation into the cytoplasmic membrane inducing structural changes and delocalization of PBP2a or disruption of its synthesis, the inhibition of the production of β-lactamases (enzymes able to destroy the β-lactam ring in antibiotics), the inactivation of the efflux pump, the destabilization of the bacterial cytoplasmic membrane, and topoisomerase inhibition resulting in the prevention of DNA replication. However, none of these processes can fully explain the synergistic effect, so it is proposed that isoflavonoids exert this activity through a combination of different mechanisms. 21
Regarding the toxicology of isoflavonoids, they are known for their unique properties such a good biodegradability and biocompatibility together with a lack of toxicity as edible products. 24 Generally, the toxicity of flavonoids is considered to be very low. For example in rats LD50 was found to be between 2 to 10 grams per animal. 25 In the case of DT Lee et al. 13 they observed no effect on normal healthy cells in vitro. Furthermore, isoflavones are regarded a safe and partially effective dietary alternative to hormone replacement therapy. 26 As we know, DT occurs in fermented soy-based food products, which are regularly consumed mostly in Asian countries 10 and, after metabolism in the liver, it is excreted in urine. 27 Therefore, DT might be supposed to be nontoxic to humans; under any of these conditions, detailed toxicological studies are needed.
It is well known that the effectiveness of antibiotic therapy can be affected by daily diet. The negative effect is usually of more interest because of the potential to harm the patient, due to the changes in the pharmaceutical, pharmacokinetic, or pharmacodynamic properties of the particular drug. The most important food–drug negative interactions are connected with fruit juices, mostly with grapefruit juice, able to increase drug bioavailability in the blood stream and thus increase the possibility of worse adverse effects. Likewise, the coadministration of milk products can influence antibiotic activity, preventing their absorption and thus predisposing to treatment failure.
On the other hand, there are many suggestions that food can also have a positive effect on antibiotic therapy, despite there being a lack of formal evidence for such.28,29 As reviewed by Hemaiswarya and Doble, 30 several herbal extracts or fruit and vegetable phenols (e.g., quercetin and resveratrol) enhance the therapeutic effect of anticancer drugs, and their consumption is beneficial. A high content of fruit, vegetables, nuts, and grains in the daily diet can bring to the organism a large proportion of antioxidants, vitamins, flavonoids, and phenols, which may interact with drugs, and thus increase their activity. 31
In view of the fact that DT is originated in human livers from genistein, is found in a number of leguminous plants [14], and has the ability to act synergistically with β-lactam antibiotics, we supposed that a genistein-rich diet can positively affect antibiotic therapy. However, further research is needed to confirm this suggestion.
In summary, our study shows a significant in vitro antistaphylococcal synergistic effect of DT combined with amoxicillin and oxacillin. To the best of our knowledge, this is the first report of the antimicrobial combinatory effect of isoflavone's human body metabolite with antibiotics. As a result of our series of experiments, DT has shown many synergistic interactions with amoxicillin and oxacillin antibiotics, together with no antagonism against various strains of S. aureus. Furthermore, when compared with an amoxicillin/clavulanic acid combination, which is active mainly against MSSA, our results with amoxicillin/DT showed a synergy mostly against MRSA.
Since the activity of both β-lactams was significantly increased when combined with DT and effective combinations can overcome antibiotic bacterial resistance, DT seems to be a promising substance for further research focused on the development of new, synergistically acting antistaphylococcal drugs especially for use against antibiotic-resistant strains. Moreover, as DT is present in many soybean products it seems that the consumption of such food could have a positive impact on the medicinal effect of antibiotic therapy. However, further research is needed before its possible use, for example, to explain the exact mechanism of synergistic action and detailed toxicological studies.
Footnotes
Acknowledgments
This work was financially supported by grants from the Internal Grant Agency of the Faculty of Tropical AgriSciences (IGA 20165009), by the Grant Agency of the Czech University of Life Sciences, Prague (CIGA 20175001), and by the National Agency for Agricultural Research, the Ministry of Agriculture, Czech Republic (QJ1510038). The authors are grateful to Mícheál Úa Séaghdha for his final linguistic revision of the English text.
Disclosure Statement
No competing financial interests exist.