
Abstract
Objectives:
To describe clinical and molecular characteristics of an outbreak due to metallo-β-lactamases (MBLs) producing Klebsiella pneumoniae collected at Charles Nicolle Hospital of Tunis and to analyze the impact of outer membrane porin (OMP) loss on carbapenem resistance levels.
Methods:
Between 2010 and 2015, 178 carbapenem-resistant Enterobacteriaceae were isolated. Screening for MBL production was performed using combined disk diffusion method, with imipenem and ethylene diamine tetraacetic acid (EDTA) as inhibitors. Resistance genes and virulence factors were identified by polymerase chain reaction (PCR) and sequencing. Genotyping was performed by pulsed-field gel electrophoresis and multilocus sequence typing. Genetic environment of carbapenemase genes was determined by PCR mapping. Conjugation assays were performed, and plasmids were assigned to incompatibility groups by PCR-based replicon typing. OMPs were profiled by sodium dodecyl sulfate-polyacrilamide gel electrophoresis, and porin genes were sequenced.
Results:
Nineteen K. pneumoniae (10.6%) showing MBL activity were isolated from patients hospitalized on four different wards. NDM-1 was the only MBL identified, in association with blaOXA-48. All strains lacked at least one OMP, and carbapenem resistance levels were remarkably elevated in strains lacking OmpK35 and OmpK36. blaNDM-1 was located in IncFIA-type conjugative plasmid, with the same genetic context in all strains. The epidemiological diffusion of blaNDM-1 was due to two clones, one major clone belonging to sequence type (ST) 147 (n = 16) and the other clone belonging to ST307 (n = 3).
Conclusions:
This study describes an outbreak of NDM-1-producing K. pneumoniae strains, isolated from a Tunisian hospital, caused by two clones belonging to ST147 and ST307; and highlights the role of OMPs loss, in combination with β-lactamase expression, in conferring high carbapenem resistance.
Introduction
C
Carbapenemases diffusion among Enterobacteriaceae is of particular importance, since these β-lactamases have the broadest spectrum of activity, usually hydrolyzing most β-lactams. In fact, species producing these enzymes are generally multidrug resistant, resulting in true therapeutic impasses. Furthermore, genes encoding for carbapenemases are most often located on plasmids and are very easily transferable. The mostly reported carbapenemases are KPC, VIM, imipenemase, NDM, and OXA-48 types.
MBL enzymes are reported in nonfermentative bacteria as well as members of Enterobacteriaceae family. The most frequently described MBLs are New Delhi MBL-variants, first identified from Klebsiella pneumoniae in a Swedish patient who was previously hospitalized in India. 5 Since then, most of the outbreaks indicated a link with the Indian subcontinent and in some cases with the Balkan countries. 6 Similar to all other MBLs, NDM efficiently hydrolyses a broad range of β-lactams, including penicillins, cephalosporins, and carbapenems, just sparing monobactams. The emergence and dissemination of NDM-producing isolates have been reported in several countries, including North Africa 7 and in Tunisia.8–10 Furthermore, Tunisia is considered as reservoir of OXA-48-producers and an increasing number of outbreaks due to OXA-48 (and its variants)-producing Enterobacteriaceae have been described.7,11
Within Enterobacteriaceae species, high rates of carbapenem resistance have been recorded among K. pneumoniae clinical isolates in our hospital. This species is a highly important pathogen that causes a wide range of hospital- and community-acquired infections, such as urinary tract infections, pneumoniae, and respiratory tract infections, 12 associated with high morbidity and mortality, mainly in immunocompromised individuals. 13
K. pneumoniae outer membrane usually contains two major nonspecific porins namely OmpK35 and OmpK36, other specific porins (i.e., LamB and PhoE), and a minor quiescent porin (OmpK37). Both OmpK35 and OmpK36 play an important role in antibiotics penetration into the cell, and their loss can contribute to decrease susceptibility or even resistance to cephalosporins and carbapenems.14,15
Since their first description at Charles Nicolle hospital in 2010, 16 regular monitoring of CRE strains has been achieved. This study aimed to characterize K. pneumoniae isolates from our hospital showing positive phenotypic testing for MBL carbapenemases, considering the following objectives: (1) to detect and identify genes coding for MBL enzymes, (2) to assess the role of OMPs in carbapenem resistance, (3) to conduct an epidemiological investigation using chromosome and plasmid fingerprints, and (4) to determine virulence factors of the selected strains.
Materials and Methods
Clinical isolates
Between 2010 and 2015, 12,702 unduplicated enterobacteria strains were isolated. Among them, 178 were ertapenem resistant. Only 19 K. pneumoniae (1 per patient) showing positive phenotypic testing for MBL carbapenemases were analyzed in this study.
Antimicrobial susceptibility testing
Species identification was performed with the API 20E system (BioMérieux, Marcy-l'Etoile, France) and confirmed by matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF; Bruker Daltonics GmbH, Bremen, Germany). Initially, susceptibility to amoxicillin, amoxicillin–clavulanic acid, cefoxitin, ceftazidime, cefotaxime, cefalotin, cefepime, ertapenem, imipenem, aztreonam, amikacin, tobramycin, netilmicin, nalidixic acid, ofloxacin, ciprofloxacin, fosfomycin, tetracycline, minocycline, and tigecycline was determined by disk diffusion using Mueller-Hinton agar (Bio-Rad), according to the European Committee on Antimicrobial Susceptibility Testing (EUCAST) guidelines. 17 Subsequently, minimal inhibitory concentrations (MICs) of amoxicillin–clavulanic acid, cefepime, cefotaxime, ceftazidime, ertapenem, imipenem, and tigecycline were determined with the Vitek 2 system (BioMérieux). In addition, MICs of ertapenem, imipenem, meropenem, and colistin were determined using E-test strips (BioMérieux) according to the manufacturer's instructions. MICs were interpreted according to EUCAST breakpoints. 17
Phenotypic assays
Screening for the production of class B carbapenemases was evaluated by comparing the inhibition zones obtained with imipenem disks, with or without ethylene diamine tetraacetic acid (EDTA) (10 μl of 0.5 M solution). 18 After overnight incubation, the presence of an enlarged zone of inhibition (>7 mm) was interpreted as positive. Each isolate was tested with a disk containing 30 μg of temocillin (Becton, Dickinson & Co.) to facilitate the detection of class D carbapenemases. 11
The production of ESBLs was evidenced by a double-disk synergy test using a combination of amoxicillin/clavulanic acid and aztreonam disks. 19
Molecular analysis of carbapenem resistance genes
Multiplex polymerase chain reactions (PCRs) were performed to detect the most frequently widespread carbapenemases genes of class A (blaKPC and blaGES), B (blaIMP and blaVIM), and D (blaOXA-48-like), as previously described. 20 The detection of blaNDM gene was performed as described elsewhere. 21 PCR products were sequenced, and the nucleotide and deduced protein sequences were analyzed with the software available from the National Center of Biotechnology Information (NCBI) website (www.ncbi.nlm.nih.gov).
Detection of associated resistance genes
ESBL genes (blaCTX-M, blaTEM, blaSHV, blaPER, blaVEB), plasmid mediated AmpC β-lactamases (blaACC, blaMOX, blaFOX, blaDHA, blaCIT, blaEBC), and broad-spectrum β-lactamase genes (blaOXA-1-like) were searched, 20 together with aminoglycoside resistance genes [acc(6′)-Ib, aac(3′)-Ia, aac(3′)-IIa, aac(3′)-IVa, aph(3′)-Ia, aph(3′)-IIa, aph(3′)-VIa, ant(2″)-Ia], 22 and plasmid-mediated quinolone resistance (PMQR) genes (qnrA, qnrB, and qnrS). 23 Subsequent sequencing of PCR products was performed. For the PMQR genes, the identification of qnr alleles was performed by sequencing, otherwise, the aac(6′)-Ib-cr variant was detected by restriction fragment length polymorphism method using FokI (New England Biolabs) restriction enzyme, as previously described. 24
Genetic environment of carbapenemase genes
The genetic environment of blaNDM-1 gene was determined by PCR mapping, as described previously. 3 For blaOXA-48 gene, primers were designed to assess the genetic environment, targeting its right as well as its left ends 25 (Table 1).
MBL, metallo-β-lactamase; Tm, melting temperature.
Isolation and analysis of outer membrane components
Outer membrane protein preparations were obtained after sonication of bacterial cells grown in Mueller-Hinton broth, followed by selective solubilization of the cytoplasmic material with sodium lauroyl-sarcosynate (2%) and ultracentrifugation. 26 Samples were boiled and run on sodium dodecyl sulfate-polyacrylamide gels (11.0%) and stained with Coomassie blue. The encoding sequences of the ompK36 and ompK35 genes of three representative isolates based on pulsed-field gel electrophoresis (PFGE) profiles (one representative strain belonging to clones A and B and to subclone A1) were amplified, sequenced, and aligned with those of ompK gene sequences of K. pneumoniae KCTC2242 producing OmpK35 and lacking OmpK36 (NCBI accession No. CP002910) and K. pneumoniae NTUH-K2044 producing OmpK36 and lacking OmpK35 (NCBI accession No. AP006725).
Conjugation experiments
Conjugation was attempted with three donor isolates selected according to the PFGE results, using rifampin-resistant Escherichia coli J53 as the recipient strain. Transconjugants were selected on Mueller-Hinton agar plates containing 500 mg/L of rifampin with either ceftazidime (8 mg/L) or temocillin (250 mg/L). Resistance profiles of the obtained transconjugants were tested by disk diffusion method and PCR, as previously mentioned.
Plasmid analysis
Plasmid DNA of three isolates and their transconjugants was isolated using Plasmid Isolation Kit (Eurobio) following the manufacturer's guidelines. Plasmids were classified according to their incompatibility group using the PCR-replicon-typing scheme. 27 PCR assays for the repA, traU, and parA genes were performed to determine the genetic backbone of the blaOXA-48 carrying plasmids. 25
Analysis of clonal relatedness
To assess clonality, all isolates were subjected to PFGE, as described by Arlet et al. 28 The obtained fingerprints were interpreted by using the criteria described by Tenover et al. 29
Multilocus sequence typing (MLST) was performed using a previously standardized MLST protocol. 30 The scheme used the following seven housekeeping genes: gapA, infB, mdh, pgi, phoE, rpoB, and tonB. The allelic profile was summarized by assigning a sequence type (ST) via a web database (www.pasteur.fr/recherche/genopole/PF8/mlstKpneumoniae.html).
Virulence factors in K. pneumoniae
The virulence profile was analyzed by PCR to check for the presence of nine genes linked to virulence in K. pneumoniae,31,32 namely capsular serotype K1 and hypermucoviscosity phenotype (magA), allantoin metabolism (allS), regulator of mucoid phenotype A (rmpA), iron system capture (iroN), capsular serotype K2 and hypermucoviscosity phenotype (cps), adhesion type 3 fimbriae (mrkD), iron transport and phosphotransferase function (kfu), siderophore (entB), and siderophore yersiniabactin (ybtS).
Results
Clinical isolates and antibiotic susceptibility
The 19 K. pneumoniae strains (10.6%) showing MBL activity were isolated from patients hospitalized in 4 different wards. Indeed, the first MBL-positive strain was collected from a patient hospitalized in the intensive care unit (ICU) and, 2 months later, the second strain was isolated from a patient hospitalized in the medical ward (Table 2). Specimen sources were urine (n = 8), blood (n = 6), wound (n = 3), and catheter (n = 2).
CIP, ciprofloxacin; CPE, carbapenemase-producing Enterobacteriaceae; F, female; FOS, fosfomycin; GEN, gentamicin; ICU, intensive care unit; IMP, imipenem; M, male; NA, not applicable; ND, not determined; TGC, tigecycline.
The median age of the 19 patients was 47.2 years (range: 6–84 years) and the male to female ratio was 1.11. Thirteen patients were treated, during hospitalization, with imipenem as first-line antibiotic (for eight patients), or as second-line antibiotic instead of amoxicillin/clavulanic acid (for five patients) (Table 2). Besides, combinations of different antibiotic families were used, namely polymyxins (mainly colistin), aminoglycosides (such as amikacin and/or gentamicin), and fluoroquinolones (mainly ciprofloxacin); in association with imipenem. Despite antibiotic treatment, 10 patients died (52.6%) during hospitalization. Death was a result of infection caused by carbapenemase-producing Enterobacteriaceae in seven patients. However, causal relationship between infection and death has not been clearly established in three patients (Table 2).
E-test results showed that the 19 isolates were resistant to ertapenem (MICs range 1.5 to >32 mg/L), while different results were obtained with the Vitek2 system, given that 16 strains were resistant and 3 were susceptible (MICs range <0.5 to >8 mg/L).
For imipenem, 12 strains were resistant and 7 were susceptible (MICs range 0.75 to >32 mg/L) according to E-test results. Furthermore, Vitek2 system revealed that 10 strains were resistant, 6 were intermediate, and 3 were susceptible to the same antibiotic (MICs range <0.25 to >16 mg/L).
Resistance to meropenem was tested only with E-test, and results showed that 15 isolates were resistant and 4 were susceptible (MICs range 1.5 to >32 mg/L).
Only three isolates displayed temocillin resistance, with total absence of inhibition zone.
Results obtained with disk diffusion method indicate that the 19 strains were resistant to tobramycin, netilmicin, chloramphenicol, tetracycline, nalidixic acid, ofloxacin, ciprofloxacin, trimethoprim-sulfamethoxazole, minocycline, and fosfomycin. Furthermore, they were resistant to amoxicillin/clavulanic acid (MICs >32 mg/L), to cefepime (MICs range 16 to >64 mg/L), to cefotaxime (MICs >64 mg/L), to ceftazidime (MICs >64 mg/L), and to tigecycline (MICs range 2 to >8 mg/L). Otherwise, all 19 isolates remained susceptible to colistin (MICs range 0.016–1 mg/L).
Molecular epidemiology
Molecular fingerprints clustered the 19 K. pneumoniae strains into 2 PFGE clonal types (A and B) and one subclone (A1). The largest cluster A belonged to ST147 (15 isolates; 78.9%), while cluster B belonged to ST307 (3 isolates; 15.8%). The subclone A1 belonged to ST147 (Table 3).
+, positive result; —, negative result; COL, colistin; EDTA, ethylene diamine tetraacetic acid; ETP, ertapenem; EUCAST, European Committee on Antimicrobial Susceptibility Testing; MER, meropenem; MIC, minimal inhibitory concentration; MLST, multilocus sequence typing; OMP, outer membrane porin; PFGE, pulsed-field gel electrophoresis; SDS-PAGE, sodium dodecyl sulfate-polyacrilamide gel electrophoresis; ST, sequence type.
Characterization of antimicrobial resistance genes
Among the MBL-type enzymes sought, only NDM-1 was detected in all strains. Moreover, three isolates coproduced blaOXA-48 gene. The blaCTX-M-15, blaOXA-1, and blaSHV-1 genes were detected in all isolates. The distribution of PMQR genes was as follows: aac(6′)-Ib-cr (n = 19), qnrB1 (n = 14), and qnrS1 (n = 4). None of the isolates produced a plasmid mediated AmpC β-lactamase.
Genetic environment of the carbapenemase genes
The PCR mapping showed that the blaNDM-1 gene was preceded by a complete copy of insertion sequence ISAba125 and followed by a bleomycin-resistance gene bleMBL, conferring resistance to bleomycin and bleomycin-like molecules.
Analysis of the genetic environment of the blaOXA-48 gene revealed two copies of the same insertion sequence IS1999, flanking a DNA fragment that includes the blaOXA-48 gene. This genetic structure forms the transposon Tn1999, which is inserted within tir gene. Downstream of blaOXA-48, a lysR gene was identified, followed by a truncated fragment of a gene encoding an acetyl coenzyme A carboxylase.
Outer membrane profiles
Analysis of OMPs showed the presence, for all isolates, of a band of ca. 32 kDa, corresponding to the OmpA-like protein 33 (Fig. 1). Furthermore, they lacked at least one porin (OmpK35, OmpK36, or both).
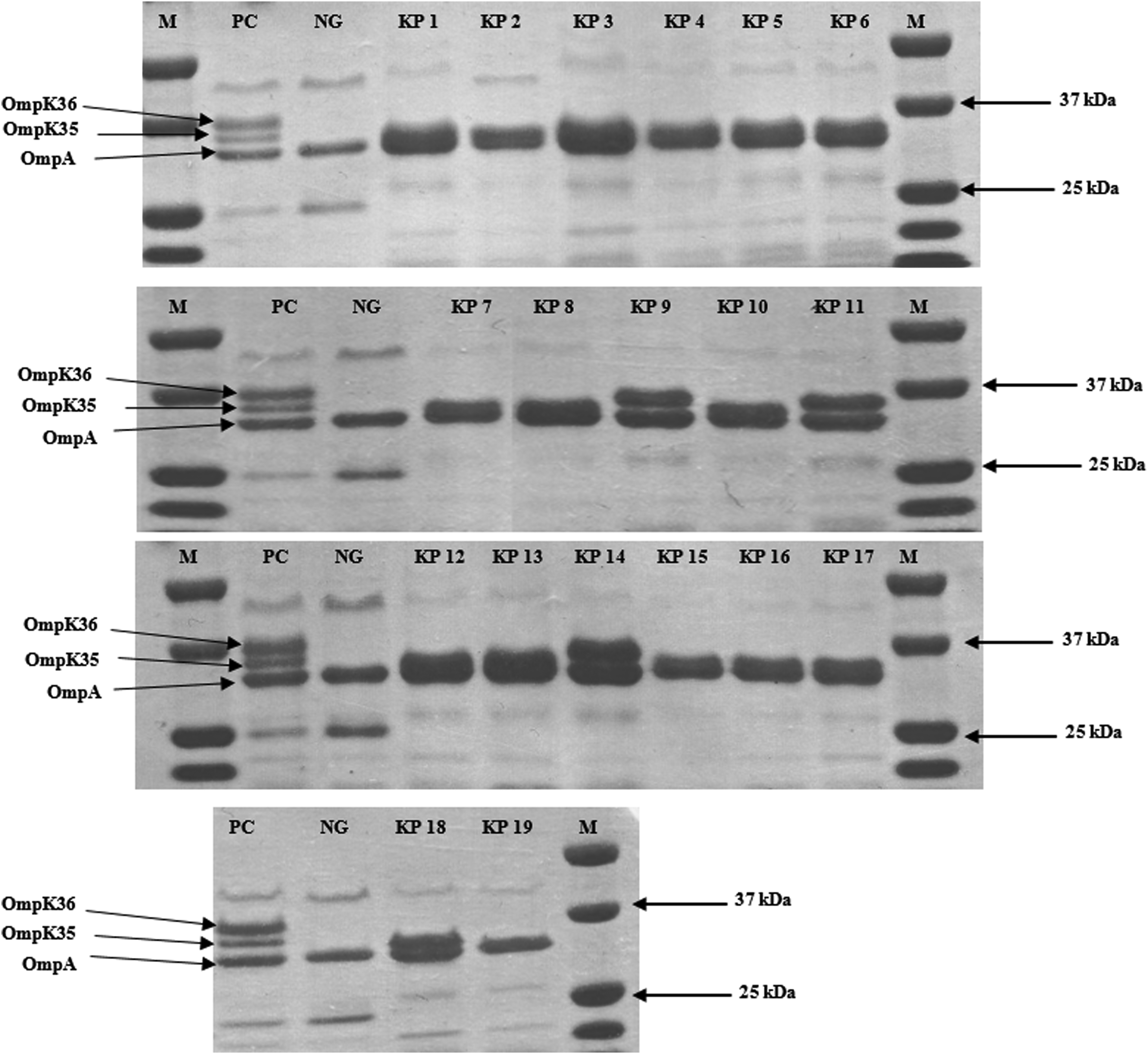
Sodium dodecyl sulfate-polyacrylamide gel electrophoretic analysis of OMPs from NDM-1-producing Klebsiella pneumoniae strain. M, molecular size markers (in kilodaltons); NG, negative control producing neither Ompk5 nor OmpK36; PC, positive control producing both OmpK35 and OmpK36.
Strain KP3 (belonging to pulsotype A, clone ST147) lacked a band of about 35 kDa corresponding to OmpK36. Sequencing revealed that ompK36 contains 38 nucleotide substitutions (including a frameshift mutation at amino acid position 178), an insertion of a 9 nucleotides fragment at position 1149 and a deletion of 3 nucleotides at position 1523. For this isolate, MICs of ertapenem, imipenemase, and meropenem were 4, 0.75, and 2 mg/L, respectively.
Strain KP14 (belonging to pulsotype B, clone ST307) also lacked OmpK36 porin. ompK36 sequencing revealed several substitutions (55 nucleotide substitutions), as well as insertions of 2, 7, 6, 3, 15, 3, and 3 nucleotides at positions 182, 272, 1174, 1315, 1323, and 1332, respectively (leading to a frameshift mutation); in addition to deletions of 3 nucleotides at position 1315 and 1565. For this isolate, MICs of ertapenem, imipenemase, and meropenem were 1.5, 1, and 1.5 mg/L, respectively.
Strain KP19 (belonging pulsotype A1, clone 147) lacked both OmpK35 and OmpK36. Sequencing analysis of ompK35 identified two point mutations (A to T at nucleotide position 1335 and A to G at nucleotide position 1622) and one nucleotide insertion at position 1337 causing a premature codon stop at amino acid position 26, leading to a truncated protein. The ompK36 gene contains 42 nucleotide substitutions, with a premature stop codon at amino acid position 25, as well as 9 nucleotides insertion at position 1177, and 3 nucleotides deletion at position 1552. This isolate showed high carbapenem resistance level (>32 mg/L).
Conjugation experiments
Four transconjugants were obtained from the three selected donors (KP3, KP14, and KP19), and all of them showed MBL phenotype (Table 4). They all exhibited blaNDM-1 gene, and additional resistance markers were cotransferred, including blaCTX-M-15, blaTEM-1, blaOXA-1, and blaOXA-48. According to E-test results, decreased susceptibility to ertapenem (MICs range 0.5–4 mg/L), to imipenem (MICs range 0.19–1 mg/L), and to meropenem (MICs range 0.125–1.5 mg/L) were reported, comparing with the donors. No other antibiotic resistance markers were cotransferred, with the exception of tobramycin and minocycline in E. coli J53 transconjugant (Tc) KP19-1 and E. coli J53 Tc KP19-2.
CHL, chloramphenicol; MNO, minocycline; NAL, nalidixic acid; NET, netilmicin; OFX, ofloxacin; SXT, trimethoprim-sulfamethoxazole; Tc, transconjugant; TET, tetracycline; TOB, tobramycin.
Plasmid analysis
Plasmid DNA analysis of the three E. coli transconjugants derived from KP3, KP14, and KP19 isolates showed that blaNDM-1 gene was carried on IncFIA-type plasmid (Table 4).
The blaOXA-48 gene was carried on a plasmid that possesses a pOXA-48-like backbone. Results of PCR assays for the repA, traU, and parA genes confirmed that each plasmid was very closely related to pOXA-48.
Virulence factors
Of the nine virulence factors searched, entB and mrkD were detected in all isolates. Moreover, ybtS was detected in 15 isolates (78.94%) (Table 3).
Discussion
In this study based on 6-year survey (2010–2015), we describe an outbreak involving 19 NDM-1-producing K. pneumoniae isolates collected from Tunisian patients in healthcare facilities, having no apparent epidemiological link to an endemic area. Thereby, Tunisia is facing a serious health problem, given that OXA-48-producing Enterobacteriaceae are already endemic in this country.
Reports describing the emergence and spread of strains carrying blaNDM-1 gene have increased around the world over the last few years. In 2013, Ben Nasr et al. 8 described the first K. pneumoniae strain coproducing NDM-1 and OXA-48 enzymes isolated from a Libyan patient hospitalized in the ICU in a Tunisian hospital, highlighting the role of travelers in the dissemination of pathogens. Furthermore, two recent reports have described the diffusion of NDM-1-producing strains in Tunisian healthcare settings.9,10
The introduction of the first NDM-1-producing strain in our hospital has been attributed to index patient 1, a 68-year-old female, transferred from a Tunisian private hospital and admitted to the ICU of our hospital in February 2014. Furthermore, the majority of the specimens were from patients hospitalized in the ICU (n = 10) showing that, in this setting, NDM-1-producing K. pneumoniae isolates are associated with nosocomial infections in seriously ill patients.
During hospitalization, most patients were treated with imipenem in association with other β-lactams, fluoroquinolones, and aminoglycosides. Despite this antibiotic treatment, the mortality rate was high, being 52.6%. Our findings show an urgent need to control the use of antibiotics, especially in the ICU, and the selection of appropriate antibiotics by monitoring the dose, the treatment duration, and the administration route to obtain optimal clinical results and to limit the selection of multidrug-resistant strains.
In all isolates, we observed that there were major discordances between MICs obtained with automated instrument and gradient MICs. In fact, if the Vitek2 system was the only technique used for the detection of carbapenem resistance, some of our strains would be lost. Reports have shown that automated instruments could fail in correctly evaluating the correct MICs of several organisms. 34
Enterobacteriaceae isolates with KPC enzymes were not initially recognized because the carbapenem MICs were in the susceptible range when applying breakpoints to susceptibility tests results. 35 This susceptibility has been observed for producers of all types of carbapenemases, 36 and this is particularly true for OXA-48 producers that do not coproduce an ESBL.37,38 To minimize these false-susceptible errors, both Clinical and Laboratory Standards Institute (CLSI) and EUCAST have added separate cutoff values for detecting putative carbapenemase producers for infection control and public health purposes that are considerably lower than the clinical breakpoints.19,39
Different carbapenem resistance levels were observed, which is probably due to differences in the expression levels of carbapenemase genes, 5 or the loss of either one or both of the two major porins (OmpK35 and OmpK36), especially in K. pneumoniae strains harboring ESBLs, 40 as observed in our strains.
All isolates showed a multidrug-resistant phenotype, but remained susceptible to colistin, according to E-test results. For colistin susceptibility testing, gradient diffusion (E-test) is not a standardized technique, according to the EUCAST guidelines. 17
Genetic environment of blaNDM-1 showed the presence of ISAba125 on the upstream region and bleomycin resistance gene bleMBL on the downstream region, which is a genetic structure similar to those reported elsewhere. 41 Furthermore, diversity of the genetic environment of blaNDM-1 has been reported. 42 Three strains were additionally carrying the blaOXA-48 gene, and PCR mapping showed the previously identified Tn1999 transposon. 25 Moreover, the genetic backbone of blaOXA-48 was linked to the epidemic plasmid pOXA-48, identified from diverse geographical origin.
All isolates also coproduce blaCTX-M-15 gene as well as blaTEM-1 and blaOXA-1 and different combinations of PMQR genes. These multidrug resistance determinants are plasmid mediated and can disseminate easily across several unrelated genera. Besides, the dissemination of blaNDM-1 and blaOXA-48 carbapenemases together with ESBLs genes in K. pneumoniae isolates are a source of concern, since carbapenems are antibiotics of last resort for the treatment of multidrug-resistant strains producing ESBL.
OmpK35 loss may be one of the factors contributing to the antibiotic resistance among ESBL-producing K. pneumoniae, and OmpK36 may play an important role in the resistance or reduced susceptibility to carbapenems in K. pneumoniae that produce ESBL or AmpC-type-β-lactamases. 43 Our strains were lacking either one or both OMP, and we observe that carbapenem MICs were remarkably elevated in strains lacking both OMPs. These results emphasize the role of the two major porins OmpK35 and OmpK36, in association with β-lactamases production, including carbapenemases and ESBL, in increasing carbapenem resistance among K. pneumoniae strains.
Transfer of blaNDM-1 and blaOXA-48 genes to E. coli J53 strain was successful. Furthermore, only blaNDM-1 was transferred from an isolate harboring both blaNDM-1 and blaOXA-48, suggesting that these two genes are not located on the same plasmid.
All K. pneumoniae strains possess several virulence genes namely mrkD that facilitates adhesion to extracellular matrix proteins and mediates binding to cells derived from the urinary tract 44 as well as the iron-scavenging system entB, sufficient to cause robust replication in the lung and sepsis. Besides, 80% of them produce yersiniabactin, a siderophore encoded by the Yersinia high-pathogenicity island that is less frequent in K. pneumoniae. These virulence factors are generally carried by plasmids that probably carry different antibiotic resistance genes, and their interspecies dissemination could increase strains pathogenicity.
The dissemination of blaNDM-1 in this study was caused by two clones, based upon PFGE and MLST results; therefore, several NDM-1-producing clones could be present in Tunisia. Besides, ST147 is a major drug-resistant pathogen among carbapenemase-producing K. pneumoniae, found worldwide and in Tunisia, 10 while ST307 has been reported in NDM-1-producing K. pneumoniae strains. 45
In conclusion, we characterize an outbreak of NDM-1-producing K. pneumoniae isolates from Tunisian patients. The emergence and diffusion of multiresistant clones producing MBL in our hospital require strengthening of infection control and prevention strategies, including appropriate antibiotic therapy and colonized patients' surveillance to minimize the spread of these organisms.
Footnotes
Acknowledgment
This study was financially supported by the Ministry of Scientific Research Technology and Competence Development of Tunisia.
Disclosure Statement
No competing financial interests exist.