
Abstract
This study examined antimicrobial resistance (AMR) profiles in commensal Escherichia coli derived from healthy fattening pigs in Thai farms that used prophylactic antimicrobials (in-feed tiamulin fumarate and amoxicillin) [PAs], therapeutic antimicrobials (injectable enrofloxacin or gentamicin) [TAs], or no antimicrobials [NAs]. Commensal E. coli were used as a proxy for overall AMR on the farms. There was a high level of multidrug resistance in all three categories of farm, with isolates showing resistance to β-lactams (amoxicillin, ampicillin, and piperacillin) and tetracyclines (tetracycline), and commonly possessing tetA, blaTEM, and plasmid replicons FIB and F. On the other hand, isolates with an extended-spectrum beta-lactamase phenotype (ESBLP) and with resistance to aminoglycosides, chloramphenicol, fluoroquinolones, nitrofurantoin, tiamulin, and trimethoprim/sulfamethoxazole were significantly more common among the PA farms (p < 0.05) than in the other two farm categories. In the PA farms, ESBLP E. coli commonly contained the blaCTX-M-1 group, blaCTX-M-9 group, or both gene groups, and were shown to transfer blaCTX-M genes in a conjugation experiment. E. coli containing N, FIC and A/C replicons were found only in PA farms. In summary, although E. coli isolates from all farms contained a core set of resistance to β-lactams and tetracyclines, the routine use of PA increased resistance rates to other important antimicrobials.
Introduction
E
Although monitoring studies has reported a high prevalence of multidrug-resistant (MDR) E. coli in pigs in Thailand (60–90%), the antimicrobials used on the farms and the ages of the animals sampled have not always been clearly defined. 3 The occurrence of AMR among commensal E. coli isolated from young piglets in Thailand has been highlighted previously, including the distribution of transferable genes encoding. 4 Nevertheless, examination of MDR E. coli derived from older fattening or slaughter-aged pigs should give a better insight into the potential risks of transmission to the consumer. 5 Furthermore, comparing resistance among isolates from farms with and without routine prophylactic antimicrobial (PA) and therapeutic antimicrobial (TA) use should give an indication about the degree to which this use may provide selective pressures encouraging development of resistance.6–9
The specific objective of this study was to determine the existence and basis of AMR in commensal E. coli isolated from fattening pigs in Thailand and compare the AMR profiles between isolates from farms with and without routine PA and TA use. The hypothesis being tested was that commensal E. coli isolates from grower pigs on farms without routine antimicrobial use would show less AMR compared to isolates from farms with either routine PA or TA use.
Materials and Methods
Study area and animal selection
Samples were collected from 25 farms located across five Provinces of Thailand. The farms were selected based on their antimicrobial usage patterns. Twelve farms routinely used PAs; two only used antimicrobials therapeutically, TAs; and 11 small farms had never used antimicrobials, NAs. The PA and TA farms had similar management systems, including open housing, the same vaccine program, and multisite production. They all had over 1,000 sows, had no pig replacement from outside sources, and had consistent management for at least 2 years in terms of antimicrobial use and sanitary and biosecurity measures taken. They were well managed and kept good records of production and antimicrobial use. The NA farms were all small (<50 sows) village-based enterprises, where the houses were open sided and contained pigs of different ages and stages of production. Feedstuffs were locally sourced.
For the 12 PA farms, a routine in-feed antimicrobial use program had been followed for at least 2 years for endemic bacterial disease prophylaxis. This included incorporating tiamulin fumarate at 100 parts per million (ppm) and amoxicillin at 250 ppm in the feed in the nursery (6–8 weeks) and grower (8–16 weeks) phases of production. Where individual pigs became sick, they were isolated and treated with TAs as appropriate.
A total of 102 fecal samples from individual 18- to 20-week-old fattening pigs with a normal appearance and no recent history of enteric or respiratory disease, or TA treatment, were obtained from the farms (7–10 samples per farm). A total of 70 fecal samples were obtained from similar fattening pigs on two TA farms that did not use PAs and only used the injectable antimicrobial enrofloxacin or gentamicin under veterinary prescription for individual treatment of specific clinical cases where the sick pigs were kept in isolation. Another 67 samples were collected from fattening pigs on the 11 NA farms, where there was no access to or use of antimicrobials or vaccination.
Sample collection and bacterial identification
The pig sampling protocol was approved by the Chulalongkorn University Animal Care and Use Committee (permit number 58/2558). For each pig, at least 5 g of rectal feces was collected directly into a sterile container and delivered to the laboratory at 4°C. All fecal samples were diluted 10-fold using sterile 0.85% NaCl and the dilutions of 10 −7–10−8 suspensions were spread on Eosin Methylene Blue Agar 10 (Oxoid, United Kingdom) plates to select the major population of bacteria. E. coli colonies were identified by their IMViC (Oxoid) biochemical reactions, comprising an indole test (+), a methyl red test (+), a Voges-Proskauer test (−), and a citrate test (−). 11 A representative pure colony from the highest dilution plate was randomly selected for further investigation.
Phylogenetic grouping
All isolates were characterized into their phylogroups using a published multiplex polymerase chain reaction (PCR)-based method that identifies eight phylogroups (A, B1, B2, C, D, E, F, and cryptic clades). 12 E. coli ATCC 25922 and E. fergusonii CUVET427 were used as the control strains.
Phenotypic resistance characterization and extended-spectrum beta-lactamase phenotype confirmation
The minimal inhibitory concentration of antimicrobials for the E. coli isolates was determined using the AST-GN 38 test kit in the Vitek2 compact automated susceptibility level detection apparatus (BioMérieux, France), except for tiamulin (Sigma-Aldrich, St. Louis, MO) where susceptibility testing was performed using an agar dilution test with results interpreted according to Clinical Laboratory Standard Institute (CLSI) standards. The 19 antimicrobials or antimicrobial combinations tested were amikacin (AK), amoxicillin (AMX), amoxicillin–clavulanic acid (AMC), ampicillin (AMP), cefpirome (CPR), cefpodoxime (CPD), cephalexin (CEX), ceftiofur (XNL), chloramphenicol (C), enrofloxacin (ENR), gentamicin (GM), imipenem (IMP), marbofloxacin (MBR), nitrofurantoin (NT), piperacillin (PIP), tetracycline (TE), tiamulin (TI), tobramycin (TM), and trimethoprim/sulfamethoxazole (SXT).
In the Vitek2 machine, the set of antimicrobial arrays was harmonized to match with veterinary guidelines, 13 including some drugs that were allowed for use in veterinary practice in the past. 14 E. coli ATCC 25922, Pseudomonas aeruginosa ATCC 27853, and Staphylococcus aureus ATCC 25913 were used as the control strains. The interpretation of susceptibility levels for AMP, CPD, XNL, GM, ENR, MBR, TE, C, and TI followed the CLSI standards for antimicrobial disks and testing for bacteria isolated from animals (VET1-0S3), 15 and the interpretation for AMX, PIP, AMC, CEX, CPR, IMP, AK, TM, NT, and SXT followed the CLSI standards for antimicrobial susceptibility testing (M100-S25). 16
ESBL phenotypic screening and confirmatory test
The E. coli isolates were screen for ESBL production using the Vitek2 machine (BioMérieux, France). 17 As a confirmation test for ESBLs, the combination disk test was performed for all isolates, as recommended. 16
blaCTX-M gene detection
The gene blaCTX-M comprising variants blaCTX-M-1, blaCTX-M-2, blaCTX-M-9, blaCTX-M-8, and blaCTX-M25/26 was detected by multiplex PCR in all extended-spectrum beta-lactamase phenotype (ESBLP) E. coli strains. 18 The identity of representative PCR amplicons was confirmed by DNA sequencing and analyzed using BioEdit version 7.0.0, with comparisons made to the GenBank database.
blaCTX-M conjugative assay
To determine whether blaCTX-M genes were located on transmissible plasmids, a conjugation assay was performed using the broth mating technique, as recommended. 19 The two selected donors used were E. coli PCU12_1 (positive for blaCTX-M-1 and a single F replicon) and E. coli PCU12_2 (positive for blaCTX-M-9 and a single FIB replicon). The recipient strain E. coli J53 was resistant to sodium azide: Azir. Transconjugants were selected on Luria Bertani agar (Oxoid) supplemented with cefotaxime (2 μg/ml) and sodium azide (100 μg/ml) (Oxoid). Antimicrobial susceptibility, a confirmatory test for ESBL phenotype, PCR detection, and DNA sequencing of blaCTX-M genes were performed on the transconjugants.
Genes encoding AMR
Bacterial DNA was extracted using a Wizard ® Genomic DNA Purification Kit (Promega, Germany). A total of 16 pairs of primers specific for resistance genes in the Enterobacteriaceae family were generated (First Base Laboratories Sdn Bhd, Seri Kembangan, Selangor, Malaysia) and PCR thermal cycling conditions followed previous recommendations. 20 Amplified genes included blaTEM and blaPSE-1 for ampicillin, amoxicillin, and piperacillin resistance, aadA1 and aadA2 for streptomycin resistance, aadB for tobramycin and gentamicin resistance, tetA and tetB for tetracycline resistance, dfrA1, dfrA10, and dfrA12 for trimethoprim resistance, sul1, sul2, and sul3 for sulfonamide resistance, and catA, catB, and cmlA for chloramphenicol resistance. A representative positive PCR amplicon for each gene was submitted for DNA sequencing and analyzed by Bioedit version 7.0.0 (Ibis Biosciences) using the GenBank database.
Plasmid replicon typing and intI detection
The presence of 18 plasmid replicons that are widespread in Enterobacteriaceae, comprising F (FIA, FIB, FIC, FIIA, and F), I1-lγ, N, P, W, HI1, HI2, L/M, T, A/C, K, B/O, X, and Y was investigated using five multiplex and three simplex PCR tests. The PCR conditions and thermal cycles were performed according to a previous study. 21 The intI gene representing integron I cassettes was detected by a simplex PCR following a previously published method. 22 Representative positive PCR amplicons for each replicon were submitted for DNA sequencing and analyzed as described above.
Data analysis
Resistance rates, replicon positivity rates, and resistance gene profiles were presented as percentages, and the AMR profiles were reported in antibiogram patterns for the three categories of farms. SPSS version 17.0 (IBM, Armonk, NY) was used to calculate odds ratios (ORs) and 95% confidence intervals (CIs) for each resistance phenotype and each replicon type, and each resistance gene between isolates from the three different categories of farm. The types and proportion of phylogroups between the farm groups were compared using the Kruskal–Wallis H test (K–W).
Results
Distribution of phylogroups
The distribution of phylogroups of the commensal E. coli tested from the three categories of farms is shown in Figure 1. The types and proportion of phylogroups of the selected E. coli colonies were quite similar in all farms (p = 0.894). Phylogroup A (PA: 47.1%, TA: 48.6%, NA: 44.8%), B1 (PA: 29.4%, TA: 25.7%, NA: 29.9%), and E (PA: 13.7%, TA: 12.9%, NA: 14.9%) were the most commonly detected groups. Phylogroups B2, C, D, and F were minor components and in total were detected at <14% in each farm category (PA: 9.8%, TA: 12.9%, and NA: 10.4%). Representatives of the cryptic clade group were not found (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/mdr).
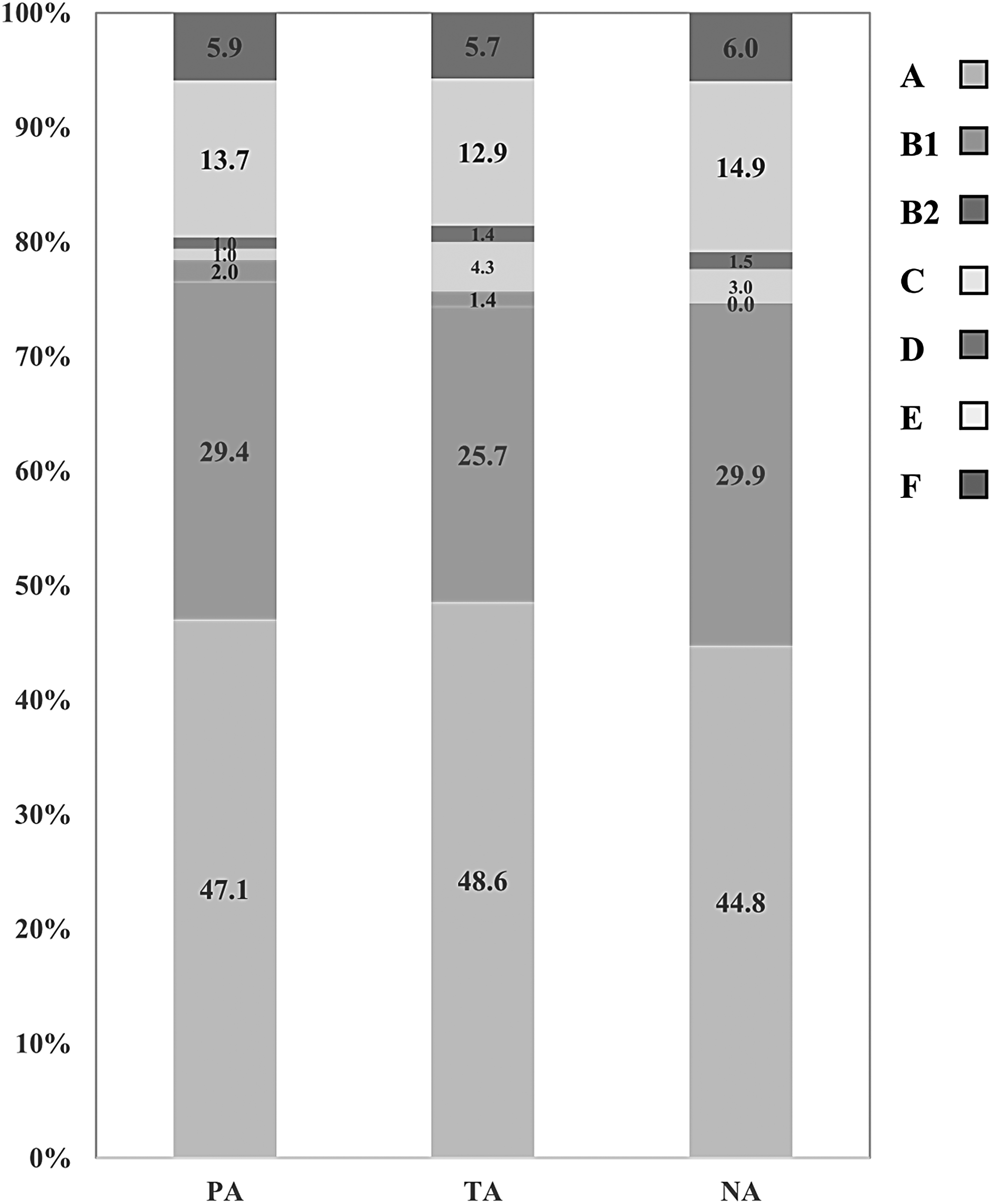
The relative abundance of phylogroup in porcine commensal Escherichia coli from the three categories of farms. Vertical bar, PA, TA, and NA. NAs, no antimicrobials; PAs, prophylactic antimicrobials; TAs, therapeutic antimicrobials.
Distribution of phenotypic AMR rates
The distribution of phenotypic resistance rates for the commensal E. coli against the antimicrobials tested in the three categories of farms is shown in Figure 2. Amikacin- and imipenem-resistant E. coli were not found in any of the farms, and resistance to amoxicillin/clavulanic and cefpirome was found only in the PA farms, and was uncommon.

Comparison of rates of AMR in porcine commensal E. coli from the three categories of farms. Open horizontal bar, NA; grey bar, TA; black bar, PA. AMR, antimicrobial resistance.
The isolates from the three categories of farms had very high rates of resistance to tetracycline, amoxicillin, ampicillin, and piperacillin (over 83.6%), and these rates did not differ significantly between the farm categories. The isolates from the PA farms had resistance rates to the other antimicrobials tested, which varied from 52.9% to 100%, while rates were much lower (0% to 42.3%) for the various antimicrobials in the other two farm categories. The rates in the PA farms were statistically significantly higher than those in both the TA and NA farms, but were not significantly different between the latter two categories. Within each of the three categories of farms, resistance rates for each drug between farms did not reveal any significant farm-related differences (Supplementary Fig. S2).
Over 50% (64/102) of the PA isolates were resistant to gentamicin, tobramycin, enrofloxacin, marbofloxacin, nitrofurantoin, chloramphenicol, and sulfamethoxazole/trimethoprim. The antibiogram AMX-AMP-PIP-CEX-CPD-XNL-GM-TM-ENR-MBR-TE-NT-C-SXT-TI was the most prevalent type found (n = 28, 27.5%) in the isolates from the PA farms. On the other hand, 18.6–41.4% of isolates from the TA farms were resistant to gentamicin, tobramycin, nitrofurantoin, enrofloxacin, marbofloxacin, chloramphenicol, and sulfamethoxazole/trimethoprim. The antibiogram AMX-AMP-PIP-ENR-MBR-TE-NT-C-SXT-TI was the most prevalent type found (n = 12, 17.1%) in E. coli from TA farms, while AMX-AMP-PIP-TE-C was the most prevalent type found in isolates from the NA farms (n = 10, 14.9%).
AMR genotypes
The resistance genes blaTEM and tetA were the most common type found among E. coli isolates from pigs on all farms (over 82.9%), with no significant differences in rates between the three farm categories (Fig. 3). The gene blaCTX-M-9 was only found in isolates from PA farms, and at a low rate. The other resistance genes, except for dfrA, were significantly more common in isolates from the PA farms than from the other two categories. Rates in TA and NA farms did not differ significantly from each other. The percentage of isolates containing catA, catB, dfrA12, sul3, tetB aadA1, aadA2, and aadB ranged from 47.1% to 67.6% in PA farms, whereas they were only present at 14.3–35.8% in isolates from the other two farm categories (Fig. 3).

Comparison of AMR genes of porcine commensal E. coli from the three categories of farms. Open horizontal bar, NA; grey bar, TA; black bar, PA.
Plasmid replicon types and confirmation of blaCTX-M gene conjugation
Plasmid replicons X, W, P, T, L/M, K, FIIAs, and B/O were not detected in this study. The two most common types that were found were F and FIB, and there were no significant differences in their rates of occurrence between the three categories of farms (Fig. 4). Rates for H12 were low and were not significantly different between the three categories of farms. Replicon types FIC, N, and A/C were only found in isolates from PA farms. All the other replicon types, while at a low prevalence, were significantly more common in isolates from the PA farms than in the other two categories, which did not differ significantly from each other. The intI gene was found at a significantly higher rate in isolates from the PA farms compared to isolates from both the TA and NA farms, while the rates in TA and NA farms were similar (Fig. 3).

Comparison of plasmid replicon types detected in porcine commensal E. coli among the three categories of farms. Open horizontal bar, NA; grey bar, TA; black bar, PA.
Conjugation assays showed that the F and FIB replicon plasmids from ESBLP were transferred with a frequency of 1.36 × 10−3 and 8.67 × 10−4 to transconjugants, respectively. The donor ESBLP strains E. coli PCU12_1 (positive for blaCTX-M-1 and with a single F replicon) and E. coli PCU12_2 (positive for blaCTX-M-9 and with a single FIB replicon) transferred blaCTX-M-1 and blaCTX-M-9 to recipient E. coli strain J53, confirming the location of the genes on these conjugative plasmids.
Using automated detection with confirmation by the combination disk assay, ESBLP E. coli were detected in 54 of the 102 (52.9%) E. coli from the PA farms, in 5 of 70 (7.1%) E. coli from the TA farms, and in none of the isolates from the NA farms (Table 1). Antibiograms, replicon type, and blaCTX-M genes of 59 ESBLP E. coli are shown in Table 1. ESBLP strains derived from PA farms contained blaCTX-M-1 group (35, 64.8%), blaCTX-M-9 group (17, 31.5%), or both groups of genes (2, 3.7%).
ESBLP incidence rate in PA farms was significantly greater than in TA and NA farms (Chi-square test at p < 0.05).
AMC, amoxicillin–clavulanic acid; AMP, ampicillin; AMX, amoxicillin; C, chloramphenicol; CEX, cephalexin; CI, confidence interval; CPD, cefpodoxime; ENR, enrofloxacin; GM, gentamicin; MBR, marbofloxacin; NA, no antimicrobial; NT, nitrofurantoin; OR, odds ratio; PA, prophylactic antimicrobial; PIP, piperacillin; SXT, trimethoprim/sulfamethoxazole; TA, therapeutic antimicrobial; TE, tetracycline; TI, tiamulin; TM, tobramycin; XNL, ceftiofur.
The five ESBLP E. coli from the TA farms only contained blaCTX-M-1. The most common antibiogram for the ESBLP E. coli was AMX-AMP-PIP-CEX-CPD-XNL-GM-TM-ENR-MBR-TE-NT-C-SXT in the PA farms (n = 28) and AMX-AMP-PIP-CEX-CPD-XNL-NT-SXT in the TA farms (n = 2). The genes blaCTX-M-2, blaCTX-M-8, and blaCTX-M-25/26 were not detected in this study. Replicons FIB (37.0%), F (22.1%), or N (18.5%) were commonly detected in the ESBLP E. coli from the PA farms, and FIB was the only replicon detected in ESBLP E. coli from the TA farms.
Discussion
Background
Antimicrobials are commonly used in the Thai pig industry to control endemic bacterial diseases in intensive farms. Many large intensive farms in Thailand use antimicrobials prophylactically in specific age groups of pigs, in anticipation of regular bacterial disease outbreaks, while some only use targeted antimicrobials for treatment of individual sick pigs.
This study aimed to investigate and compare AMR rates in commensal E. coli from fattening pigs on farms that had different patterns of antimicrobial usage: farms routinely using PAs; farms only using antimicrobials for specific therapy; and farms not using antimicrobials at all. Similarly sized and managed farms in the first two categories were included in this study. The two farms that only administered TAs to sick pigs were somewhat unusual in that they only used injectable enrofloxacin or gentamicin, and this choice might limit the development of resistance compared to the more typical use of a broader range of targeted antimicrobials on many farms.
Unfortunately, it was not possible to match the husbandry and conditions in the third category of farm with those in the first two categories, as the only farms that could be identified as not using antimicrobials at all were small village-based piggeries where they had neither access to nor the financial capacity to purchase antimicrobials. It is possible that other issues associated with factors such as stocking rates, nutrition, or genetics may have had an influence on the findings on these farms.
Phylogroups
The phylogroups of the selected E. coli colonies were determined to ensure that antimicrobial use had not altered the phylotype profiles on the farms, and differences in antimicrobial susceptibility profiles were not biased by the choice of colonies. The analysis confirmed that there was no significant difference in the distribution of phylogroups of E. coli selected between the three groups of farms. Representatives of all seven phylogroups were found, with groups A, B1, and E predominating. No correlation was found between the phylogenetic background of the isolates and their antibiograms.
Rates of phenotypic resistance
The rates of phenotypic resistance among the commensal E. coli from all three categories of farms for piperacillin, amoxicillin, ampicillin, and tetracycline were similar and very high. This indicates that there is a widespread and almost universal set of resistances to these drugs in E. coli from fattening pigs in Thailand, whether or not they have been exposed to antimicrobials in the past.
This is disturbing since it implies that this resistance is not reversible, even in the absence of antimicrobial selection pressure. The resistance phenotypes found were associated with the common occurrence of tetA and blaTEM, genes that frequently have been reported in porcine commensal E. coli in Southeast Asia.3,23–26 Our study suggests the existence of resistant clones that are widespread and persist in the environment. According to the fitness-cost model, tetA and blaTEM might be maintained in E. coli in animals and the environment without needing selective pressure from the use of tetracyclines and β-lactams, respectively.27,28 As a consequence, resistance to β-lactams and tetracycline is not a useful proxy to reflect the overall trends in AMR in surveillance programs.
In relation to the other antimicrobials tested, no resistance was found to amikacin and imipenem, and resistance to amoxicillin/clavulanic and cefpirome was found only in the PA farms, and only in a few isolates. On the other hand, high rates of resistance to all the other antimicrobials tested were commonly found in isolates from PA farms, but were less commonly found in isolates from the other farm categories; furthermore, the resistance rates for enrofloxacin, chloramphenicol, sulfamethoxazole/trimethoprim, and nitrofurantoin were substantially higher than previously reported in the pig industry in Thailand.3,4,26
Importantly, apart from tiamulin fumarate and amoxicillin, the other antimicrobials had not been used in pigs in the PA farms, including chloramphenicol and nitrofurantoin that have been banned from use in Thai livestock since 1999. 29 From this, it seems that the regular prophylactic use of tiamulin fumarate and/or amoxicillin has been selected for commensal E. coli that are resistant to these two antimicrobials in the PA farms, and also for coresistance in these isolates to the other antimicrobials that were tested. For example, coselection for resistance to these antimicrobials may have occurred with resistance to amoxicillin on the same mobile genetic elements or adjacent genes. 30 Resistance to tiamulin mainly derives from chromosomal mutations in the 23S rRNA and rplC genes that result in reduced drug binding, and consequently may lead to cross-resistance to other antimicrobials that target the bacterial ribosome. 31
In addition, genes such as cfr that encode rRNA methyltransferase, which methylates position A2503 on the 23S rRNA and confers resistance to chloramphenicol, florfenicol, clindamycin, pleuromutilins (tiamulin), oxazolidinones, and streptogramin, can be located on plasmids or transposons that are transferable between bacterial species. 32 Their transfer potentially also could coselect for resistance genes located on these elements. An investigation into the genetic basis of resistance to tiamulin in these isolates, and potential cross-resistance to other drugs, will be the subject of a future study.
Genotypic resistance
The genes aadA1, aadA2, aadB, sul1, sul2, sul3, dfrA12, catA, catB, and cmlA, as well as the integron gene intI were detected in isolates from the PA group at a significantly higher rate than in isolates from the TA and NA groups. This group of resistance genes is usually located on class I integrons.,33–35 and especially, dfrA1, dfrA12, aadA1, and aadA2 are commonly found on class I integron gene cassettes in isolates from Southeast Asia.3,26 This group of resistance genes might be class 1 integron-borne resistance determinants that were coselected with integron I and have become widely spread among isolates from PA farms under ongoing antimicrobial selection pressure.
ESBLP E. coli
Generally, the occurrence of ESBL-producing bacteria in pigs varies depending on microenvironmental factors and the growth phase of the pigs. 36 Commensal strains of E. coli are considered to represent an important reservoir of ESBLs. 37 This study found a high rate of commensal ESBLP E. coli in pigs close to slaughter age on PA farms (Table 1), and this rate was greater than in younger pigs that previously have been tested in Thailand [48.4% 38 ; 44.3% 23 ].
The high frequency of ESBLP E. coli supports the suggestion that amoxicillin may select for these strains in pig farms and is consistent with the results of a previous study. 7 While no ESBLP E. coli were found in the farms that did not use NA, it was unclear what resulted in ESBLP also being present in the two farms that only used therapeutic enrofloxacin or gentamicin by injection. Infrequent use of these antimicrobials for sick pigs would not be predicted to select for ESBLP E. coli. The strains are unlikely to have been acquired from other PA farms, as they would have been expected to be resistant to a greater range of antimicrobials than was found.
Over 50% of the E. coli from the PA farms showed ESBL properties, commonly possessing blaCTX-M-1 group or blaCTX-M-9 group genes that have a worldwide distribution in both healthy and ill humans in Southeast and East Asia.39–41 By contrast, only a few ESBLP E. coli were detected in the TA farms, and none in the farms that did not use antimicrobials. Previously, ESBL phenotypes were reported in 2.4–75% of porcine E. coli, but in those studies, there was lack of historical data, especially relating to antimicrobial use.25,26,39 This study demonstrated a relationship between the use of prophylactic in-feed antimicrobials in pig farms and a high risk of finding ESBLP E. coli. Moreover, all ESBLP E. coli in this study were resistant to enrofloxacin, which has been on the list of World Health Organization (WHO) concerns for monitoring programs. 2
However, as indicated previously, the prevalence of ESBL-related genes like blaTEM-1 variants (blaTEM-25), blaSHV-1 variants (blaSHV-12), and blaVEB-1 should be confirmed by PCR and sequencing. Resistant strains from pigs could represent a potential risk to human health, and this requires a more detailed investigation. The genes blaCTX-M-1 and blaCTX-M-9 that were detected with single FIB and F replicons, and could be transferred in a conjugation assay, had the potential to undergo horizontal transfer to other strains and the associated environment.42,43
Replicons
FIB and F plasmids are narrow-host-range genetic elements, which have been reported worldwide in human and avian Enterobacteriaceae, associated with tetA, blaTEM group, and blaCTX-M group genes.44,45 The FIB and F plasmid-encoded plasmid addiction systems are self-beneficial for their maintenance and spread, both, with and without antibiotic selection pressure. On the other hand, in our study, broad-host-range plasmids N and HI1 contained blaCTX-M-1 and blaCTX-M-9, and these are known to have epidemiological significance in humans and livestock in Europe, Asia, and the United States.46–49 Plasmid N was only detected in isolates from the PA group: this distribution might have arisen from a high fitness cost of the plasmid such that it is less well maintained in an environment lacking antibiotic selective pressure. 50
Some other potential plasmid-mediated resistance genes were not investigated in this study, and these included floR and ampC that previously have only been detected at very low rates in studies in Southeast Asia and East Asia.51,52 Other resistance genes, including plasmid-mediated quinolone resistance genes, which have been associated with resistant strains from human patients in this area, also need to be studied to see if they may have an origin in isolates from pigs.
Summary
In summary, we report the recent MDR status of commensal E. coli in fattening pigs in Thailand in farms with different patterns of antimicrobial usage. The resistance rates among isolates from the farms included in each category of antimicrobial usage were generally very similar for all the drugs tested. Overall, all farms that regularly used prophylactic tiamulin fumarate and amoxicillin had significantly higher rates of AMR to many antimicrobial drugs than did farms only using targeted injectable TAs, or no antimicrobials at all: this included having a significantly higher risk of having ESBLP E. coli, with the detection of cmlA, catB, aadA1, aadA2, aadB, and dfrA12 genes, and the presence of replicons Y, N, FIC, and HI1.
By inference, increased resistance also was likely to be present in other pathogenic bacterial species in farms routinely using PAs. Overall, these findings indicate that judicious targeted use of antimicrobials for therapy limits the development of AMR in commensal E. coli in pig farms. Somewhat surprisingly, E. coli that were resistant to β-lactams and tetracyclines, with blaTEM and tetA genes, and possessing an F plasmid were common in all Thai pig farms whether or not they used antimicrobials, and so, these particular attributes could not be used to monitor an antimicrobial stewardship policy.
Footnotes
Acknowledgments
This study was supported by the Research and Academic Service, the Faculty of Veterinary Science, H.M. the 90th Year Anniversary Ratchdapisek Somphot Endowment Fund, Chulalongkorn University, STAR; Diagnosis and Monitoring of Animal Pathogen (DMAP), Center of Excellence in Emerging and Re-emerging Infectious Diseases in Animals (EIDAs), Research and Researcher for Industrials Funds, (PHD5610005), and Huevepharma (Thailand) Ltd.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.