
Abstract
Acinetobacter baumannii is an important nosocomial pathogen which causes a wide range of infections. In this study, we addressed the role of class 1 integron, ISAba1 and ISAba125 associated with antimicrobial resistance in 72 clinical isolates of A. baumannii collected from clinical settings in Tehran, Iran. Moreover, to study the clonal relatedness of strains, repetitive extragenic palindromic-PCR (rep-PCR) assay was carried out. PCR revealed that blaOXA-51-like, blaOXA-23-like, blaOXA-24/40-like, blaOXA-58-like, blaNDM, integrase gene (intI1), ISAba1, and ISAba125 were present in 86.11% (62/72), 84.72% (61/72), 30.55% (22/72), 0% (0/72), 0% (0/72), 58.33% (42/72), 97.22% (70/72), and 65.27% (47/72) of the strains, respectively. Sequencing of 39 internal variable regions of class 1 integrons showed seven gene cassette arrays, including aadA4-catB8-aadA1 (2.77%), aadA1-aadA4 (1.38%), aacC4-aadA1 (2.77%), aacC4 (22.22%), aadA1 (13.88%), aadA4 (5.55%), and catB8 (5.55%). We detected ISAba1 in the upstream of blaOXA-23-like, blaOXA-51-like, and blaADC in 54.16% (39/72), 9.72% (7/72), and 56.94% (41/72) of the strains, respectively. Whereas, there was a low frequency of disruptions in carO and dacD genes: 5.55% (4/72) and 4.16% (3/72). Rep-PCR analysis revealed that the isolates were genetically diverse. However, Cl-12 and Cl-15 were the largest clusters and they were recovered from various hospitals. Our analysis showed the high rates of class 1 integrons as a repertoire of aminoglycoside-modifying enzymes. It seems that linkages of ISAba1-blaOXA-23-like and ISAba1-blaOXA-69, and disruptions in carO or dacD can develop resistance to carbapenems among clinical isolates of A. baumannii.
Introduction
A
Class 1 integrons are versatile gene acquisition systems that have accumulated various number of resistance genes from the environmental pool of these elements. 4 They have a major role in the dissemination of antibiotics, disinfectants, and heavy metal resistance genes. 5
On the other hand, insertion sequences (ISs) are the smallest and the most abundant transposable elements (<2.5 kb) with the capability of independent transposition in microbial genomes. Several families of IS have been associated with antimicrobial resistance.6,7 They can be associated with antibiotics resistance through two mechanisms: (i) transposition of IS elements in the upstream of resistance gene that effects its expression through nucleotide changes in the promoter or (ii) causing inactivation of a gene by disrupting it. 8 These mechanisms increase resistance to antibiotics.
For example, the expression of blaADC is enhanced by the entrance of the ISAba1 or ISAba125 in upstream of the gene providing a strong promoter for gene expression, resulting in resistance to ceftazidime (CAZ). 9 Moreover, ISAba1 has been found in the upstream of blaOXA-51-like, blaOXA-23-like, and blaOXA-58-like genes. 6 Also, ISAba125 has been detected in the upstream of blaNDM-1 & 2. 6 Overexpression of AdeABC, as resistance–nodulation–division efflux pumps, is caused by mutations in the adeRS genes encoding a two-component regulatory system, which confer resistance to numerous antibiotics in A. baumannii. 10 A previous research has shown that a disrupted AdeS kinase protein generated by ISAba1 insertion correlates with tigecycline resistance in A. baumannii. 11
Previous studies have described the disruptions of carO gene by ISAba1, ISAba10, ISAba825, or ISAba15.8,12–14 Disruption of carO blocks the uptake of antibiotics, including imipenem (IMP) and increases the minimum inhibitory concentration (MIC) to this carbapenem. Moreover, ISAba125, which disrupts the dacD gene encoding a penicillin-binding protein, has been identified in an endemic carbapenem-resistant clone. 15 The objective of the present study was to determine the diversity of class 1 integrons, and disruption of carO and dacD by ISs among A. baumannii isolates in Tehran, Iran.
Materials and Methods
Bacterial isolates and antimicrobial susceptibility testing
The isolates were selected from a collection of consecutive nonduplicate clinical isolates of A. baumannii obtained from inpatients in six general hospitals in Tehran, Iran. In this study, 72 isolates which were nonsusceptible to ≥1 agent in ≥3 antimicrobial categories, were considered for further investigation. The isolates were recognized phenotypically by analytical profile index 20NE protocol (BioMérieux, France) as A. baumannii and genetically confirmed by the presence of a chromosomally blaOXA-51 gene, which is intrinsic to this species. 16 Prototype A. baumannii ATCC 19606 was used as a positive control strain. Antimicrobial susceptibility testing was determined by the disk diffusion method according to Clinical and Laboratory Standards Institute (CLSI, 2017) recommendations. The following antibiotic disks purchased from MAST Co., Ltd. (United Kingdom) were used in this study: ciprofloxacin (CIP; 5 μg), (CAZ; 30 μg); gentamicin (GEN; 10 μg), (IMP; 10 μg); minocycline (MIN; 5 μg); and ampicillin/sulbactam (SAM; 10/10 μg). The MICs of IMP and colistin (COL) were determined using broth microdilution and the results were interpreted according to the CLSI guidelines. Escherichia coli ATCC 25922 was used as reference strain in disk diffusion and broth microdilution methods.
PCR detection of class 1 integrons
The genomic DNA was extracted by the DNA Genomic Extraction Kit (Thermo Scientific, Lithuania) according to the manufacturer's instructions. Seventy-two A. baumannii isolates were screened for the presence of class 1 integrons by PCR with primers specific for the intI1 and mapped with primers complementary to conserved segments of class 1 integrons as shown in Table 1. Sequencing of the purified PCR products were performed using DNA Analyzers (Applied Biosystems, Inc.). The nucleotide sequence analysis was carried out by online BLAST tool at NCBI website (https://blast.ncbi.nlm.nih.gov/Blast.cgi). The sequences were then manually assembled by using CLC main workbench software version 5.5 (CLC Bio, Aarhus, Denmark).
PCR detection of blaOXA-51-like, blaOXA-23-like, blaOXA-24/40-like, blaOXA-58-like, and blaNDM genes
All isolates were screened for the presence of blaOXA-51-like, blaOXA-23-like, blaOXA-24/40-like, blaOXA-58-like, and blaNDM genes according to PCR protocol using primers as shown in Table 1. As mentioned in the introduction, these genes are known to be disrupted by ISs ISAba1 and ISAba125. Thus, they were included in the present study.
PCR detection of ISAba1, ISAba125, linkages, and disruptions
PCR assays were performed to detect ISAba1 and ISAba125 in 72 isolates of A. baumannii. To determine the presence of ISAba125 in upstream of the blaADC gene, as well as the presence of ISAba1 elements in upstream of the blaOXA-51-like, blaOXA-23-like, blaOXA-58-like, and blaADC genes, new pairs of primers were designed by AlleleID software (v7.5; Premier Biosoft International, CA). Moreover, PCR was conducted for detection of carO, adeS, and dacD genes. Selected PCR products were subjected to sequencing. The list of primers used in this study has been shown in Table 1.
Repetitive extragenic palindromic-PCR
To study the clonality of A. baumannii isolates, computer assists repetitive extragenic palindromic-PCR (rep-PCR) typing was carried out using primer combinations rep-1 and rep-2 (Table 1) as previously described. 16 The DNA banding patterns were analyzed by GelCompar II software version 4.0 (Applied Maths, Sint-Martens-Latem, Belgium). Degrees of homology were determined by Dice comparisons, and clustering correlation coefficients were calculated by the UPGMA (unweighted pair group method with arithmetic averages). Clustering was performed at 80% similarity cutoff.
Results
Clinical data and antimicrobial susceptibility
The samples were isolated from catheter 43.05% (31/72), wound 15.27% (11/72), sputum 15.27% (11/72), blood 11.11% (8/72), urine 8.33% (6/72), abscess 2.77% (2/72), tracheal aspirates 1.38% (1/72), and synovial fluid 1.38% (1/72). The susceptibility testing by disk diffusion method showing resistance to CIP, CAZ, GEN, IMP, MIN, and SAM were 100% (72/72), 95.83% (69/72), 84.72% (61/72), 79.16% (57/72), 33.33% (24/72), and 15.27% (11/72) among isolates, respectively (Table 2). MIC assays showed 76.38% (55/72) of isolates were resistant to IMP, whereas all isolates susceptible to colistin (Table 2).
Based on CLSI guidelines, the isolates were considered resistant to IMP and colistin if the MICs were ≥8 and ≥4 μg/ml, respectively.
Represents linkage of ISAba1 and blaOXA-69.
Represents linkage of ISAba1 and blaOXA-80.
A, abscess; B, blood; C, catheter; SF, synovial fluid; SP, sputum; TA, tracheal aspirates; U, urine; W, wound.
CAZ, ceftazidime; CIP, ciprofloxacin; GEN, gentamicin; IMP, imipenem; IVR, internal variable region of class 1 integrons; MIC, minimum inhibitory concentration; MIN, minocycline; SAM, ampicillin/sulbactam.
PCR amplification and mapping of class 1 integrons
The presence of integrase gene (intI1) was detected by PCR in 58.28% (42/72) of A. baumannii isolates, indicating that class 1 integron is widespread among clinical isolates. The cassette arrangement of class 1 integron was characterized by amplification and sequencing of gene cassettes in the internal variable region of class 1 integrons. Seven gene cassette arrays, including aadA4-catB8-aadA1 (2.77%, 2/72) with band size 2,378 bp, aadA1-aadA4 (1.38%, 1/72) with band size 1,650 bp, aacC4-aadA1 (2.77%, 2/72) with band size 1,680 bp, aacC4 (22.22%, 16/72) with band size 517 bp, aadA1 (13.88%, 10/72) with band size 578 bp, aadA4 (5.55%, 4/72) with band size 665 bp, and catB8 (5.55%, 4/72) with band size 650 bp, were obtained among isolates. In the three remaining strains (4.16%, 3/72), intI1 was detected without cassette gene.
Detection of blaOXA-51-like, blaOXA-23-like, blaOXA-24/40-like, blaOXA-58-like, and blaNDM genes
PCR screening revealed that blaOXA-51-like, blaOXA-23-like, and blaOXA-24/40-like genes were present in 86.11% (62/72), 84.72% (61/72), and 30.55% (22/72) of the strains, whereas blaOXA-58-like and blaNDM were not detected in any of the strains.
PCR-based analysis of ISAba1 and ISAba125 carriage in A. baumannii
The presence of ISAba1 and ISAba125 among isolates was 97.22% (70/72) and 65.27% (47/72), respectively. PCR assays using primers targeting the linkages between ISs and beta-lactamase genes, showing the presence of ISAba1 in the upstream of blaOXA-23-like, blaOXA-51-like, and blaADC, was 54.16% (39/72), 9.72% (7/72), and 56.94% (41/72) among isolates, respectively. Sequencing of ISAba1-blaOXA-51-like gene revealed two alleles, blaOXA-69 (5/7) and blaOXA-80 (2/7). The presence of ISAba125 in the upstream of blaADC was not detected in any of the isolates. Also, PCR amplification showing the rates of disruptions in carO, dacD, and adeS genes were 5.55% (4/72), 4.16% (3/72), and 0% (0/72), respectively. Moreover, sequence analysis revealed that carO and dacD genes were disrupted by ISAba1 and ISAba125, respectively.
Clonal relatedness among A. baumannii strains
Molecular typing of 72 isolates of A. baumannii by rep-PCR has been illustrated in Fig. 1. The profiles generated with rep-PCR primers showed 4–11 bands, ranging in size from 0.1 to 2.5 kb. We identified 15 rep-PCR clusters with an 80% similarity cutoff (Table 2). Cl-12 and Cl-15 were the largest clones, and both of them included five strains of A. baumannii. Each of Cl-5, Cl-9, Cl-10, and Cl-14 contained three strains of A. baumannii. Analysis of isolates by rep-PCR technique further demonstrated 44.44% (32/72) of isolates considered either singleton (not related to another rep-PCR type) or small clusters containing two identical clones.
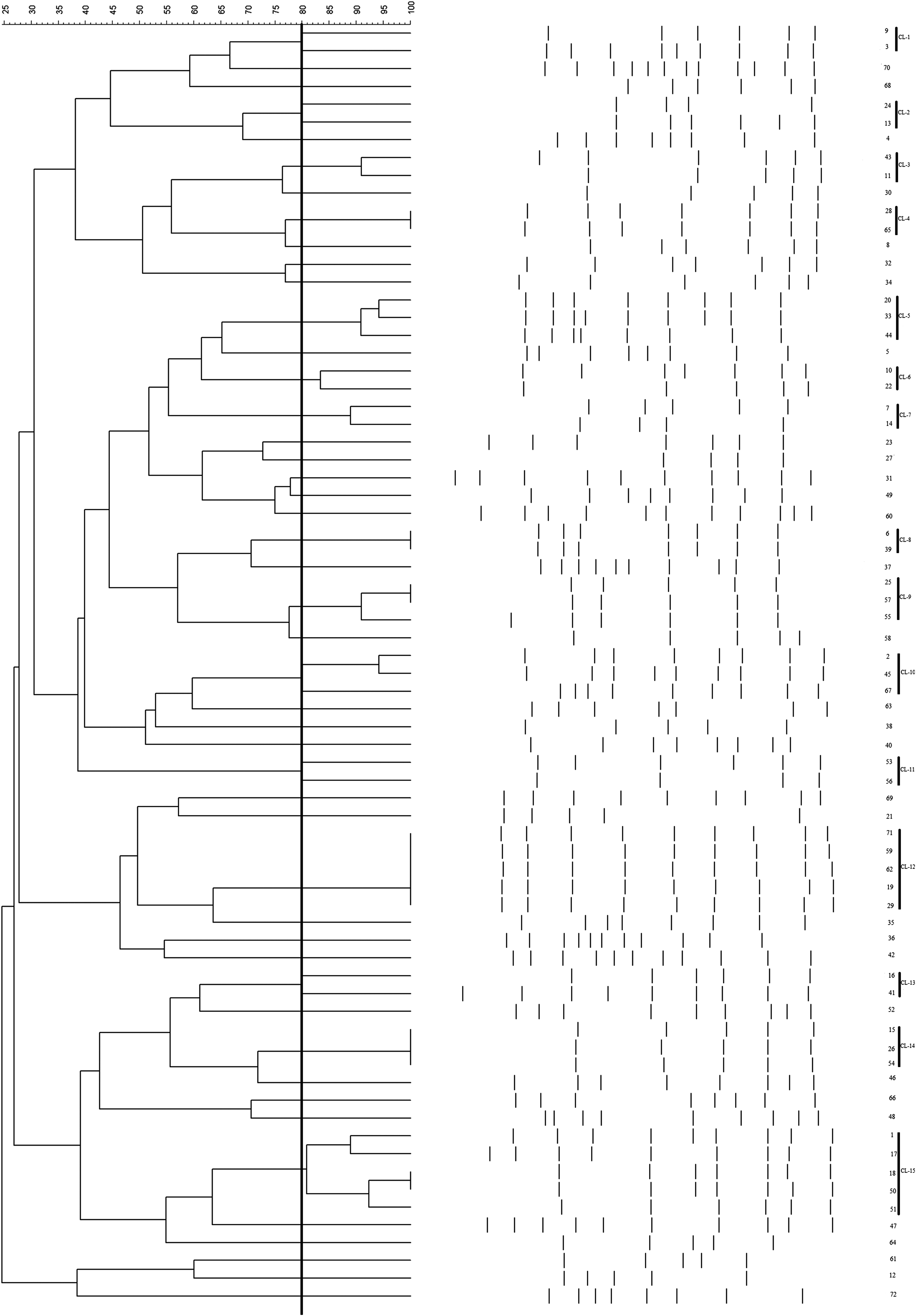
Typing based on repetitive sequence-based PCR patterns in 72 strains of Acinetobacter baumannii. Clustering was performed based on unweighted pair group method with arithmetic mean. The vertical line shows 80% similarity cutoff.
Accession numbers
The sequenced DNAs of the ISAba1-blaADC, carO-ISAba1, dacD-ISAba125, ISAba1- blaOXA-23-like, aadA4-catB8-aadA1, and ISAba1-blaOXA-51-like have been deposited in GenBank database under accession numbers KY039476, KY039477, KY039478, KY039479, KY228988, and KY228989, respectively.
Discussion
A. baumannii is a significant causative agent of nosocomial infections with a particular ability to develop antimicrobial resistance and cause nosocomial outbreaks of infection around the world. 1 Resistance to many antibiotics gives rise to treatment failure of infections associated with this bacterium and results in high morbidity and mortality. 17 Antimicrobial resistance is a global health crisis, because of bacterial capability for accumulating resistance genes through horizontal gene transfer, genomic rearrangement, and genetic mutation. 18 Mobile genetic elements (e.g., integrons and ISs) play a fundamental role in increasing of antimicrobial resistance in A. baumannii isolates. 19
Integrons are remarkable genetic platforms with the ability to acquire, rearrange, and express diverse genes sampled from the microbial pan-genome. 20 On the other hand, they are useful markers for epidemic strains of A. baumannii and epidemiological studies can use valuable information that is suggested by their typing. 21 In this study, 58.28% (42/72) of strains containing class 1 integrons, carry seven gene cassette arrays, including aadA4-catB8-aadA1 (2.77%, 2/72), aadA1-aadA4 (1.38%, 1/72), aacC4-aadA1 (2.77%, 2/72), aacC4 (22.22%, 16/72), aadA1 (13.88%, 10/72), aadA4 (5.55%, 4/72), and catB8 (5.55%, 4/72), and three remaining strains (4.16%, 3/72) showed lack of any cassette gene. Our results revealed that the class 1 integron in A. baumannii isolates is a repertoire of aminoglycoside-modifying enzymes; as previous studies from Iran have shown, there are different cassettes of aminoglycoside-modifying enzymes in A. baumannii strains.22,23
The carbapenem-hydrolyzing class D beta-lactamases are most widespread beta-lactamases with carbapenemase activity in A. baumannii, which have been represented as blaOXA-23-like, blaOXA-24/40-like, and blaOXA-58-like groups. 8 Moreover, blaOXA-51 and blaOXA-69, as two variants of blaOXA-51-like group, have low carbapenemase activities. 24 In this study, we detected the high prevalence (54.16%) of ISAba1 in the upstream of blaOXA-23-like gene. Moreover, sequencing of ISAba1-blaOXA-51-like revealed the presence of two alleles, blaOXA-69 (5/7) and blaOXA-80 (2/7). The oxacillinases have a weak carbapenem-hydrolyzing activity, but when an insertion element is located in the upstream of blaOXA genes, it can rise the high resistance to carbapenems.8,25 Therefore, in the present study, these linkages can be one of the reasons for high resistance to IMP. Conversely, disruption of the chromosomal-encoded carO and dacD genes in isolates of A. baumannii results in permeability defect of IMP influx, thus demonstrating the significant role of carO and dacD in IMP resistance. 8 In this study, the disruption of carO as well as dacD genes due to insertion of ISAba1 and ISAba125 were observed in a few of isolates as shown in Table 2. ISs contributed significantly to the development of antibiotic resistance, as its importance is demonstrated by data from this study and previous studies.6,9,26 However, there are various ranges of mechanisms involved in antimicrobial resistance in A. baumannii, including beta-lactamases, aminoglycoside-modifying enzymes, multidrug efflux pumps, the alteration of target sites, permeability defects, and mobile genetic elements such as class 1 integrons and ISs. 18 The contribution of each resistance mechanism to induction of antimicrobial resistance among bacteria is different and needs a multilayer study, such as gene detection, gene expression, and determination of the efficacy of each resistance apparatus.
Analysis of rep-PCR indicated a high diversity among isolates. There are both local and widespread clones in clinical settings. The widespread clones can exist in different hospitals. For example, in clustering of isolates, we detected Cl-12 and Cl-15 as largest clusters that were distributed in various hospitals.
In conclusion, we applied newly designed primers for simultaneous detection of most prevalent ISs associated with antibiotics resistance in A. baumannii. Our analysis showed the high rates of class 1 integrons as a repertoire of aminoglycoside-modifying enzymes. It seems that linkages of ISAba1-blaOXA-23-like, ISAba1-blaOXA-51-like, and disruptions in carO or dacD can develop resistance to carbapenems among clinical isolates of A. baumannii. Overall, A. baumannii containing ISAba may have a selective advantage when their survival is dependent on the efficient expression of a variety of antibiotic resistance genes or lack of expression of some adhesions or enzymes.27,28 More studies are required to clarify the roles of the ISs in antibiotic resistance, biofilm formation, and bacterial pathogenesis.
Footnotes
Acknowledgment
The authors would like to thank the personnel in the Bacteriology Department of the Pasteur Institute of Iran for their help. This research was supported by the Pasteur Institute of Iran.
Disclosure Statement
No competing financial interests exist.