
Abstract
The aim of this study was to determine the expression of eight other functional transporter genes upon acrAB inactivation and also the expression of acrAB when the function of eight other transporters are impaired in Salmonella enterica. We used single- or multigene deletion mutants (i.e., ΔacrA, ΔacrB, ΔtolC, ΔacrAB, ΔacrEF, ΔacrD, ΔmdsABC, ΔmdtABC, ΔemrAB, ΔmacAB, ΔmdfA, ΔmdtK, ΔacrABramA, ΔacrABmarA, and ΔacrABsoxS) and real time (RT)-PCR to quantify the expression of different pump and regulator genes; infection ability was characterized by adhesion and invasion assays. The expression of acrAB operon was increased upon acrB inactivation. Single deletion of acrA or tolC also increased expression of acrB. The deletion of acrAB increased expression of eight other functional efflux pumps genes and vice versa, in which increased expression of ramA and marA was also detected. Mutants containing single deletions of functional pump genes were attenuated in cells. In conclusion, there is a feedback mechanism that coordinates regulation of AcrAB-TolC and eight other functional efflux pumps through the global transcriptional regulators ramA and marA in S. enterica serovar Typhimurium.
Introduction
F
The tripartite protein complex AcrAB-TolC is encoded by two distinct operons, acrAB and tolC-ygiAB. Expression of acrAB is controlled by multiple regulators. AcrR represses the expression of acrAB locally. At a global level, acrAB expression is regulated by stressful conditions and by global transcriptional regulators, such as MarA and SoxS.10,11 The acrAB locus is a member of the mar, sox, and rob regulons of E. coli, and it meditates low-level resistance to antibiotics once activated. Ruiz and Levy demonstrated that when the AcrAB-TolC pump in Escherichia coli is inactivated or inhibited, cellular metabolites accumulate, inactivating AcrR and/or upregulating soxS and marA expression. Ultimately, acrAB expression is upregulated to restore homeostasis. 12 S. enterica serovar Typhimurium and E. coli share similarity in both the structure and function in marRAB regulon 13 ; they also have close homology in soxRS regulon. 14 However, a study from Ricci et al. suggested neither marA nor soxS are crucial for S. enterica serovar Typhimurium to generate multidrug-resistant mutants. 15 RamA, a homolog of MarA that is absent in E. coli, has been shown to be involved in MDR in S. enterica. The overproduction of RamA increases expression of the efflux pump AcrAB.16,17 No spontaneous mutants could be selected under ciprofloxacin pressure when ramA was inactivated; this confirmed that ramA plays an important role in MDR development. 15 Some RND efflux pumps also have an effect on the adhesion and invasion abilities of Salmonella in addition to their role in drug efflux.18,19 In S. enterica serovar Typhimurium, acrB inactivated mutants have a decreased ability to invade tissue cells in vitro20,21; furthermore, mutants lacking acrAB or acrEF are attenuated in mice. 22
Besides the RND family (i.e., AcrAB, AcrEF, AcrD, MdsAB, and MdtABC) pumps, there are four other functional drug efflux transporters in S. enterica serovar Typhimurium, 23 belonging to three families defined by sequence similarity: the major facilitator superfamily (MFS) (i.e., EmrAB and MdfA), the multidrug and toxic compound extrusion family (i.e., MdtK), and the ATP binding cassette (ABC) family (i.e., MacAB). 23 Because of the functional redundancy between RND efflux pumps, increased expression of homologous pumps will in a way compensate for the loss of function of certain inactivated pump genes. Eaves et al. detected significantly increased expression of acrD upon inactivation of acrB or acrF in Salmonella (3.6- and 4.9-fold respectively) and these findings suggest a coordinated regulation of acrB, acrD, and acrF; their expression also has an influence on marA and soxS expression. 24 Blair et al. showed increased expression of all RND pump genes upon inactivation of single or multiple acr genes and proposed a feedback mechanism that activates the transcription of homologous RND efflux transporter genes in Salmonella; this compensatory expression was related to increased expression of regulators (e.g., ramA, marA, and soxS). 25 Overproduction of MdtK, EmrAB, and MdfA from other pump families also extrude quinolones, similar to RND efflux pumps. Currently, few studies have investigated the expression of functional pump genes from other families when other pumps are impaired and vice versa.
In this study, we determined whether there is a compensatory function between RND family pumps and efflux pumps from other families; and the impact of inactivation of different pumps on virulence of S. enterica serovar Typhimurium by using a clinical ciprofloxacin-susceptible strain, SH10.
Materials and Methods
Bacterial isolates, plasmids, and growth conditions
S. enterica serovar Typhimurium SH10, was used in this study for the construction of gene deletion mutants (Table 1). The phenotype and genetic characteristics of SH10 were described in the previous study. 26 Bacterial strains were grown overnight in Luria-Bertani broth at 37°C.
Culture medium was commercially obtained from Huankai Microbiology & Technology Ltd. (Guangdong, China). Agar was purchased from Difco (England). RNAiso Plus, Prime Script TM RT Reagent Kit with gDNA Eraser, SYBR® Premix Ex Taq™ (TliRNaseH Plus) were purchased from Takara Biotechnology Ltd. (Japan). Quantitative PCR strip-tubes were obtained from Axygen Ltd. PAβN (Phenylalanine-Arginine Beta-Naphthylamide) was purchased from Sigma-Aldrich Ltd. (America). PAβN is a potent efflux pump inhibitor that targets the RND family pumps and acts as a competitive inhibitor of substrate binding and/or extrusion.
Construction of gene deletion mutants
Single- or multigene deletion mutants were constructed according to Datsenko and Wanner. 27 The kanamycin resistance gene kan, which is flanked by Flp recognition sites, was amplified by PCR (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/mdr). The pKD4 plasmid carrying the kan gene was used as a template. The resulting PCR products were used to transform the recipient SH10 harboring pKD46 plasmid that expressed red recombinase. PCR was used to confirm the mutated chromosomal loci. 27 The kan gene was eliminated using the pCP20 helper plasmid. P22 phage (a kind gift from Laura JV Piddock, the University of Birmingham, United Kingdom) was used for transduction for multiple gene disruption.
RNA extraction and RT-PCR experiments
Bacterial strains were grown in minimal media to an OD600 of 0.6, and total RNA was isolated using the RNAiso Plus Kit. The PrimeScript™ RT Reagent Kit with gDNA Erase (TakaRa, Japan) was used to eliminate genomic DNA and synthesize cDNA. The cDNA was quantified with a iQ5 multicolor real-time PCR system (Bio-Rad, Hercules) using specific primers (Supplementary Table S2) and iQ SYBR Green Supermix (TakaRa, Japan). 16S rRNA was used as a control gene. The relative expression level of each gene was calculated using the ΔΔCt method. 28 Data are presented as mean ± SD. Three independent assays were performed, and each RNA sample was tested in triplicate.
Adhesion and invasion assays
The ability of gene deletion mutants to adhere to and invade INT 407 cells (human embryonic intestine cell line) was assayed using a previously described method. 29 INT 407 cells contain HeLa marker chromosomes, and were derived via HeLa contamination (stored in our laboratory). Each assay was performed at least three times with three technical replicates for each bacterial strain.
Motility assays
Parent strain and deletion mutants were grown in Luria-Bertani broth at 37°C overnight without antibiotic. An agar concentration of 0.3% was used to determine the swimming ability of strains, 10-μl inoculum was stabbed into the center of the plates and the radius of migration was measured after these plates were incubated for 16 hr at 37°C. Each assay was performed at least three times.
Statistical analysis
All experiments were repeated at least three times, and RT-PCR, adhesion, and invasion data were analyzed using one-way ANOVA. p-Values <0.05 were considered statistically significant.
Results
marA and ramA were activated upon inactivation of AcrAB-TolC
The transcriptional level of acrAB was increased upon inhibition or inactivation of AcrAB-TolC (Table 2). RT-PCR data show that the expression of acrA and acrB increased significantly with the addition of PAβN.
Bold fonts indicate statistically significant increases in expression.
When acrA or acrB was inactivated, the expression of acrB or acrA and tolC increased ∼1.92- or 1.76-fold and 1.48-fold (p < 0.05), respectively. The expression of acrA and acrB were also upregulated with tolC inactivation. Furthermore, both marA and ramA were upregulated (∼0.4- to 2- and 1.29- to 2-fold, respectively); on the other hand, soxS expression decreased.
Inactivation of acrAB increases the expression efflux pumps of the RND family and other families
To determine whether the expression of homologous and other family pumps were affected when acrAB was inactivated, we constructed a ΔacrAB mutant and measured the expression level of efflux pumps genes (Fig. 1).

Transcriptional levels of efflux pump genes. *p < 0.05, **p < 0.01.
There was a 1.75- to 2.40-fold increase in the expression of RND family pumps, including acrEF, acrD, mdsABC, and mdtABC, and a 1.59- to 1.66-fold increase in the expression of pumps from other families, including macAB, emrAB, and mdfA. However, mdtK expression did not change (Fig. 1).
We deleted the single transcriptional regulator in ΔacrAB and measured the expression of these efflux pumps. Deletion of ramA decreased expression (0.19- to 0.88-fold) of all pumps except mdtK; deletion of marA decreased (0.26- to 0.85-fold) expression of all pumps except emrA, mdfA, and mdtK, and; the delection of soxS decreased (0.27- to 1.26-fold) expression of all the pumps except mdtK and mdfA (Fig. 1).
acrAB-tolC, ramA, and marA expression increased upon inactivation of other efflux pumps
We constructed eight other efflux transporter deletion mutants (i.e., ΔacrEF, ΔacrD, ΔmdsABC, ΔmdtABC, ΔemrAB, ΔmacAB, ΔmdfA, and ΔmdtK) and measured the expression level of acrAB-tolC, and the transcriptional regulators marA and ramA.
Expression of acrAB-tolC and both transcriptional activator genes increased significantly (Table 3); meanwhile soxS expression decreased significantly and was almost not expressed at all. the mutants (particularly ΔacrAB) were significantly attenuated when efflux transporter genes were inactivated (Fig. 2).
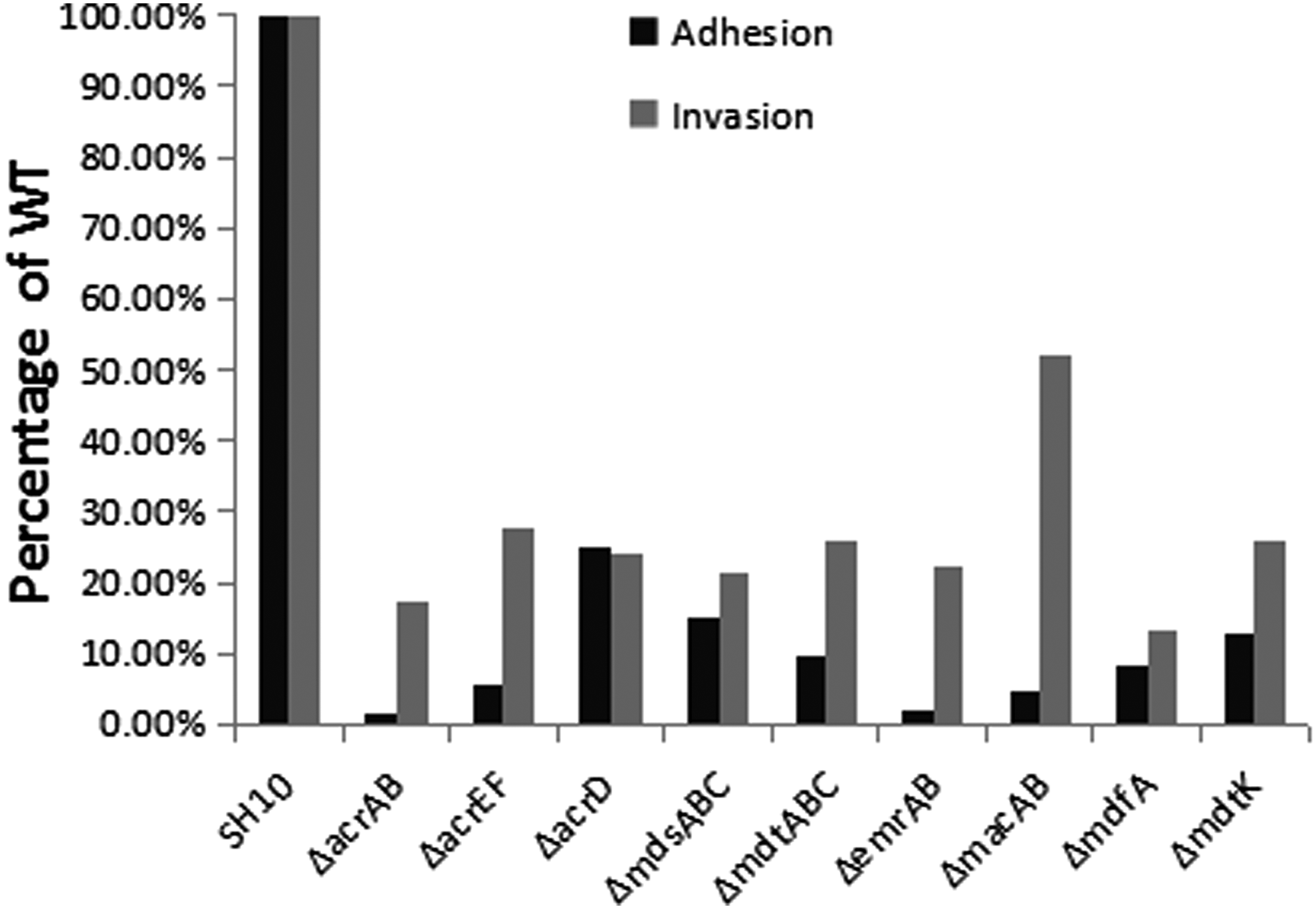
Adhesion and invasion abilities of gene deletion mutants.
Bold fonts indicate statistically significant increases in expression.
Inactivation of functional pump genes affects motility
Deletion of different functional efflux pump genes increased the swimming ability of the mutants when compared with the parent strain, and acrEF, mdsABC, mdtABC, or mdfA deletion mutants increased significantly (p < 0.05) (Fig. 3). Inactivation of soxS, but not of ramA, or marA, prevented the ΔacrAB-mediated increase in swimming ability (Table 4).
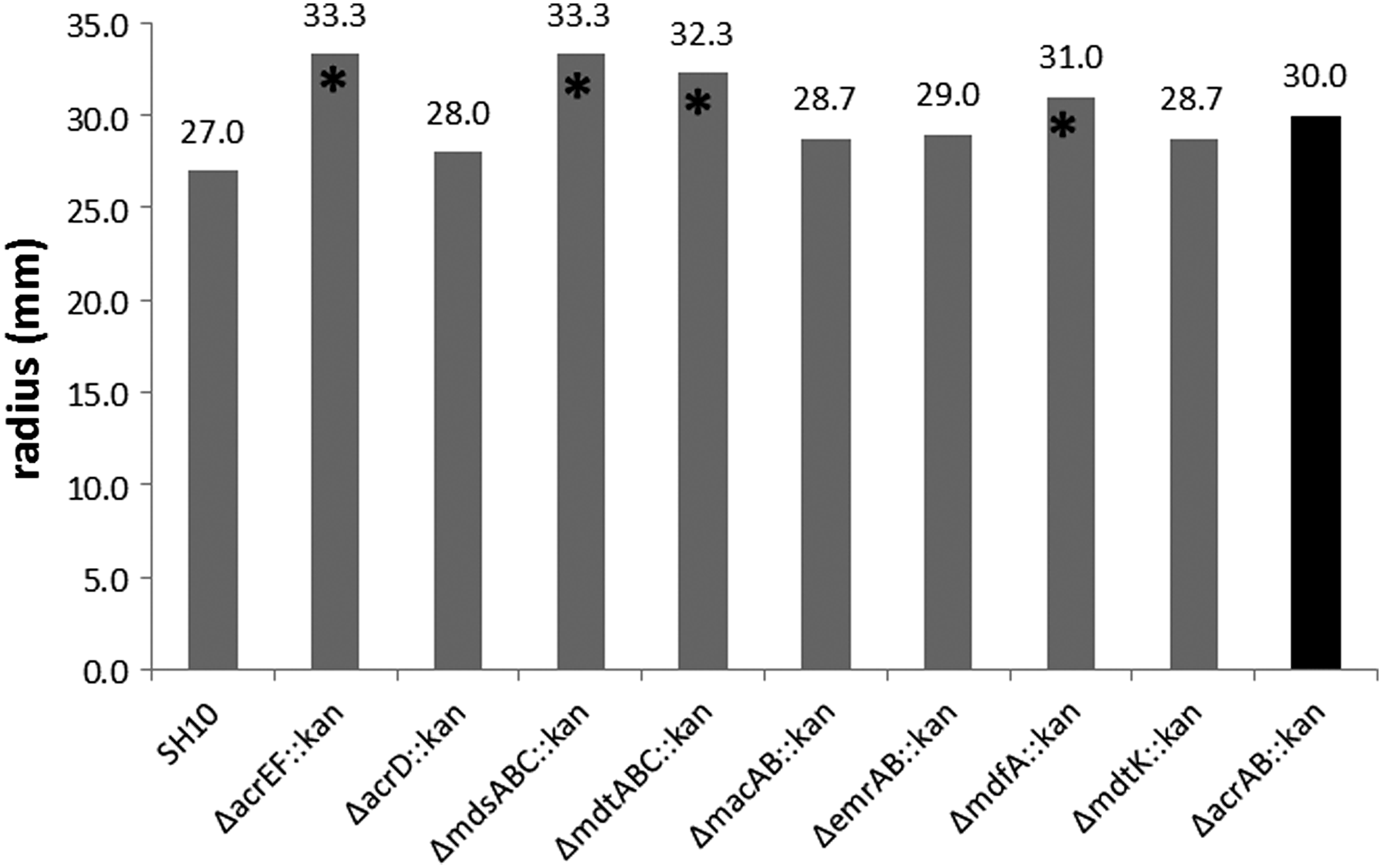
Impact of efflux pump genes deletion on swimming ability of mutants. *p < 0.05.
LB, Luria-Bertani.
Discussion
AcrAB-TolC is the main efflux pump in both S. enterica and E. coli, and the expression of it is modulated by multiple levels. In contrast to E. coli, mar and sox regulons are not critical for efflux-mediated MDR through overproduction of acrAB in Salmonella, ramA is the most important regulator and the overproduction of it results in upregulated acrAB expression in Salmonella and thus leading to MDR. In addition to the role of AcrAB-TolC pump in drug efflux, it also plays a role in virulence of pathogens,20,21 cell metabolism, 30 and regulation of gene expression.12,31
Ruiz and Levy 12 found increased expression of acrAB in E. coli when the efflux pump was inactivated and inhibited; this increase disappeared upon the deletion of transcriptional regulators acrR, soxRS, or marA. They also detected increased expression of soxS and marA, but not acrR. Their results indicated that when AcrAB-TolC is inactivated or inhibited in E. coli, increased expression of soxS and marA triggers the upregulation of acrAB expression to restore homeostasis. 12 AcrAB in Salmonella spp. has an ∼90% amino acid identity similar to AcrAB in E. coli; additionally, the function of AcrAB with the outer membrane channel protein TolC also exists in Salmonella spp. 32 Our results showed that the expression of the transcriptional regulator genes marA and ramA (p < 0.05) in acrA or acrB single deletion or inhibition mutants increased significantly and thus upregulated the expression of the acrAB promoter. This suggests that in S. enterica serovar Typhimurium, there is a positive regulation mechanism triggering the upregulation of the acrAB promoter through increased expression of the transcriptional regulators ramA and marA when acrAB-tolC was inhibited or inactivated. This is the first study to demonstrate that there is a feedback mechanism in S. enterica serovar Typhimurium similar to E. coli.
Five RND family efflux systems have been identified in S. enterica serovar Typhimurium, including AcrAB, AcrD, AcrEF, MdtABC, and the S. enterica specific MdsABC.33,34 The efflux pump AcrB and its homologs are the most clinically relevant RND pumps since they have the broadest spectrum of substrates. 35 ramA, marA, and soxS regulate the expression of arcAB globally.10,11 Blair et al. 25 found that the expression of all RND family pump genes was increased following single or multiple inactivation of the acr gene. The authors then proposed a feedback mechanism by which transcription of other RND homologous efflux transporter genes are activated in Salmonella. 25 We also investigated that the inactivation of acrAB results in increased expression of the pump genes acrF, acrD, mdsB, mdtB, macA, emrA, and mdfA, but not mdtK. Further deletion of ramA in ΔacrAB led to a reduction in the expression of all tested pump genes with the exception of mdtK; further deletion of marA caused decreased expression of pump genes with the exception of emrA, mdfA, and mdtK. Meanwhile the expression of acrAB-tolC and both transcriptional regulator genes (i.e., ramA and marA), but not soxS, significantly increased in eight other single functional efflux pump gene deletion mutants. Our data demonstrate a compensatory regulation mechanism whereby the inactivation of acrAB activates the expression of homologous RND family efflux pump genes, and other functional efflux pump genes belonging to the MFS (emrAB and mdfA) and ABC (macAB) families through coordinated upregulation of the global transcriptional regulator genes ramA and marA.
RND efflux pumps also have an effect on the adhesion and invasion abilities of Salmonella.18,19 Inactivation of acrB in S. enterica serovar Typhimurium leads to decreased invasion abilities in tissue cells in vitro,20,21 and S. enterica serovar Typhimurium lacking acrAB or acrEF was attenuated in mice. 22 Studies from Piddock and colleagues showed no impact on virulence of INT 407 cells upon inactivation of acrD 21 and Nishino et al. found minimal impact on BALB/c mouse survival once acrD inactivated 23 ; however, another study detected attenuated adhesion and invasion of INT 407 cells upon acrD inactivation. 25 Data from our study indicate that inactivation of nine functional efflux pump genes will also reduce the adhesion and invasion abilities of S. enterica serovar Typhimurium to INT 407 cells.
We show that single inhibition or inactivation of AcrAB-TolC stimulates the expression of the transcriptional regulators ramA and marA, resulting in upregulation of the acrAB promoter in S. enterica. We also show that the inactivation of acrAB leads to upregulation of acrEF, acrD, mdsABC, mdtABC, macAB, emrAB, and mdfA expression through the regulators ramA and marA. Finally, we show that the upregulation of acrAB following inactivation of acrEF, acrD, mdsABC, mdtABC, macAB, emrAB, mdfA, or mdtK is also controlled by the expression of ramA and marA.
In conclusion, our study demonstrate a feedback mechanism between nine functional efflux pump genes through co-regulation of ramA and marA in S. enterica serovar Typhimurium and single deletion of functional efflux pump genes attenuated virulence of mutants.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (31272602) and the Program for Changjiang Scholars and Innovative Research Team in University of Ministry of Education of China (Grant No. IRT13063).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.