
Abstract
Three ColV virulence plasmids carrying antibiotic resistance genes were assembled from draft genome sequences of commensal ST95, ST131, and ST2705 Escherichia coli isolates from healthy Australians. Plasmids pCERC4, pCERC5, and pCERC9 include almost identical backbones containing FII-2 and FIB-1 replicons and the conserved ColV virulence region with an additional ColIa determinant. Only pCERC5 includes a complete, uninterrupted F-like transfer region and was able to conjugate. pCERC5 and pCERC9 contain Tn1721, carrying the tet(A) tetracycline resistance determinant in the same location, with Tn2 (blaTEM; ampicillin resistance) interrupting the Tn1721 in pCERC5. pCERC4 has a Tn1721/Tn21 hybrid transposon carrying dfrA5 (trimethoprim resistance) and sul1 (sulfamethoxazole resistance) in a class 1 integron. Four FII-2:FIB-1 ColV-ColIa plasmids in the GenBank nucleotide database have a related transposon in the same position, but an IS26 has reshaped the resistance gene region, deleting 2,069 bp of the integron 3′-CS, including sul1, and serving as a target for IS26 translocatable units containing blaTEM, sul2 and strAB (streptomycin resistance), or aphA1 (kanamycin/neomycin resistance). Another ColV-ColIa plasmid containing a related resistance gene region has lost the FII replicon and acquired a unique transfer region via recombination within the resistance region and at oriT. Eighteen further complete ColV plasmid sequences in GenBank contained FIB-1, but the FII replicons were of three types, FII-24, FII-18, and a variant of FII-36.
Introduction
P
Colicin V (ColV) plasmids are large (80–180 kb), low copy number and contain replicons of both FII and FIB types.1,2 They have been reported in Escherichia coli from human 3 and avian2,4,5 infections and from poultry-associated Salmonella.6,7 They include a transfer region that is closely related to that of plasmid F,2,5,8 and some have been shown to be conjugative. 1 ColV is a bacteriocin that has been associated with virulent strains of E. coli since the 1920s, 1 but is not itself a virulence determinant. 9 However, plasmids containing the ColV operon also include determinants that are associated with bacterial virulence. 1 Early studies on ColV plasmids revealed that they provide host cells with an increased ability to scavenge iron, as well as to tolerate stresses associated with mammalian hosts, such as phagocytosis and serum complement. 1
A number of the virulence-associated genes and operons found in ColV plasmids have been characterized. They include determinants of three different siderophore systems that enable bacteria to scavenge free iron more effectively, and this is advantageous in conditions of iron limitation, such as in urine or serum. 10 The first ColV plasmid-associated siderophore to be identified was aerobactin, 11 but they also contain genes for salmochelin production and uptake, 12 and the Sit (Salmonella iron transport) system, which transports manganese as well as iron. 13 A further, putative iron transport system called Eit (E. coli iron transport) has also been identified. 14 Other virulence determinants in ColV plasmids include the iss increased serum survival gene that confers resistance to serum complement. 15 The hlyF gene, previously believed to encode a hemolysin, 16 has recently been shown to encode a protein involved in the regulation of outer membrane vesicle production. 17 A putative outer membrane protease, OmpT, 18 and a putative ABC transport system, Ets (E. coli transport system), 5 have also been found, but not studied in detail. A gene for a temperature-sensitive hemagglutinin, tsh, is in some ColV plasmids. 19
The oldest ColV plasmids, including those used for the early studies on the group, have not been completely sequenced. However, a 2006 study identified a conserved virulence region linked to the FIB replicon that contains the ColV, aerobactin, salmochelin, sit, and ets operons and the ompT, hlyF, and iss genes. 5 The tsh gene and eit operon were found less frequently in avian E. coli isolates, and they were considered part of a “variable” virulence region. 5
We recently reported the sequence of pCERC3, a ColV virulence-multiresistance plasmid from 22.1-R1, a commensal ST95 E. coli isolated in 2010 from the feces of a healthy Australian adult. 8 Apart from containing all of the virulence-associated genes of the conserved virulence region, pCERC3 contains a complex region conferring resistance to seven antibiotics. A number of other ColV plasmids for which sequences were available were found to contain antibiotic resistance determinants (see Table S2 in Ref. 8 for detail). This convergence of virulence and antibiotic resistance determinants in conjugative plasmids is of great concern, particularly when these plasmids become associated with pathogenic clones such as E. coli ST95 and ST131.
The Hall Lab commensal E. coli collection contains 51 antibiotic resistant strains, including 22.1-R1, and 60 susceptible strains.20–22 FII and FIB replicons were shown by PCR to be the most common types in the collection and were usually found together. 22 The genomes of the resistant strains have recently been sequenced, 23 and here, we report the complete sequences of three further ColV virulence-resistance plasmids recovered from these commensal E. coli genomes. These plasmid sequences and the most closely related complete ColV plasmids found in the GenBank nucleotide database were used to define a shared plasmid backbone, and to track the acquisition and evolution of regions that include antibiotic resistance genes found in them.
Materials and Methods
Plasmid assembly
The strains used in this study are described elsewhere.20–24 Draft genome sequences of 1.10-R8, 11.3-R3, and 14.3-R4 previously obtained 23 are available from NCBI under BioProject number PRJNA335932 as BioSample accessions SAMN05567340, SAMN05567350, and SAMN05567322, respectively. Contigs containing resistance genes, colicin genes, and ColV plasmid-associated virulence determinants were recovered by using standalone Basic Local Alignment Search Tool (BLAST) (www.ncbi.nlm.nih.gov/books/NBK52640).
Junctions between contigs were predicted by using the sequence of pCERC3 and confirmed by using PCR with specifically designed primers. PCR used 20 ng of genomic DNA as a template and amplification conditions that have been previously described. 20 Amplicons were resolved by electrophoresis and sequenced as previously described. 25 Sequencher 5.2.3 (Gene Codes Corporation, Ann Arbor, MI) was used for the final sequence assembly. Reading frames were predicted by using Gene Construction Kit (Textco BioSoftware, Raleigh, NC) and annotated manually.
Multilocus sequence typing and plasmid replicon sequence typing
Contigs containing multilocus sequence typing (MLST) genes were recovered from draft genome assemblies by using standalone BLAST. Alleles and sequence type (ST) numbers were assigned by using the Warwick E. coli MLST database (http://mlst.warwick.ac.uk/mlst/dbs/Ecoli). 26 14.3-R4 was previously assigned to ST3182 (adk10, fumC11, gyrB41, icd10, mdh7, purA8, recA2) by using Sanger sequencing from PCR templates. 22 However, using sequences from the 14.3-R4 genome, the 460 bp gyrB locus was allele 4, which differs by three single nucleotides from the gyrB sequence previously obtained. Thus, the ST of 14.3-R4 has been revised to ST2705 (adk10, fumC11, gyrB4, icd10, mdh7, purA8, recA2).
Contigs containing FII and FIB plasmid replicons were recovered from draft genome assemblies by using standalone BLAST. These contigs and complete plasmid sequences from GenBank were subjected to replicon sequence typing of the FII and FIB replicons by submission to the plasmid MLST (pMLST) site http://pubmlst.org/plasmid.
Sequence analysis
DNA and amino acid sequences were compared by using nucleotide and protein BLAST searches (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Insertion sequences were identified by using IS Finder (https://www-is.biotoul.fr). Resistance genes in ColV plasmids found in GenBank were identified by using ResFinder (https://cge.cbs.dtu.dk//services/ResFinder).
Plasmid copy number relative to the bacterial chromosome was determined by dividing the average coverage of FII and FIB replicon-containing contigs by the average coverage of the seven chromosomal contigs containing MLST genes.
Conjugation
The ability of plasmids to conjugate from commensal strains was tested by using the nalidixic acid-resistant laboratory E. coli strain UB5201 (pro met recA56 nalA) 27 as a recipient. Surface and liquid matings were performed as previously described. 22 The conjugation frequency of pCERC5 was determined by mating UB5201 containing pCERC5 with streptomycin-resistant E. coli UB1637 (his lys trp recA56 rpsL-SmR). 27
Nucleotide sequence accession numbers
The complete sequences of pCERC4, pCERC5, and pCERC9 have been deposited in GenBank under accession numbers KU578032, KU664810, and KY007017, respectively.
Results
FII-2 and FIB-1 ColV plasmids from commensal E. coli strains
The complete 4,059 bp ColV operon was found in the draft genomes of three E. coli strains, 1.10-R8 (ST95), 11.3-R3 (ST131), and 14.3-R4 (ST2705), in the Hall Laboratory collection. These strains had previously been shown to contain FII and FIB plasmid replicons by using PCR, 22 and here, these replicons were typed as FII-2 and FIB-1 by using the genome data. Elements of the conserved ColV virulence region, including the aerobactin, salmochelin, and sit siderophore synthesis operons, the ets operon, ompT, and iss were present in the draft genomes, but the tsh gene and eit operon were not found.
Complete FII-2:FIB-1 plasmids were assembled from 7 (1.10-R8 and 14.3-R4) or 10 (11.3-R3) contigs found in the draft genomes and named pCERC4, pCERC5, and pCERC9. These plasmids ranged in size from 135,661 to 143,812 bp (Table 1), and they contained most of the antibiotic resistance genes previously identified. 23 Only the tet(B) tetracycline resistance determinant of 1.10-R8 was located in Tn10 elsewhere. The copy number of each plasmid, determined by using coverage relative to the bacterial chromosome, was 1.2, 1.3, and 2.8 for pCERC4, pCERC5, and pCERC9, respectively. In addition to the FII-2 and FIB-1 replicons and the conserved virulence region, each plasmid included genes for production of colicin Ia and genes that were responsible for replication, maintenance, and transfer (Fig. 1). They each contained two copies of IS1, single copies of IS2, IS629, ISEc32, and partial copies of IS200C and IS21. pCERC4 contained an additional IS3 inserted into the traS gene of the transfer region and copies of IS26 and IS4321 in the antibiotic resistance gene region. Each plasmid featured an antibiotic resistance gene region conferring resistance to one or two antibiotics, located at RR1 in pCERC4 and RR2 in pCERC5 and pCERC9.
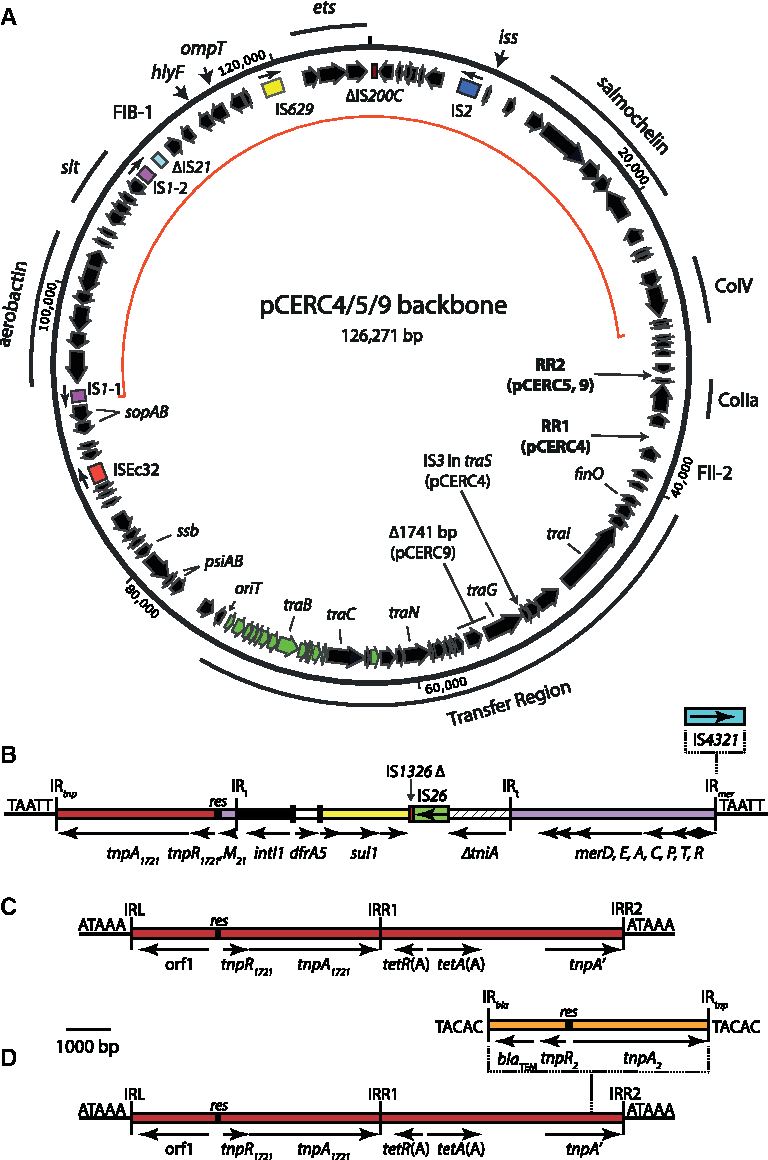
Maps of pCERC4, pCERC5, and pCERC9.
The tet(B) tetracyline determinant is in a complete copy of Tn10 located elsewhere.
Contains IS4321 in IRmer.
Ap, ampicillin; Su, sulfamthoxazole; Tc, tetracycline; Tp, trimethoprim.
pCERC4, pCERC5, and pCERC9 represent a sublineage of ColV plasmids
To generate backbone sequences for comparison, the resistance transposons and the target site duplications (TSD) generated by their insertion were removed from pCERC4, pCERC5, and pCERC9. The IS3 and associated TSD in pCERC4 were also removed. The resulting backbones were 126,270 and 126,271 bp for pCERC4 and pCERC5, respectively (Table 1). Relative to the other plasmids, the pCERC9 backbone (124,528 bp) is missing 1,741 bp from the transfer region. This deletion appears to be the result of recombination between two copies of the 4 bp sequence TTGC. Apart from the deletion in pCERC9, the backbones of all three plasmids are >99.9% identical. Thus, the FII-2:FIB-1 backbone contains both the ColV and ColIa determinants, the conserved virulence region, and a complete 32.7 kb F-like transfer region. Similar to the FII-18:FIB-1 plasmid pCERC3, 8 the transfer region found in the FII-2:FIB-1 backbone is mosaic. Genes at the oriT (origin of transfer) end of the transfer region (green in Fig. 1A) are >99.5% identical to the corresponding genes in plasmid F, whereas the rest of the transfer region is 93.8–99% identical to that of F.
The antibiotic resistance gene in pCERC4
The two antibiotic resistance genes in pCERC4, dfrA5 and sul1, are located within a region, hereafter referred to as RR1 (Fig. 1A) and part of a hybrid Tn1721/Tn21 transposon created via a crossover in the res site (Fig. 1B). The inverted repeats (IR) that bound this transposon are flanked by the TSD TAATT, indicating that this hybrid transposon form moved to this location. The transposition module at the left end of the transposon, from the IR at the tnp end (IR tnp ) to the res resolution site, is derived from Tn1721 or Tn1722. 28 The sequence to the right of res is derived from Tn21, with a class 1 integron in the same position as the In2 integron in Tn21. 29 The class 1 integron contains a dfrA5 gene cassette and features the classical sul1-containing 3′-conserved segment (3′-CS). A copy of IS1326 at the end of the 3′-CS has been interrupted by IS26, which has removed the remainder of IS1326 and part of the tni region. This exact junction between IS26 and the remainder of the tniA gene is present in many GenBank entries, including integrons containing both the sul1-containing 3′-CS (e.g., GenBank accessions KF976462, KR822247, and KP899805) and integrons with the rarer sul3-conserved segment. 8 The complete mer operon of Tn21 is present, and a copy of IS4321 30 is inserted into IR mer .
The RR2 type regions in pCERC5 and pCERC9
In pCERC5 and pCERC9, the antibiotic RRs (Fig. 1C, D) are in the same position in the plasmid backbone, RR2 (Fig. 1A). In both, a copy of Tn1721, containing tetA(A) (tetracycline resistance), is inserted between the ColV and ColIa operons, with the TSD ATAAA (Fig. 1C, D). In pCERC5 a second transposon, Tn2, 24 containing blaTEM (ampicillin resistance), has been inserted into the tnpA′ gene of Tn1721, creating the TSD TACAC (Fig. 1D).
Transfer of the FII-2:FIB-1 plasmids
To test the conjugative ability of pCERC4, pCERC5, and pCERC9, 1.10-R8, 11.3-R3, and 14.3-R4 were mated with the laboratory strain UB5201. As expected, pCERC4 and pCERC9 transconjugants were not recovered under solid surface or liquid mating conditions. For pCERC9, the 1,741 bp deletion in its transfer region, removing traH and 362 bp from the 5′ end of traG, is likely responsible for the inability to transfer. For pCERC4, the inability to transfer is likely due to polar effects of the insertion of IS3 in traS, which is part of a set of transfer genes expressed as an operon from traY to traX.31,32 IS3 may interfere with the expression of genes downstream of traS, including traD and traI, which are required for transfer. 33
pCERC5 was transferred from 11.3-R3 to E. coli UB5201 during a solid surface mating experiment. To assess transfer frequency under standardized conditions, UB5201 containing pCERC5 was mated with E. coli UB1637. pCERC5 was transferred from UB5201 to UB1637 with an average frequency of 3.1 × 10−5 transconjugants/donor (average of three determinations) on an agar surface and about fivefold lower, 5.5 × 10−6 transconjugants/donor (three determinations) in liquid. Surprisingly, the 2,826 bp traG gene in pCERC5 contains a G-to-T substitution at position 422, which introduces a stop codon. This splits the traG gene into two smaller reading frames, traGORF1 and traGORF2, which span positions 1–423 and 496–2,826 of traG, respectively. However, the mutation has not abolished the conjugative ability of pCERC5. These open reading frames (ORFs) are in frame with traG, and their products must retain the essential functions of TraG, which contributes to pilus formation, mating pair stabilization, and entry exclusion. 34
Lineage FII-2 of ColV plasmids
All complete ColV plasmids found in GenBank (last search July 11, 2017) were typed by using the pMLST website and three main groups, distinguished by the FII replicon type, were found (Table 2). Four plasmids contained the FII-2:FIB-1 replicon combination and the same backbone as pCERC4, 5, and 9 (>99.9% identical), and their general structures are shown in Fig. 2A. These plasmids were from E. coli isolates belonging to ST95 (pSF-088-1), 35 ST131 (pG749_1) 36 and untyped E. coli (pDB4277 and pECAZ147_1) (GenBank accessions KP398867 and CP018994, respectively), all derived from human infections in the United States, Europe, or Israel. All three plasmids contain antibiotic resistance genes that are located in variants of the antibiotic RR1 found in pCERC4, which is also from an ST95 E. coli.
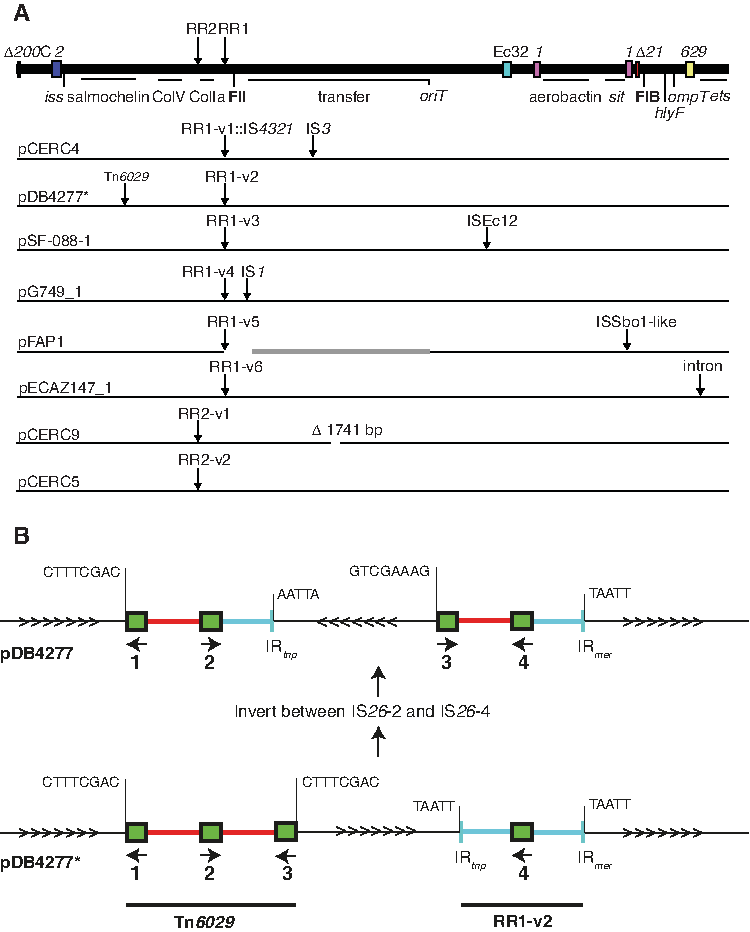
Variation in ColV FII-2:FIB-1 lineage plasmids.
Contains a complete ColV operon and the complete ColV plasmid conserved virulence region; GenBank accession AY545598 contains runs of Ns and was considered incomplete; last search July 11, 2017.
Additional colicins; plasmids were screened for the presence of colicin Ia (cia), B (cba), and M (cma) genes.
Source (community urinary tract infection) and year of isolation were provided by Dr David C. Bean.
A plasmid name is not included in this GenBank accession.
These plasmids are essentially identical.
Two nucleotide differences relative to FIB-1.
One nucleotide difference relative to FIB-40.
The authors of this GenBank entry work in China.
Two nucleotide differences relative to FII-36.
In addition, in pDB4277 a copy of Tn6029 has been inserted into the iroE gene of the salmochelin operon, and a homologous recombination event involving copies of IS26 from RR1 and Tn6029 has resulted in the inversion of 30,222 bp between them (Fig. 2B). This inversion was reversed in silico (Fig. 2B), yielding pDB4277*, to simplify comparisons.
Evolution in situ of RR1
It is likely that RR1 began with the insertion of the Tn1721/Tn21 hybrid transposon adjacent to the ColIa operon, as in pCERC4, before evolving in situ. Variants of this region, none of which contain IS4321 in IR mer , are found in exactly the same position in pDB4277, pSF-088-1, pECAZ147_1, and pG749_1. In those plasmids, the copy of IS26 between the 3′-CS and tni has deleted 2,069 bp to the left, including sul1, leaving 24 bp of 3′-CS (Fig. 3). This copy of IS26 has served as a target for further IS26-mediated insertions in pSF-088-1, pECAZ147_1, and pG749_1 (Fig. 3). In pSF-088-1 and pECAZ147_1, Tn6029, containing the blaTEM (ampicillin resistance), sul2 (sulfamethoxazole), and strAB (streptomycin) genes, has presumably been generated by the insertion of an IS26 translocatable unit (TU),37,38 as the sequence on either side of the original IS26 has been maintained (Fig. 3). Similarly, an additional TU containing aphA1a (kanamycin and neomycin resistance) has been inserted next to IS26 in RR1 of pG749_1 to generate Tn4352 (Fig. 3). In pECAZ147_1, a further segment from Tn1721 carrying the tet(A) tetracycline resistance determinant is also present (Fig. 3). This segment could have been lost or gained via homologous recombination between tnpA and tnpA′, which are identical.
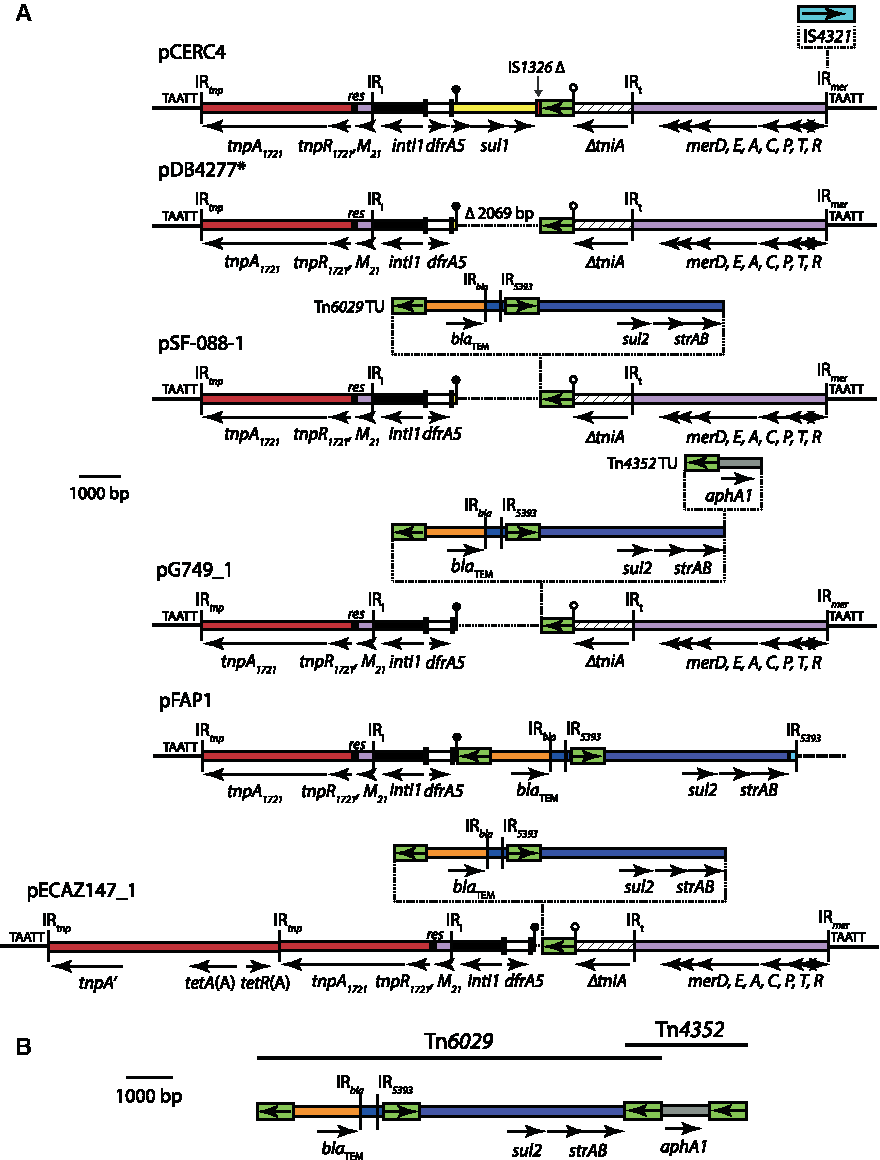
Evolution of RR1 in FII-2:FIB-1 lineage ColV plasmids.
The FIB-1 ColV plasmid from E. coli FAP1 is derived from ColV lineage FII-2
The unnamed plasmid of E. coli strain FAP1 (henceforth referred to as pFAP1 for simplicity), isolated from the feces of a healthy Dutch pig in 2011, 39 contains an FIB-1 replicon, but it does not contain an FII replicon (Table 2). However, it is clearly derived from the FII-2 ColV lineage. It contains a variant of RR1 (Fig. 3), which is adjacent to a unique transfer region (Fig. 3). This RR1 variant is most closely related to that found in pSF-088-1 (Fig. 2). In pFAP1, recombination between Tn5393-derived sequence in Tn6029 and a complete copy of Tn5393 has introduced the last 98 bp of Tn5393 (light blue in Fig. 3). Now, the Tn5393-derived sequence is bounded by IR 5393 instead of IS26. Adjacent to IR 5393 is a plasmid F-like transfer region that is different to the one usually found in ColV FII-2 plasmids (Fig. 2). Beyond oriT at the other end of this transfer region, pFAP1 is >99.9% identical to the ColV FII-2 backbone (Fig. 2). Thus, it appears that recombination at one end between RR1 and Tn5393 in a second plasmid, and between copies of oriT at the other, is responsible for bringing the novel segment into pFAP1.
Discussion
Diversity in ColV virulence plasmids
It is clear that there are a number of variations in the backbone of ColV plasmids. Consequently, to fully understand the role of ColV plasmids, which carry multiple virulence determinants, in the spread of antibiotic resistance determinants, it will be important to define each plasmid backbone and then identify sublineages with regions that have arisen via evolution in situ from a single original insertion of a resistance transposon. This will facilitate tracking plasmids, by removing variability due to addition or loss of mobile elements. Here, we have described three new ColV virulence plasmids that were found in commensal strains of E. coli from the feces of healthy individuals who had not taken antibiotics for at least 6 months before sampling. 20 In two cases, the commensal E. coli we studied were from STs that were known to be associated with pathogenesis, namely ST95 (pCERC4) and ST131 (pCERC5). All three plasmids have the same FII-2:FIB-1 backbone type, carry a ColIa determinant, and contain antibiotic resistance determinants. However, they belong to two distinct sublineages when the location and structure of the resistance gene regions are considered.
Sequences of other members of the FII-2:FIB-1 lineage found in GenBank all included only one of the regions that we identified, namely RR1. However, this region has evolved to include further resistance genes. An additional Tn6029 was also found in the backbone of pDB4277, a member of the RR1 sublineage, and an inversion has arisen via homologous recombination between two IS26 (Fig. 2B).
The power of using backbone structure combined with insertion locations to type plasmids is emphasized by the fact that pCERC5 from an ST131 human commensal isolate has RR2 and pG749_1 from an ST131 human infection 36 carries RR1. Though both ST131 isolates were found to carry fimH22, making them members of clade B, according to the typing scheme of Petty et al., 40 the two FII-2:FIB-1 plasmids found in ST131 are from different lineages. A recent screen of 16 ST131 clade B isolates from various locations revealed that four contained FII-2:FIB-1 replicon types. 41 However, the countries of isolation of the FII-2:FIB-1-containing isolates and whether they carried a ColV plasmid or resistance genes could not be traced from the published data.
In contrast, plasmid pSF-088-1, from a clinical ST95 E. coli isolate from the USA, and pCERC4, from a commensal ST95 E. coli from Australia, carry FII-2:FIB-1 plasmids from the same sublineage, consistent with the spread of this strain.
Role of colicin production
Colicins are believed to play a key role in shaping E. coli populations. 42 ColV plasmids also encode determinants for the synthesis of additional colicins such as ColIa, as in the FII-2:FIB-1 and FII-24:FIB-1 groups, or ColB and ColM (Table 2). Production of these colicins may confer an advantage on E. coli in the human gastrointestinal tract, and this could contribute to the persistence of strains carrying these plasmids and their associated antibiotic resistance and virulence determinants. In addition, the siderophore systems encoded by these backbones are likely to give ColV plasmid-containing strains a significant advantage, enabling them to outcompete other commensal strains for free iron in the gut.
FII-2:FIB-1 ColV-ColIa plasmids are widely distributed
Plasmids belonging to the FII-2:FIB-1 lineage examined here were from E. coli isolated in Australia, the United States, Europe, or Israel (Table 2), indicating a global distribution. The E. coli isolates were from a variety of sources, the commensal reservoir, human infections including cases of bacteraemia 35 and urinary tract infection (David C. Bean, pers. comm.), or from the feces of a healthy pig. 39
Other ColV plasmid lineages
A further 20 complete plasmids containing the ColV operon and associated virulence determinants found currently in the GenBank nucleotide database (Table 2) contained FII replicons of three additional types, FII-18 as in pCERC3, FII-24, and a single two-nucleotide variant of FII-36, and most carried FIB-1. Their backbones remain to be defined and compared with those of the FII-2:FIB-1 type described here. Ten of these plasmids, representing the FII-24 and FII-18 lineages, contain one or more antibiotic resistance genes (Table 2) and this convergence of virulence and antibiotic resistance determinants is a matter of concern.
The sources of plasmids from the other ColV lineages (Table 2) were similar to those of the FII-2:FIB-1 group. They were from E. coli or Salmonella enterica isolated in various countries between 1989 and 2017. E. coli isolates were from cases of human urinary tract infection, 43 neonatal meningitis3,44 or from animal disease2,4,45–47 and from healthy humans.8,48 The S. enterica isolates were from healthy chickens or chicken meat. 7 Again, several of these plasmids were recovered from E. coli isolates belonging to ST95 (pCERC3, pECOS88)3,35 or the ST95 clonal complex (p1ColV5155) 4 and ST131 (pJIE186-2). 43
Conclusions
Clearly, ColV plasmids of various lineages are globally distributed and found in important pathogenic clones, and these may contribute to their success. In addition, most sequenced plasmids in all three of the main lineages contain antibiotic resistance genes, enabling transfer of virulence and resistance properties together. It will be interesting to examine ColV plasmids from the remaining lineages, to determine whether, similar to the FII-2:FIB-1 group, transposons carrying resistance genes have been acquired on only a few occasions, followed by evolution in situ.
Finally, the carriage of such virulence-antibiotic resistance plasmids by commensal E. coli underlines the importance of the commensal reservoir. These bacteria can cause extraintestinal infections, or they can transfer the virulence and antibiotic resistance determinants that they harbor to other pathogens. 49
Footnotes
Acknowledgments
The authors thank Dr David C. Bean for supplying information about pDB4277. R.A.M. was supported by an Australian Postgraduate Award, and the study was supported by the School of Molecular Bioscience at The University of Sydney.
Disclosure Statement
No competing financial interests exist.