
Abstract
The aim of this study was to compare the antimicrobial resistance profiles of top five enterohemorrhagic Escherichia coli (EHEC) and enteropathogenic E. coli (EPEC) to E. coli isolated from the fecal flora of the same adult cattle. Previous prevalence studies had led to the isolation by immunomagnetic separation (IMS) of 39 EHEC and 80 EPEC. Seven EHEC were resistant (17.9%), and six were multidrug resistant (MDR) (15.4%). None of the top five EHEC was resistant to azithromycin. Nine EPEC O26:H11 (11.3%) were resistant. They were all resistant to tetracycline, and four were MDR (5.0%). An E. coli strain was isolated from the feces (without preselection by IMS) of 97 bovine carriers of top 5 strains. All these strains were susceptible to antibiotics. Comparative analyses did not reveal any differences between the cytotoxic activities of resistant EHEC and their susceptible counterparts or in the production of attachment and effacement lesions. These results highlighted the higher percentage of resistance of EHEC and EPEC strains compared to other E. coli. They also showed that resistance traits did not have any impact on the expression of virulence phenotypes in EHEC strains.
Introduction
E
Cattle are considered reservoirs of EHEC that can be transmitted to humans through the ingestion of contaminated foodstuff and water. 5 In France, the prevalence of top five EHEC was estimated to 1.8% in slaughtered adult cattle. 6 As described in other prevalence studies, screening of cattle for the presence of EHEC also led to the isolation of enteropathogenic E. coli (EPEC), which were stx negative. They belonged to top five serotypes and carried the associated eae subtypes. 7 They were classified as atypical EPEC (aEPEC) based on the absence of the bfpA gene and the EAF plasmid. 8 aEPEC have been isolated from various animal species. 9 The ability of aEPEC to cause illness in humans is still controversial. 10 Nevertheless, it has been suggested that these particular aEPEC sharing similar virulence genes with EHEC might be either progenitors of EHEC 11 or derivates of EHEC which have lost their stx genes either in cattle's gut or during isolation procedure. Indeed, loss of Stx-encoding phages has been shown to occur both in vitro and in vivo.12,13
The usefulness of antimicrobial therapy for EHEC infections remains controversial, as antibiotic treatment can lead to an increased risk of developing HUS. 14 Nevertheless, a recent review provided criteria for a possible antibiotic regimen for EHEC-infected patients. 15 Moreover, the use of azithromycin during the outbreak of Shiga toxin–producing enteroaggregative E. coli O104:H4 in Germany led to a decreased carriage of the epidemic strain in patients. 16 The antibiotic resistance of EHEC strains is therefore a matter of concern. Although several studies have reported the isolation of antibiotic resistant EHEC from cattle and humans,17–19 only a few studies have explored the relationship between antibiotic resistance and virulence traits in this pathotype.20–22 It should be stressed that in these epidemiological studies the frequency of resistance was compared in pathogenic and nonpathogenic E. coli originating from different individuals. As far as we know, no study has investigated, in the same animal, the difference in resistance profile between EHEC strains and E. coli strains, which might have been submitted to the same antibiotic pressure. Such studies might help to investigate whether antibiotic selection pressure or genetic association contribute to the emergence of antibiotic resistance in EHEC.
Therefore, the objective of this study was to perform a comparative analysis of the antimicrobial resistance profiles of EHEC and aEPEC belonging to the top five serotypes versus other E. coli isolated from the fecal flora of the same adult cattle. Moreover, the relationship between antimicrobial resistance and virulence traits was explored by evaluating the expression of virulence phenotypes in resistant EHEC strains and their susceptible counterparts.
Materials and Methods
Collections of EHEC, EPEC, and E. coli
A previous prevalence study, conducted in French slaughterhouses, had resulted in the isolation from adult cattle of 39 EHEC of serotype O157:H7, O26:H11, O103:H2, O111:H8, O145:H28, and 80 EPEC of the same serotypes, but lacking the gene stx. 6 These strains had been isolated by immunomagnetic separation (IMS) after enrichment of feces. Table 1 gives the distribution per serotype. These 119 strains had been isolated from 109 fecal samples, as some cattle carried 2 top 5 strains. The 109 bovine came from 86 distinct farms. In the present study, an attempt was made to isolate at least one E. coli strain, without genetic characteristics of top five strains, from each bovine carrier. To this end, a fecal sample that had not been subjected to enrichment and had been stored at −20°C was 10-fold diluted. This suspension was plated onto RAPID'E. coli 2 agar plates (Bio-Rad, Marnes-La-Coquette, France), which were then incubated for 18–24 hours at 42°C. After incubation, about eight well isolated colonies from each fecal sample were picked at random. Colonies were grown at 37°C for 24 hours in 96-well microplates containing LB broth. The identification of E. coli was confirmed by indole test, and confirmed E. coli were stored at −80°C after addition of 30% glycerol until analysis.
EHEC, enterohemorrhagic E. coli; EPEC, enteropathogenic E. coli.
At least one E. coli strain per bovine was investigated for the presence of the EHEC–associated genetic markers stx1, stx2, eae, and ehxA by polymerase chain reaction (PCR) as previously published. 23 When antibiotic resistance was detected in EHEC and aEPEC, up to four other E. coli isolates from the same animal were tested. E. coli that tested positive for stx and eae positive were further characterized. Subtyping of the stx1 and stx2 genes was performed as described by Scheutz et al. 24 For eae-positive E. coli isolates, the presence of subtypes γ, ß, ɛ, and θ was screened by real-time PCR. 25 The presence of top five serotypes (O157:H7, O26:H11, O145:H28, O111:H8, and O103:H2) was also investigated by real-time PCR.25,26 Finally, E. coli that have shown to harbor genetic characteristics of top five strains were excluded from the comparative analysis of antimicrobial resistance profiles of top five strains versus other E. coli.
Antimicrobial susceptibility testing and characterization of resistance genes
Antimicrobial susceptibility testing was performed by disk diffusion method. The antibiotic disks (Bio-Rad) used in this study were as follows: ampicillin, amoxicillin plus clavulanic acid, cefalexin, cefuroxime, cefotaxime, ceftazidime, cefepime, ertapenem, gentamicin, streptomycin, tetracycline, chloramphenicol, trimethoprim, sulfonamides, nalidixic acid, and ciprofloxacin. Antimicrobial susceptibility tests were performed and interpreted according to the guidelines of the French Society for Microbiology (SFM) and the European Committee on Antimicrobial Susceptibility Testing (EUCAST). 27 Isolates were considered as multidrug resistant (MDR) when they exhibited resistance to at least three antibiotic classes. 28 Resistant isolates were screened for the main resistance genes: (blaTEM), (strA-strB, aadA1), [tet(A), tet(B)], (sulI, sulII, sulIII), (cmlA, catI, catII, catIII, floR), respectively, using primers and control strains as previously described. 29 EHEC and EPEC were screened for the presence of intI1, intI2, and intI3 genes using a triplex real-time PCR. 30 Finally, the azithromycin minimum inhibitory concentrations for the 39 EHEC were determined by E-test (BioMérieux, Marcy l'Etoile, France) as recommended by the manufacturer.
Vero cell cytotoxicity assay
Vero cells were seeded at 5.103 Vero cells per well on 96-well microplates and cultivated for 24 hours in Dulbecco's Minimum Essential Medium containing nonessential amino acids (DMEM 1X; Gibco, Life Technologies), supplemented with 10% fetal calf serum (FCS; Gibco) at 37°C in a 5% CO2 atmosphere. Bacteria were cultured in LB broth at 37°C overnight and then subcultured in LB broth supplemented with 200 mM of mitomycin C (to increase the production of Stx) at 37°C for 16 hours. A volume of 2 mL of bacterial culture containing 109 colony forming unit/mL adjusted with LB broth was centrifuged at 6,000 g for 5 minutes at 4°C. The supernatant was filtered through a 0.22 μm pore-size membrane filter (Millipore, Carrigtwohill, Ireland) and serially diluted (1:4) with Dulbecco's phosphate-buffered saline (PBS; Sigma-Aldrich, St Louis, MO).
The interaction was performed by adding 10 μL of each dilution to each well containing Vero cells. After 72 hours of interaction at 37°C, the cells were washed thrice with Hank's Balanced Salt Solution (HBSS; Sigma-Aldrich), fixed with 3.7% paraformaldehyde for 15 minutes, and then washed with 100 μL of PBS thrice and 100 μL of 0.01 M Tris-HCl, pH 8.5. The fixed cells were then stained with 100 μL of 1% methylene blue in Tris-HCl for 45 minutes, washed again with 100 μL distilled water and 100 μL of Tris-HCl, and finally dried overnight. The methylene blue was extracted with 100 μL of 0.1 M HCl by shaking. After 15 minutes of extraction, 75 μL of cells was transferred to new 96-well microplates, and the color density of each well was measured by TECAN microplate reader (Infinite M200) at 660 nm. The E. coli strains EDL933 O157:H7 (stx1a, stx2c, eae-γ) and DH5α were used as positive and negative controls, respectively. All assays were repeated twice independently.
Fluorescent-actin staining test
Bacteria were inoculated in LB broth and incubated at 37°C overnight. HeLa cells were seeded at 3.7 × 104 cells per well on Lab-Tek 8 Chamber Slides (Nunc) and incubated for 24 hours in the same conditions as those of Vero cells. HeLa cells were washed thrice with preheated HBSS (37°C). The interaction was done in 500 μL of DMEM buffered with 25 mM of HEPES complemented with 5% FCS, with a starting inoculum of 102 of bacteria per HeLa cell. After 6 hours of interaction at 37°C, the cells were washed six times with HBSS, fixed with 4% paraformaldehyde in PBS for 20 minutes at room temperature, and then permeabilized with 0.25% Triton X-100 in PBS for 5 minutes. Fluorescent-actin was labeled with Rhodamine-phalloidin (Molecular Probes) in accordance with the manufacturer's instructions. EPEC E2348/69 O127:H6 and DH5α were used as positive and negative controls, respectively.
Results
Isolation of E. coli from bovine carriers
In a previous prevalence study conducted in French slaughterhouses, 39 EHEC and 80 EPEC had been isolated from the feces of adult cattle. 6 Fecal samples (104) from these 109 bovine carriers had been stored, and an E. coli could be isolated from 103 samples. Screening of EHEC–associated genetic markers showed that 14 E. coli, isolated without enrichment, were positive for at least 1 of the tested markers (Table 2). For eight bovine carriers, E. coli strains isolated without enrichment were positive for only one EHEC–associated genetic marker (Table 2). Five strains were positive for stx1d or stx2c genes, and three strains were positive for the eae gene. The subtype eae-θ was identified in two strains.
The presence of top five serotypes was investigated; n.d., not determined.
n.d., eae subtype not determined.
IMS, immunomagnetic separation; S, susceptible; TET, tetracycline.
For the other six bovine carriers, these E. coli strains had the genetic characteristics of top five EHEC and EPEC. Moreover, for five of these six cattle (A126, A75, C24, I77, and J25), the EHEC or EPEC strains isolated with or without enrichment displayed the same genetic characteristics and resistance phenotype. This suggested that they might belong to the same clone. For the J28 bovine, the top five strains isolated with or without enrichment were different. An EHEC O157:H7 had been previously isolated by IMS after enrichment. The attempt to isolate an E. coli without enrichment led to the isolation of EPEC O26:H11 from the same bovine. These six bovine carriers were therefore excluded from the comparative analysis of antimicrobial resistance profiles of EHEC and/or EPEC versus other E. coli.
Comparative analysis of antibiotic resistance of EHEC, EPEC, and E. coli
All 97 E. coli strains, which were isolated without enrichment, were susceptible to the tested antibiotics. On the contrary, 16 (13.4%) of the 119 EHEC and EPEC strains showed resistance to at least 1 of the tested antibiotics (Fig. 1). Among the 39 EHEC tested, 7 (17.9%) were resistant to at least 1 antibiotic (Table 3). They belonged to serotypes O157:H7, O26:H11, O103:H2, and O111:H8. Six were MDR, and the observed resistance was mainly to ampicillin, streptomycin, tetracycline, and sulfonamide. One strain was resistant to nalidixic acid. Moreover, all 39 EHEC strains were susceptible to azithromycin. With regard to the resistance of EPEC strains, 9 (11.3%) of the 80 strains were resistant (Fig. 1). All these resistant strains belonged to the serotype O26:H11 (Table 3). They were all resistant to tetracycline, and four were MDR. The main antibiotic resistance genes involved in the resistance phenotypes of EHEC and EPEC were blaTEM, strA-strB, tet(A), and sulII. Moreover, only one strain was positive for the integrase gene intI1 (strain H52-O26-1). Finally, for each cattle carrying a resistant EHEC and/or EPEC strain, up to four other E. coli isolates were isolated and tested (Table 3). They were all susceptible.
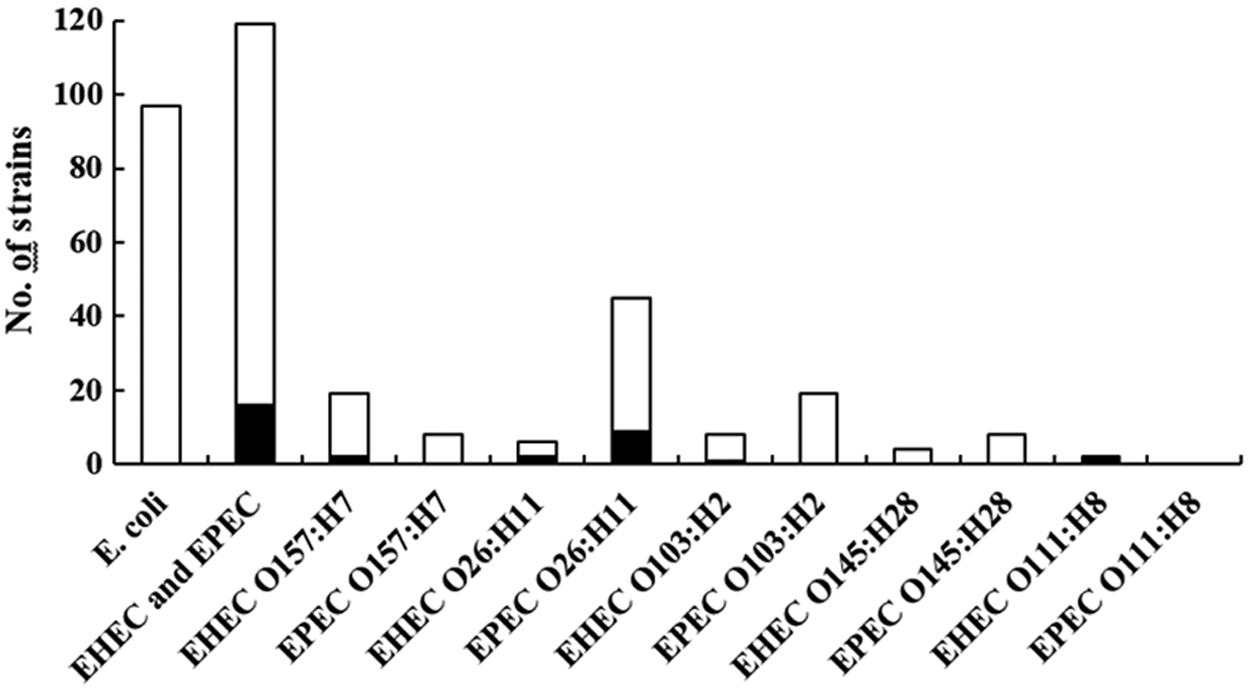
Number of tested and resistant Escherichia coli, EHEC and EPEC. Filled bars indicate the number of antibiotic-resistant strains. EHEC, enterohemorrhagic E. coli; EPEC, enteropathogenic E. coli.
The following antibiotic resistance genes were screened: blaTEM, strA-strB, aadA1, tet(A), tet(B), sulI, sulII, sulIII, cmlA, catI, catII, catIII, and floR.
n.a., as the EPEC and the E. coli strain, isolated without preselection by IMS, were shown to possibly belong to the same clone.
AMC, amoxicillin plus clavulanic acid; AMP, ampicillin; CHL, chloramphenicol; n.a., not applicable; NAL, nalidixic acid; n.d., not determined; SSS, sulfonamides; STR, streptomycin; TET, tetracycline.
Results of cytotoxicity and fluorescent-actin staining tests for resistant and susceptible EHEC
The cytotoxic activity and adhesion to HeLa cells of EHEC strains were investigated in seven resistant EHEC and six susceptible counterparts. Six of the resistant EHEC displayed toxicity to Vero cells, and five of the susceptible EHEC strains tested displayed toxicity to Vero cells (Fig. 2). The resistant strain H13-O157-1 (O157:H7, eae-γ1, stx2c) and the susceptible strain K143-O157-1 (O157:H7, eae-γ1, stx2c) had no detectable cytotoxic effect compared to the negative control strain E. coli DH5α. The ability of these EHEC strains to produce attachment and effacement lesions (A/E) on cultured HeLa cells was also determined by the fluorescent-actin staining test. The test was positive for resistant and susceptible strains, as an accumulation of filamentous actin was identified at the sites of adhesion (data not shown).
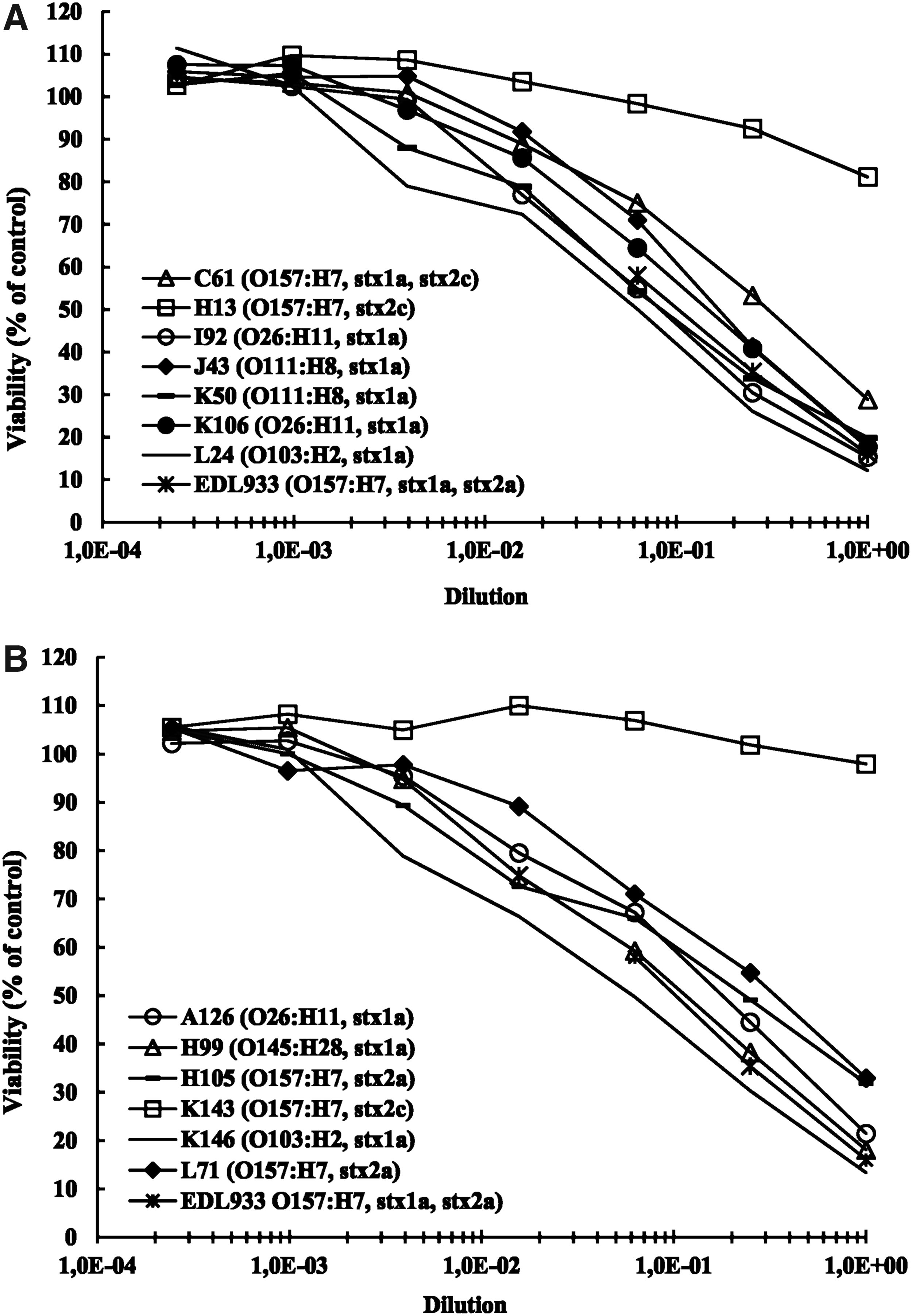
Cytotoxic activity of seven resistant EHEC strains
Discussion
EHEC (39) and EPEC (80) of serotypes O157:H7, O26:H11, O111:H8, O103:H2, and O145:H28 had been previously isolated in a prevalence study conducted in French slaughterhouses. 6 Our study showed that 9 (11.3%) of the 80 EPEC strains were resistant, although this percentage is nevertheless low in comparison to the results reported in other prevalence studies. 31 Regarding the 39 EHEC tested, 7 showed resistance to at least one of the tested antibiotics (17.9%); and 6 were MDR (15.4%). Similarly, only a few other studies reported low prevalences of resistance in EHEC strains.32,33 In contrast, high degrees of resistance had been reported in EHEC O157:H7 and other top five EHEC originating from cattle (O26:H11; O111:H8; O103:H2).19,20,34,35 The reasons for these differences remain unclear and require further investigation. Finally, it should be stressed that all the EHEC tested in our study were susceptible to azithromycin. A recent study conducted in France also demonstrated that, except for two atypical strains, 508 EHEC clinical strains were susceptible to azithromycin. 36
The possible interplay between antibiotic resistance and virulence in EHEC was investigated. Indeed, some studies intended to document the impact of resistance traits on pathogenicity for other pathovars.37–39 Given that antibiotic resistance should have a fitness cost, it might be associated with a reduction of bacterial virulence. In this study, resistant determinants detected in EHEC were blaTEM, strA-strB, tet(A), and sulII. These genes are usually carried on transmissible genetic elements, such as transposons and plasmids, which might be associated with biological costs.40,41 The investigation of a possible link between resistance and virulence is a complex topic.37–39 Observation of an impact, or not, seemed to depend on the resistance and virulence genes and the bacterial species studied. In this study, we investigated the ability of the seven resistant EHEC to cause illness in humans by examining in vitro the cytotoxic activity to Vero cells and the adhesion to HeLa cells. We also tested whether the presence of resistance genes affected the expression of the virulence phenotypes by performing the same investigation on six susceptible counterpart EHEC strains.
All EHEC strains showed ability to produce attachment and effacement lesions. They also displayed toxicity to Vero cells, with the exception of one resistant and one susceptible EHEC O157:H7 strain. Both these strains were positive for the stx2c gene, in agreement with previously published data. 42 Indeed, Shiga toxin 2c (Stx2c) was shown to be at least 25 times less potent than Stx2a to induce toxicity to Vero cells. 43 These various data suggested that the expression of virulence phenotypes was not impacted by antibiotic resistance carriage in EHEC strains. We demonstrated in this study that presence of the usual resistance genes in EHEC strains had no impact on their cytotoxic activity and their ability to produce attachment and effacement lesions in HeLa cells.
We then explored the relationship between antibiotic resistance and virulence in EHEC and EPEC strains by performing a comparative analysis of the antimicrobial profiles of EHEC and EPEC versus other E. coli strains isolated from the same cattle. The first step was to isolate E. coli strains from feces that had not been subjected to enrichment and to check that these strains were devoid of the characteristics of EHEC and EPEC strains. The EHEC and/or EPEC and the E. coli strains isolated without enrichment were shown to possibly belong to the same clone for only five bovine carriers. This is in agreement with the fact that top five EHEC and EPEC strains are usually present at low concentrations in feces, thus explaining the need for enrichment before their isolation. 44
All E. coli strains were susceptible to the tested antibiotics. We had previously shown that wastewater from a French adult cattle slaughterhouse only contained 5% of resistant E. coli. 45 Similarly, indicator E. coli isolated from adult cattle in Austria were shown to exhibit low percentages of resistance to ampicillin and chloramphenicol (from 1.4% to 2.3%). In meat-producing cattle from Spain and Germany, only 0.6–19.8% of indicator E. coli were resistant to one of the antibiotics tested. 46
The percentage of resistance was higher for EHEC and EPEC than for the other E. coli (13.4% vs. 0%), which suggests a correlation between virulence and antibiotic resistance traits in such strains. The question of a relationship between virulence and resistance traits in intestinal pathotypes has been addressed in only a few studies. Cabal et al. compared the resistance of E. coli EHEC strains O157:H7 versus non-O157:H7 E. coli isolated from cattle feces. These authors reported a significantly higher proportion of resistant isolates among EHEC O157:H7 isolates than in non-O157:H7 E. coli strains for most of the antibiotics tested. 20 In another study conducted on healthy pigs, a positive statistical association between resistance and various virulence factors, including stx1, stx2, eae, estA, estB, elt, aidA-1, and fedA1, was demonstrated in E. coli isolated from feces. 21 Recently, a significantly higher prevalence of antibiotic resistance for diarrheagenic E. coli isolates than for commensal E. coli was highlighted in an epidemiological study of populations living in rural Ecuador. 22 We also showed in this study that the percentage of resistance was higher for pathogenic isolates than for commensal isolates and hypothesized that the higher percentage of resistant strains observed in pathogenic strains might be due to selective pressure and/or genetic linkage between virulence and resistance genes.
It must be stressed that our comparative study was performed on E. coli originating from the same animal, whereas the previously cited investigations were epidemiological studies in which the frequency of resistance was compared in E. coli isolates originating from different animals or humans. In our study, the EHEC, EPEC, and other E. coli would probably have been subjected to the same antibiotic pressure in the bovine's digestive tract. If the level of resistance was driven by antibiotic pressure, the same level of resistance would have been observed in both populations. Consequently, our data imply that genetic linkage between virulence and resistance genes rather than selection pressure would be involved in the co-occurrence of virulence and resistance. Only a few studies investigated the localization of antibiotic resistance genes in top five EHEC and aEPEC. Venturini et al. highlighted a co-location of resistance and virulence genes on an EHEC virulence plasmid.47,48
Plasmid pO26-CRL111, extracted from an EHEC O26:H- isolated from a patient with hemorrhagic colitis, has been reported to carry concomitantly genes encoding resistance to six classes of antibiotics and several virulence genes, including ehxA.47,48 Resistance genes were carried by a derivate Tn21 transposon. This derivate Tn21 transposon was also identified in two nearly identical plasmids, which carried both putative and established E. coli virulence genes (1) pO26-CRL125, from the same EHEC O26:H- strain, and (2) pO111-CRL115, from a bovine O111 aEPEC. 48
Conclusions
We showed that adult cattle slaughtered in France can be reservoirs of resistant EHEC and EPEC strains of serotypes O157:H7, O26:H11, O103:H2, O111:H8, O145:H28. However, the frequencies of resistance were generally low, compared to other studies. The percentage of resistance was higher in EHEC and EPEC strains than in other E. coli isolated from the same adult cattle. This report raises concern about the possible risk of expansion of such pathogenic strains, selected notably by antibiotic selection pressure within the bovine carriers.
Footnotes
Acknowledgment
This work was supported by funds from the French National Research Agency (ANR), program “Eco-Tech,” study “Défi-Viande” (ANR-09-ECOT-00603).
Disclosure Statement
No competing financial interests exist.