
Abstract
Food-producing animals represent one of the sources of antibiotic resistant commensal bacteria. There is an increasing awareness that these bacteria might have the potential to transfer their resistance genes to other (pathogenic) bacteria. In this study, 50 commensal Escherichia coli strains originating from food-producing animals and resistant to the “highest priority, critically important antibiotics” cefotaxime and/or ciprofloxacin, were selected for further characterization. For each strain (i) an antibiogram, (ii) the phylogenetic group, (iii) plasmid replicon type, (iv) presence and identification of integrons, and (v) antibiotic resistance transfer ratios were determined. Forty-five of these strains were resistant to 5 or more antibiotics, and 6 strains were resistant to 10 or more antibiotics. Resistance was most common to ampicillin (100%), sulfamethoxazole, ciprofloxacin (82%), trimethoprim, tetracycline (74%), cefotaxime, (70%) and ceftazidime (62%). Phylogenetic groups A (62%) and B1 (26%) were most common, followed by C (8%) and E (4%). In 43 strains, more than 1 replicon type was detected, with FII (88%), FIB (70%), and I1 (48%) being the most encountered types. Forty strains, positive for integrons, all harbored a class I integron and seven of them contained an additional class II integron. No class III integrons were detected. The antibiotic resistance transfer was assessed by liquid mating experiments. The transfer ratio, expressed as the number of transconjugants per recipient, was between 10−5 and 100 for cefotaxime resistance and between 10−7 and 10−1 for ciprofloxacin resistance. The results of the current study prove that commensal E. coli in food-production animals can be a source of multiple resistance genes and that these bacteria can easily spread their ciprofloxacin and cefotaxime resistance.
Introduction
L
Food-producing animals represent one of the sources of antibiotic resistant bacteria, but it remains challenging to determine their risk to human health due to the complexity of transmission routes between farms and consumers and the frequent transfer of resistance genes among host bacteria.3,4
Both pathogenic bacteria and commensal bacteria can harbor and transfer antimicrobial resistance genes. While the ingestion of antibiotic resistant pathogenic bacteria may represent a direct hazard to public health, the transmission of antibiotic resistance genes from commensal bacteria may represent an indirect hazard, increasing the gene pool from which (pathogenic) bacteria can acquire resistance genes. 5
Since the first observation of multiresistant bacteria in the 1950s, 6 it has become clear that antibiotic (multi)resistance is actually an ancient natural phenomenon, exhibited by the presence of (transferable) resistance genes in metagenomes of ancient isolated caves and permafrost sediments.7–10 Emergence of multiresistance and transfer of antimicrobial resistance genes are associated with genetic elements such as plasmids, transposons, and integrons. Plasmids inherently are able to exist extra-chromosomally, replicate autonomously, and have the ability to be horizontally transferred between distinct hosts. 11 Many known antibiotic resistance genes are located on plasmids. 12 Historically, plasmids have been classified in incompatibility (Inc) groups on the basis of mutual incompatibility, that is, two plasmids of the same group, share common replication and partitioning elements, and are unable to stably coexist in the same cell. 13 Plasmids have been shown to be very efficient tools in the acquisition and dissemination of antimicrobial resistance between members of the Enterobacteriaceae family. 14 Many resistance plasmids carry additional genetic elements such as transposable elements and integrons.
In 1989, integrons were identified as key players in the dissemination of resistance genes among Gram-negative bacterial pathogens, mainly by their location on transposable elements and conjugative plasmids. 15 Integrons are immobile elements that can easily capture, integrate, express, and release gene cassettes encoding antibiotic resistance genes. 16 Based on the sequence of the encoded integrases, five different integron classes can be distinguished among the mobile integrons, of which only the first three are historically associated with the dissemination of multiresistance. 16
On January 1, 2014, the harmonized monitoring and reporting of antimicrobial resistance became mandatory in the EU for the pathogens Salmonella spp. and Campylobacter jejuni and was recommended for Campylobacter coli, indicator commensal Escherichia coli, Enterococcus faecalis, and Enterococcus faecium. In 2011, the Federal Agency for the Safety of the Food Chain (FASFC) in Belgium started with the monitoring of antimicrobial resistance in indicator bacteria in fecal samples of food-producing animals. 17 Since commensal E. coli have to survive in the lower intestinal tract, they have to gain resistance by acquisition of resistance genes or the development of mutations when challenged by antimicrobial pressures imposed on their host. Hence, commensal E. coli are generally regarded as good indicators for resistance among Gram-negative bacteria. 14 In addition, monitoring the commensal antibiotic reservoir is important as it has been suggested that commensal antibiotic resistance can be a greater global health threat than the direct selection pressure on the pathogenic bacteria. More specifically because the occasional de novo development of resistance due to mutations in a pathogen may be less frequent and less impactful than the constant gene traffic from the vast commensal reservoir into the relatively small pathogen pool. 18
To manage the use of antibiotics and cope with the worldwide problem of emerging multiresistance, the WHO ranked antibiotics according to their importance in human medicine. 19 Among the highest priority, critically important antibiotics on the list are ciprofloxacin (second generation fluoroquinolone) and cefotaxime (third generation cephalosporins). Several studies indicate that the prevalence of cefotaxime and/or ciprofloxacin resistant bacteria in food-producing animals has reached worryingly high values.20–24
In this study, a collection of commensal E. coli strains originating from food-producing animals was selected based on the presence of resistance to cefotaxime or ciprofloxacin. The goal of this study was to further characterize these selected strains by (i) determining the antibiogram, and assessing the (ii) phylogenetic group, (iii) plasmid replicon type, (iv) presence and identity of integrons, and (v) antibiotic resistance transfer ratio.
Materials and Methods
Bacterial strains
A total of 845 E. coli isolates were analyzed for antibiotic susceptibility (EUMSV2 plate Trek Diagnostics, West Sussex, United Kingdom) by the Belgian national reference laboratory, Veterinary and Agrochemical Research Centre (CODA-CERVA) within the nationwide antimicrobial resistance surveillance program of commensal indicator bacteria of the Belgian Federal Agency for Safety of the Food Chain (AFSCA-FAVV) for 2013. These strains originated from cecal content of broiler chickens (n = 234) and fecal material of pigs (n = 205), cattle (n = 204), and veal (n = 202). Samples were taken under the authority of the Belgian Food Agency at the slaughterhouse or at the farm. Eighty-five strains resistant to cefotaxime and/or to ciprofloxacin were selected. For the ciprofloxacin resistant strains, only the strains which were resistant to ciprofloxacin and not to nalidixic acid were selected, as this reduces the chance that the resistance is caused by a chromosomal mutation. From this selection, 50 strains, showing pure culture on agar plates, mainly originating from broiler chickens, were further analyzed in this study (i.e., 15 ciprofloxacin resistant and 34 cefotaxime resistant strains and 1 strain resistant to both antibiotics).
Antimicrobial susceptibility profiles
In 2014, harmonized monitoring became mandatory for indicator commensal E. coli at EU level and the panel of antimicrobial substances for the antimicrobial susceptibility testing changed. 25 Therefore, the antibiotic susceptibility profiles of the 50 selected strains were determined again with the new panel of 14 antibiotics using the microbroth dilution method EUVSEC Sensititre™ (Thermo Fisher™, West Sussex, United Kingdom). Using an overnight Tryptic Soy Broth culture, a bacterial suspension with an optical density of 0.5 McFarland was prepared by suspending one to three colonies in sterile distilled water. Ten microliters of this suspension were added to 11 ml cation adjusted Mueller Hinton broth with N-tris(hydroxymethyl)methyl-2-aminoethanesulfonic acid (TES) buffer (Thermo Fisher). Fifty microliters of this inoculated broth were transferred to an EUVSEC plate, which contains dilutions of 14 lyophilized antibiotics according to the European directives. 25 The plates were incubated for 24 hr at 35°C before being analyzed with the Sensititre® Vizion® system (Thermo Fisher). The minimum inhibitory concentration (MIC) is defined as the lowest concentration at which no visible growth can be detected. A strain is considered resistant to an antimicrobial substance if the MIC value is higher than the EUCAST epidemiological cutoff values (ECOFF values, https://mic.eucast.org/Eucast2).
Phylogenetic groups
The phylogenetic group of the strains was determined using the protocol of the quadruplex phylogroup assignment method of Clermont et al. with some minor modifications. 26 PCRs were carried out in a total volume of 50 μl containing 1× buffer II (Thermo Fisher) 1.5 mM MgCl2, 40 μM dNTPs, 2 U of AmpliTaq DNA polymerase (Thermo Fisher), 30–80 ng DNA, and the appropriate primers. The amounts of primers used in the quadruplex PCR were 20 pmol, except for AceK.f (100 pmol) and ArpA1.r (100 pmol). In the allele-specific phylogroup E PCR the amounts of primers were 20 pmol for ArpAgpE.f and ArpAgpE.r and 12 pmol for trpBA.f and trpBA.r. In the allele-specific phylogroup C PCR the amounts of the primers trpAgpC.1 and trpAgpC.2 were 20 pmol, while the primers providing the internal control were adjusted: AceK.f (100 pmol) and ArpA1.r (100 pmol). PCRs were performed under the following conditions for the quadruplex PCR: denaturation 5 min at 94°C, 35 cycles of 30 sec at 94°C, 30 sec at 55°C, and 30 sec at 72°C, and a final extension step of 7 min at 72°C. For the allele-specific phylogroup E PCR the annealing temperature was 57°C and for the allele-specific phylogroup C PCR it was 59°C. The PCR amplification products were separated by electrophoresis on a 1% SeaKem LE agarose gel (Cambrex Bio Science, Wiesbaden, Germany) in 0.5 × TAE buffer (Thermo Fisher), visualized by ethidium bromide staining and photographed under ultraviolet (UV) light.
Plasmid replicon typing
To determine which plasmid replicons were present in the strains, five triplex PCRs and five simplex PCRs were optimized based on the PCR-based replicon typing method of Carattoli et al. 27 PCRs were carried out in a total volume of 50 μl containing 1× buffer II, 1.25 mM MgCl2, 50 μM dNTPs, 1.5 U of AmpliTaq DNA polymerase, 30–80 ng DNA. In the triplex PCRs 1 μM of each primer was added, except in the X–L/M–N PCR, in which 2 μM of the X primers were added. For the simplex PCRs, the same composition was used with 1 μM of each primer. The PCR program was as follows: denaturation for 5 min at 94°C, followed by 30 cycles of 1 min at 94°C, 30 sec at the appropriate annealing temperature and 1 min at 72°C, and a final extension step of 5 min at 72°C. The annealing temperature of the 10 PCRs was as follows: 54°C for FII, 60°C for A/C–T–FIIS, FIA–FIB–W, X–L/M–N, B/O, K, R, and U, 64°C for Y–P–FIC, and 65°C for HI1–HI2–I. The PCR amplification products were separated by electrophoresis on a 1% SeaKem LE agarose gel in 0.5 × TAE buffer, visualized by ethidium bromide staining, and photographed under UV light.
Presence of integrons
The presence of integrons was detected with PCR using degenerate primers hep35 and hep36, targeting the conserved regions of integrase genes intI1, intI2, and intI3. 28 The PCR mix contained 1 × buffer II, 0.75 mM MgCl2, 1.5 U AmpliTaq DNA polymerase, 50 μM dNTPs, 1 μM of each primer, and 2 μl DNA extract (25 ng/ml) in a total volume of 50 μl. The reaction parameters were 2 min denaturation at 94°C followed by 30 cycles at 94°C for 2 min, 55°C for 1 min, and 72°C for 1 min, and a final extension at 72°C for 10 min as described by Nagachinta and Chen. 29 The PCR amplification products were separated by electrophoresis on a 1% SeaKem LE agarose gel in 0.5 × TAE buffer, visualized by ethidium bromide staining and photographed under UV light.
Determination of integron class
Integron-positive samples were further analyzed to identify the integron class (class I, class II, or class III) by restriction fragment length polymorphism as described by White et al. 30 Briefly, PCR products amplified with hep35/hep36 primers from the conserved regions of the integrase genes were digested (37°C, 3 hr) with HinfI (New England Biolabs, Ipswich, MA). Digestion products were separated by electrophoresis (100 V) on a 2% SeaKem LE agarose gel in 0.5 × TAE buffer, visualized by ethidium bromide staining and photographed under UV light.
Afterward, the assigned integron classes were confirmed by PCR with class-specific primers, IntI1-F and IntI1-R for integron class I and IntI2-F and IntI2-R for integron class II. 31 The composition of the PCR mix was similar as the one described above. The PCR parameters for integron class I were 3 min denaturation at 94°C, followed by 35 cycles of 1 min at 94°C, 30 sec at 68°C, 1 min at 72°C, and a final extension for 7 min at 72°C. The ones for integron class II were 5 min denaturation at 94°C, followed by 30 cycles of 30 sec at 94°C, 30 sec at 62°C, 1 min at 72°C, and a final extension for 8 min at 72°C. The PCR amplification products were separated and visualized as described above.
Antibiotic resistance transfer
The transfer of cefotaxime (FOT) and ciprofloxacin (CIP) resistance was analyzed by liquid mating. The recipient strain was E. coli K12 MG1655, which was first made rifampicin (RIF) resistant by serial inoculation on Tryptic Soy Agar plates with increasing concentrations of rifampicin (4–128 mg/L). Donor (50 strains, FOTR and/or CIPR) and recipient (E. coli K12 RIFR) strains were each inoculated on Luria Bertani (LB) plates without any antibiotics and incubated overnight at 37°C. The next day 1 colony of each strain was transferred to 5 ml LB broth containing the selective antibiotic (50 mg/L rifampicin for the recipient strain, 0.25 mg/L cefotaxime or 0.064 mg/L ciprofloxacin for the donor strains) and incubated by shaking (150 rpm) at 37°C. After 16 hr the strains were washed thrice with LB. Subsequently the strains were 10-fold diluted in LB, and liquid mating was performed by adding 300 μl of a donor strain to 1,500 μl of the recipient strain in a glass test tube. All tubes were incubated by shaking (150 rpm) at 37°C for 23 hr. After incubation, cultures were diluted and plated on selective LB plates. For the mating experiments with cefotaxime resistant donor strains this was LB with rifampicin (50 mg/L) and cefotaxime (0.25 mg/L) for quantification of the transconjugants, LB with rifampicin for the recipients, and LB with cefotaxime for the donors. For the ciprofloxacin resistant donor strains this was LB with rifampicin (50 mg/L) and ciprofloxacin (0.064 mg/L) for the transconjugants, LB with rifampicin for the recipients, and LB with ciprofloxacin for the donors. After overnight incubation at 37°C, the colonies on the plates were counted. The detection limit was 10 CFU/ml (1 colony/100 μl), and the limit of quantification was set at 1.5 × 102 CFU/ml (≥15 colonies/plate were regarded as reliable for quantification). The transfer ratio was calculated by dividing the number of transconjugants by the number of recipients.
The reproducibility of the liquid mating technique was assessed by performing triplicate setups of six selected donor strains. Negative controls (donor strain MB 5876 without added recipient strain and vice versa) were included.
Statistical analysis
To elucidate associations between sets of two nominal variables (origin, phylogenetic group, integron class, and plasmid replicon type) a two-tailed Fisher's exact test with Bonferroni correction for multiple testing was performed. p-Values lower than 0.003 (after Bonferroni correction) were considered as significant.
Quantitative data were examined for normality by the Shapiro–Wilk test and Q-Q plots. Both ciprofloxacin and cefotaxime transfer ratios and the number of observed antibiotic resistances were not normally distributed. A nonparametric Kruskal–Wallis test by ranks with Bonferroni correction was conducted to reveal if the transfer ratios were different among phylogenetic groups, strain origins, integron classes, and plasmid replicon types. p-Values lower than 0.003 (after Bonferroni correction) were considered as significant.
The calculations were performed using the statistical software R version 3.3.3 (R Foundation for Statistical Computing, Vienna, Austria).
Results
Antimicrobial susceptibility profiles
The antimicrobial susceptibility profiles of the 50 selected E. coli strains were determined using the antibiotic panel (Table 1). Forty-five strains were resistant to five or more antibiotics. Among the most commonly observed resistances were ampicillin (100%), sulfamethoxazole, ciprofloxacin (both 82%), trimethoprim, tetracycline (both 74%), cefotaxime (70%), and ceftazidime (62%). Two strains (MB5900 and MB5917) isolated from broiler chickens were only resistant to two antibiotics, more specifically ciprofloxacin and ampicillin. In contrast, there were 2 strains (MB5903 and MB5916) originating from veal, which were resistant to 11 antibiotics (sulfamethoxazole, trimethoprim, ciprofloxacin, tetracycline, azithromycin, cefotaxime, nalidixic acid, chloramphenicol, ceftazidime, ampicillin, and gentamicin). All tested strains were susceptible to meropenem and tigecycline. There was only 1 strain in the collection (MB5918, originating from broiler and resistant to 10 antibiotics), which was resistant to colistin.
Resistant to ciprofloxacin but not to nalidixic acid, fresistant to cefotaxime, phylogenetic groups were assessed by quadruplex PCR 26 ; antimicrobial susceptibility profiles were determined by a microbroth dilution method (EUVSEC Sensititre™).
Plasmid replicon types were assessed by PCR as described by Carattoli 27 . Transfer ratios (No. of transconjugants/No. of recipients) were calculated after liquid mating (23 hr, 37°C in LB broth) with an Escherichia coli K12RIFR acceptor strain. n.d. Strain failed to grow on selective plates. Integron classes were assigned by PCR with class specific primers and confirmed by HinfI restriction digest of the conserved regions of the integrase gene.
Gel electrophoresis of the class II specific PCR amplicon resulted in an intense band between 1,636 and 2,000 bp instead of the expected 746 bp band. HinfI restriction digest of the conserved regions of the integrase gene gave a normal banding pattern, indicative for the presence of class I and class II integrons.
AMP, ampicillin; AZI, azithromycin; CHL, chloramphenicol; CIP, ciprofloxacin; COL, colistin; FOT, cefotaxime; GEN, gentamicin; LB, Luria Bertani; MER, meropenem; MIC, minimum inhibitory concentration; NAL, nalidixic acid; n.d., not detected; R, resistant (i.e., MIC > ECOFF value); S, susceptible; SMX, sulfamethoxazole; TAZ, ceftazidime; TET, tetracycline; TGC, tigecycline; TMP, trimethoprim.
Phylogenetic groups
The phylogenetic group was determined using a quadruplex and two duplex PCR (Table 1). Table 2 presents the distribution of the phylogenetic groups per animal species. Phylogenetic group A and B1 were the most common phylogenetic groups among the analyzed strains. In each animal species only one strain belonged to phylogenetic group C.
Plasmid replicon typing
The presence of 20 plasmid replicons was investigated by means of 5 triplex PCRs and 5 simplex PCRs. Replicons of following plasmid incompatibility groups were not detected among the analyzed strains: A/C, FIIAS, HI2, L/M, T, W, and X. In 43 strains, more than one replicon was detected. In strain MB5933 six replicons were detected. The most common replicons were FII, FIB, and I1 (Fig. 1).

Distribution of the plasmid replicons determined by PCR-based replicon typing.
Integrons
Forty out of the 50 tested strains were positive for integrons (Table 1). All positive strains harbored a class I integron and seven of them contained an additional class II integron. No class III integrons were detected.
Antibiotic resistance transfer
The transferability of cefotaxime or ciprofloxacin resistances was determined by the reproducible liquid mating method (Fig. 2 and Table 1). In each mating 108 CFU of the recipient strain was added to 2.8 × 106–7.8 × 107 CFU of each cefotaxime resistant donor strain and 1.6–5.4 × 107 CFU of each ciprofloxacin resistant donor strain. There were five cefotaxime resistant strains that failed to grow on the selective LB plates. Hence, after mating, neither donors nor transconjugants were detected for these strains. For nine other strains, the number of transconjugants was below the limit of quantification. For the remaining 21 strains, the cefotaxime resistance transfer ratio, that is, the ratio of the number of transconjugants to the number of recipients, was in the order of magnitude of 10−5–100. A transfer ratio of 100 means that all recipients have gained the plasmid. Regarding the ciprofloxacin resistant strains, there were two strains for which the number of transconjugants was below the detection limit. For the other 14 strains, the transfer ratio was in the range of 10−7–10−1.
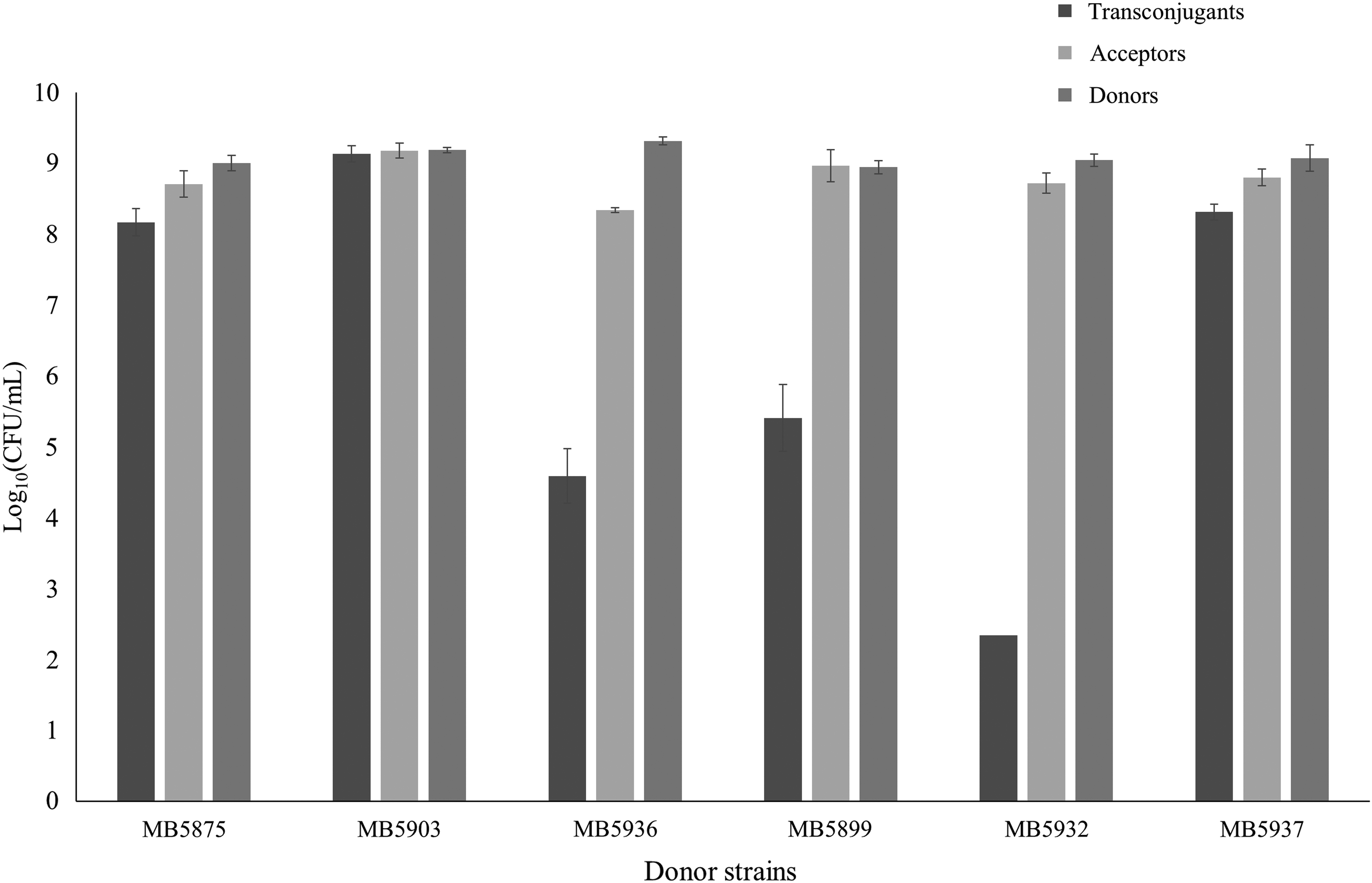
Reproducibility of the liquid mating method. Liquid mating was performed in LB at 37°C for 23 hr with various Escherichia coli donor strains and one acceptor strain (E. coli K12 RIFR). Mean ± standard deviation, n = 3. LB, Luria Bertani.
In the control setup in which the recipient (RIFR) was incubated without a donor strain, no cefotaxime resistant bacteria were detected. However, incubation of a donor strain (FotR) without recipient resulted in some rifampicin resistant bacteria (<1.5 × 102 CFU/ml), caused by spontaneous mutations (3.2 × 10−9 mutated donors/total number of donors).
Transfer ratios could not be linked to strain origin, phylogenetic group, integron classes, and plasmid replicon types (Kruskal–Wallis tests, p > 0.003 after Bonferroni correction).
Moreover, no significant associations could be found between strain origin, phylogenetic group, integron class, and plasmid replicon type (Fisher's exact test, p > 0.003).
Discussion
Antibiotics are widely used in food-producing animals for therapeutic treatment of clinically sick animals, for disease prophylaxis during periods of high risk of infection and for growth promotion. Several studies have demonstrated associations between antibiotic use in food-producing animals and the prevalence of antibiotic-resistant bacteria and resistance genes in those animals.3,32–37 As antibiotic use is regarded as an important driver of resistance selection and spread, its use as a growth promoter is forbidden in Europe since 2006. 38 Antibiotic resistant bacteria from food-producing animals can spread to humans through, for example, the food supply or direct contact with animals and through several indirect routes, for example, the use of contaminated manure as soil fertilizer.39,40 The current study characterized cefotaxime and/or ciprofloxacin resistant, commensal E. coli, previously isolated from food-producing animals and determined their resistance transfer potential.
Besides their cefotaxime and/or ciprofloxacin resistance, the majority of the strains were resistant to several other antibiotics, and six strains were even highly multiresistant, exhibiting resistance to 10 up to 11 antibiotics. The dominant resistances detected in the current study were those toward ampicillin, sulfamethoxazole, trimethoprim, and tetracycline. Penicillins, sulfonamides, and tetracyclines are among the most commonly used antibiotic compounds in food-production animals in Belgium. 41 Chantziaras demonstrated that the use of these antimicrobials strongly correlates with the level of resistance toward these agents found in commensal E. coli isolates in pigs, poultry, and cattle in Belgium. 32
Although these commensal strains are generally regarded as nonpathogenic for humans, presence of antibiotic resistance genes in these strains is very relevant as they might transfer their resistance genes to other (pathogenic) bacteria. The results of the liquid mating assay demonstrated that ciprofloxacin and cefotaxime resistance could be easily transferred; however, considerable variation in transfer ratio was observed between the tested strains (CIPR transfer ratio: 10−7–10−1 and FOTR transfer ratio: 10−5–100). This variation could not be explained by strain origin, phylogenetic group, integron class, or replicon type, but might be caused by the genetic background of the donor or plasmid related factors affecting transfer, for example, by interfering with the cell contact. Strain (and species) variation in transfer ratios despite identical mating conditions has also been observed by others.42–44
Its important to note that in the current study, the 2 strains with the highest number (11) of observed antibiotic resistances had some of the highest transfer ratios. However, no significant correlation between the number of antibiotic resistance genes and transfer ratios could be detected.
As there is extensive genetic substructure within the species E. coli, they can be assigned to phylogenetic groups. There is increasing evidence that strains of various phylogenetic groups differ in phenotypic and genotypic characteristics. 26 In general, B1, B2, A, and D are regarded as the most prevalent phylogroups. 45 In the current study the tested commensal E. coli mainly belonged to group A and B1. No members of group B2 and D were detected. While some authors claim that commensal strains predominantly belong to group A and B1,26,46 which is consistent with our results, this could not be confirmed by others. 47 These discrepant observations could be due to matrix (human vs. animal) and population differences and to different phylotyping methods. Apart from the phylogroup, the replicon types were determined for each strain. The replicon type of the plasmid determines if the plasmid can stably coexist with other plasmids in the recipient cell. Type FII and FIB were most prevalent, followed by I1. The first two replicon types are indicators of the presence of IncF plasmids. These plasmids are often detected in the Enterobacteriaceae and represent one of the most frequently reported plasmid types. F plasmids can contain multiple F replicons and, like I1 plasmids, have been described as narrow host range plasmids.48,49
Both IncF and IncI1 have been shown to be associated with the spread of antibiotic resistance genes, including extended-spectrum β-lactamase, quinolone, and aminoglycoside resistance genes.10,49 IncF plasmids express long flexible pili and are efficiently transferred in liquid cultures, whereas IncI plasmids express short rigid pili and are known to transfer more efficiently among cells on a surface. 50 The liquid matings performed in the current study did not allow to verify the enhanced transfer of IncF plasmids in liquid cultures as many strains harbored multiple plasmids and it was not determined which particular plasmid(s) was/were transferred.
Eighty percent of the E. coli strains contained integrons, which is high compared to other studies (e.g., 40%, 31 7.5%, 44 60%, 51 and 60.1% 52 ). This might be explained by differences in characteristics of the analyzed strain collection as in several studies; also antibiotic susceptible strains and/or pathogenic strains were taken into account. Our study exclusively tested cefotaxime/ciprofloxacin resistant commensal strains, mainly from broiler (70%) origin. The selection of cefotaxime/ciprofloxacin resistant strains may have favored the presence of integrons. This statement is supported by other studies which reported a high (66–97%) prevalence of integrons in multiresistant (incl. cefotaxime/ciprofloxacin) E. coli strains.23,53–55
In the current study, class I was the most abundant integron group, and no class III integrons were detected. This is in line with multiple other studies which demonstrate that class I integrons are most frequently encountered, followed by class II integrons and the rarely detected class III integrons.31,52,56 As integrons are known to be involved in the emergence of multiple antibiotic resistance it is not surprising that the 2 strains with the lowest amount (2) of observed antibiotic resistances did not have detectable integrons, and the 6 highly multiresistant strains (10 and 11 resistances) all carried class I integrons.
The results of the current study prove that commensal E. coli in food-producing animals can be a source of multiresistant bacteria and that some bacteria can easily spread their ciprofloxacin and cefotaxime resistance genes to recipients under certain tested in vitro conditions. Further research is needed to annotate the plasmids, to determine the location of antibiotic resistance genes and integrons, and to identify factors that enable plasmid maintenance and stability in the recipient. Moreover, the high antibiotic resistance transfer ratios observed in vitro in certain strains should be validated in human gastrointestinal environments to meet the need of quantitative data under circumstances more closely related to the actual situation of food consumption.
Lazarus et al. 57 concluded that there is evidence that a proportion of human extraintestinal infections caused by expanded-spectrum cephalosporin-resistant E. coli originates from food-producing animals, with poultry as the more likely source. They also denoted that there is a lack of knowledge regarding the specific parameters, such as the magnitude and geographical extent of the problem surrounding the transmission of bacteria and genetic elements from poultry to humans. In the current study, the majority of the cefotaxime and/or ciprofloxacin resistant strains were from broiler origin. As the number of tested strains per animal species was similar, this confirms that broiler chickens are an important source of resistant bacteria. It has been estimated that the probability to be exposed to more than 1,000 CFU of cephalosporin resistant E. coli by a prepared serving with broiler meat is about 1.5%. 58 The implications for human health are unknown, because the minimal infectious dose and the factors influencing the transfer of antimicrobial resistance genes from E. coli to the human intestinal bacterial community are not understood well enough.
The high number of antibiotic resistances and high transfer ratios demonstrated in commensal E. coli originating from Belgian farm animals in 2013 underscore that commensal E. coli in food production animals can be a reservoir and vector for antibiotic resistances.
Footnotes
Acknowledgments
Many thanks to Prof. A. Carattoli for providing the positive controls for the PCR-based replicon typing. The research that yielded these results was funded by the Belgian Federal Public Service of Health, Food Chain Safety and Environment through the contract [RF 13/9 TRAMRISK].
Disclosure Statement
All authors disclose no commercial associations that might create a conflict of interest in connection with this study.