
Abstract
Acinetobacter baumannii is an important pathogen of nosocomial infections. Nosocomial outbreaks caused by antibiotic-resistant A. baumannii remain a significant challenge. Understanding the antibiotic resistance mechanism of A. baumannii is critical for clinical treatment. The purpose of this study was to determine the whole-genome sequence (WGS) of an extensively drug-resistant (XDR) A. baumannii strain, XDR-BJ83, which was associated with a nosocomial outbreak in a tertiary care hospital of China, and to investigate the antibiotic resistance mechanism of this strain. The WGS of XDR-BJ83 was performed using single-molecule real-time sequencing. The complete genome of XDR-BJ83 consisted of a 4,011,552-bp chromosome and a 69,069-bp plasmid. The sequence type of XDR-BJ83 was ST368, which belongs to clonal complex 92 (CC92). The chromosome of XDR-BJ83 carried multiple antibiotic resistance genes, antibiotic efflux pump genes, and mobile genetic elements, including insertion sequences, transposons, integrons, and resistance islands. The plasmid of XDR-BJ83 (pBJ83) was a conjugative plasmid carrying type IV secretion system. These results indicate that the presence of multiple antibiotic resistance genes, efflux pumps, and mobile genetic elements is likely associated with resistance to various antibiotics in XDR-BJ83.
Introduction
A
The resistance mechanism of A. baumannii is complex and diverse. In addition to the inherent resistance genes, A. baumannii can increase the minimum inhibitory concentration (MIC) of antibiotics through rapid gene mutations or the transfer of exogenous resistance genes by mobile genetic elements, such as insertion sequences (ISs), transposons, or plasmids. 9 The A. baumannii genome can harbor a variety of resistance islands (RIs), which could be inserted into different target genes. Moreover, there is a special RI for A. baumannii that is characterized by interrupting the comM gene and designated A. baumannii resistance (AbaR).
With the rapid development of sequencing technology, many whole-genome sequences (WGSs) of A. baumannii strains have been obtained. To date, most of the reported WGSs of A. baumannii were obtained using next-generation sequencing; however, single-molecule real-time (SMRT) sequencing is increasingly applied. SMRT sequencing has the longest average read lengths and the highest consensus accuracy and is thus gradually becoming a mainstream technology for bacterial genome sequencing. Accumulation of a large number of WGSs and development of bioinformatics technology have made comparative genomics an effective method to study the resistance mechanism of A. baumannii.10–12
Our previous study described nine clones (clone A to I) of 60 A. baumannii isolates associated with a nosocomial outbreak at Chinese People's Liberation Army (PLA) General Hospital in Beijing, China, in 2007 by pulsed-field gel electrophoresis. 13 Clone A was the most common clone during this outbreak, and all clone A strains were MDR or extensively drug-resistant (XDR) A. baumannii. 8 We completely sequenced and annotated a clone A strain, designated XDR-BJ83, by SMRT sequencing to explore the genome characteristics and genomic basis of antibiotic resistance of this strain.
Materials and Methods
Ethics statement
All protocols associated with collection and storage of the A. baumannii strain XDR-BJ83 were approved by the Hospital Review Board of the Chinese PLA General Hospital. Written consent was obtained from the patient for his information to be used for medical research.
Bacterial strain
The clinical A. baumannii strain XDR-BJ83 was obtained from the Chinese PLA General Hospital in Beijing, China. This strain was isolated from a bile sample of a 66-year-old male patient with obstructive jaundice and biliary tract infection in July 2007. XDR-BJ83 was considered to be the pathogen that caused the biliary tract infection. The species was identified using the VITEK-2 system (BioMérieux, Marcy I'Étoile, France) in the clinical microbiology laboratory.
Antibiotic susceptibility testing
Susceptibility testing for XDR-BJ83 was performed using Etest strips (BioMérieux) according to recommendations proposed by the Clinical and Laboratory Standards Institute. The tested antibiotics included ampicillin, piperacillin/tazobactam, cefepime, ceftriaxone, cefoperazone/sulbactam, meropenem, imipenem, levofloxacin, ciprofloxacin, amikacin, gentamicin, minocycline, tigecycline, colistin, and trimethoprim/sulfamethoxazole.
Genome sequencing and assembling
The genomic DNA of XDR-BJ83 was extracted using the TIANamp Bacteria DNA Kit (Tiangen Biotech Co. Ltd., Beijing, China) according to the manufacturer's instructions. The concentration and purity of extracted DNA were tested using a spectrophotometer to ensure that the concentration was higher than 150 ng/μl, specifically OD260/OD280 was within 1.8 to 2.0 and OD260/OD230 within 2.0 to 2.2. The length of DNA fragments was confirmed to be longer than 15 kb by agarose gel electrophoresis. A 5- to 10-kb DNA library was constructed using the SMRTbell™ Template Prep Kit 1.0 (Novogene Bioinformatics Institute, Beijing, China). Then, the library was deposited onto an SMRT cell with P6/C4 chemistry and sequenced using the PacBio RS II platform (Pacific Biosciences, Menlo Park, CA) by a commercial vendor. The PacBio reads were de novo assembled using the SMRT Analysis pipeline v2.3.0 (Pacific Biosciences).
Genome annotation
Gene prediction and annotation of the assembled genome were performed using the NCBI prokaryotic genome annotation pipeline (PGAP). 14 The PGAP used GeneMarkS+ 4.0 for identification of protein-coding sequences, 15 RNAmmer for rRNA genes, 16 tRNAscan-SE for tRNA genes, 17 and Rfam and Cmsearch for small ncRNA genes.18,19 CRISPRFinder was chosen to identify the clustered regularly interspaced short palindromic repeats (CRISPRs). 20 PHAST was used to identify prophages. 21 IS Finder was used to characterize ISs. 22 Functions of genes were identified by comparing with the NCBI nonredundant protein database, and the functional classification of these genes was performed by aligning to the cluster of orthologous group (COG) database. 23 All of the protein-coding sequences were uploaded to the Comprehensive Antibiotic Resistance Database (CARD) for identifying antibiotic resistance genes by online BLASTp search. 24 ResFinder of Center for Genomic Epidemiology (CGE) was used to confirm the resistance genes. 25 Drug transporter and efflux pump genes were annotated by using the Transporter Classification Database. 26 Genomic Island Prediction software (GIPSy) was used to characterize RIs. 27 The comM gene of XDR-BJ83 was examined to identify the AbaR-type RI. Gene sequence alignment with other strains was performed using NCBI BLAST online.
Multilocus sequence typing
Multilocus sequence typing (MLST) of XDR-BJ83 was performed according to both the Oxford scheme and the Pasteur scheme. The FASTA sequence of the predicted genes was uploaded to the CGE server, and then allelic profiles and STs were generated.
Phylogenetic analysis
The genome data of reference strains used for phylogenetic analysis were downloaded from the FTP site of NCBI, including the A. baumannii strains, AB0057 (NC_011586.2), Ab04-mff (NZ_CP012006.1), AB307-0294 (NC_011595.1), AC29 (NZ_CP007535.2), AC30 (CP007577.1), ACICU (NC_010611.1), AYE (NC_010410.1), BJAB07104 (NC_021726.1), BJAB0715 (NC_021733.1), BJAB0868 (NC_021729.1), D36 (NZ_CP012952.1), DU202 (NZ_CP017152.1), LAC-4 (NZ_CP007712.1), KBN10P02143 (NZ_CP013924.1), MDR-TJ (NC_017847.1), MDR-ZJ06 (NC_017171.1), SDF (CU468230.2), TCDC-AB0715 (CP002522.2), TYTH-1 (NC_018706.1), XH386 (NZ_CP010779.1), and YU-R612 (NZ_CP014215.1). SDF was used as an outgroup strain for comparison, which was isolated from human body louse. The sequences of seven housekeeping genes (gltA, gyrB, gdhB, recA, cpn60, gpi, and rpoD) used in the Oxford MLST scheme of these strains were extracted and then aligned by using ClustalW. 28 The phylogenetic tree was constructed according to these aligned housekeeping genes using MEGA7 with the neighbor-joining method. 29
GenBank accession numbers
The genome sequence of XDR-BJ83 has been deposited in GenBank under the accession numbers CP018421 (chromosome) and CP018422 (plasmid).
Results
Antibiotic susceptibility profiles
The MICs of 15 tested antibiotics are shown in Table 1. XDR-BJ83 was resistant to penicillins, cephalosporins, carbapenems, fluoroquinolones, aminoglycosides, and sulfonamides. It was only susceptible to polymyxins. The MICs of this strain to minocycline and tigecycline were 8 and 4 μg/ml, respectively, which are intermediate values.
MIC, minimum inhibitory concentration; XDR, extensively drug-resistant; R, resistant; I, intermediate; S, susceptible.
Genome sequence and general features
A total of 29,383 clean reads were generated with an average length of 8,915 bp, resulting in a mean coverage of 64 × (sequence depth). After de novo assembly, the complete genome of XDR-BJ83 yielded a 4,011,552-bp circular chromosome with 39.0% G + C content and a 69,069-bp plasmid (designated pBJ83) with 33.8% G + C content (Fig. 1). The general characteristics of XDR-BJ83 genome are summarized in Table 2. We predicted 3891 potential protein-coding sequences from the genome of XDR-BJ83, and 1015 (26.1%) of these genes encode hypothetical proteins. According to the COG annotation, 2858 (73.5%) of these genes were annotated and classified into 23 COG terms.
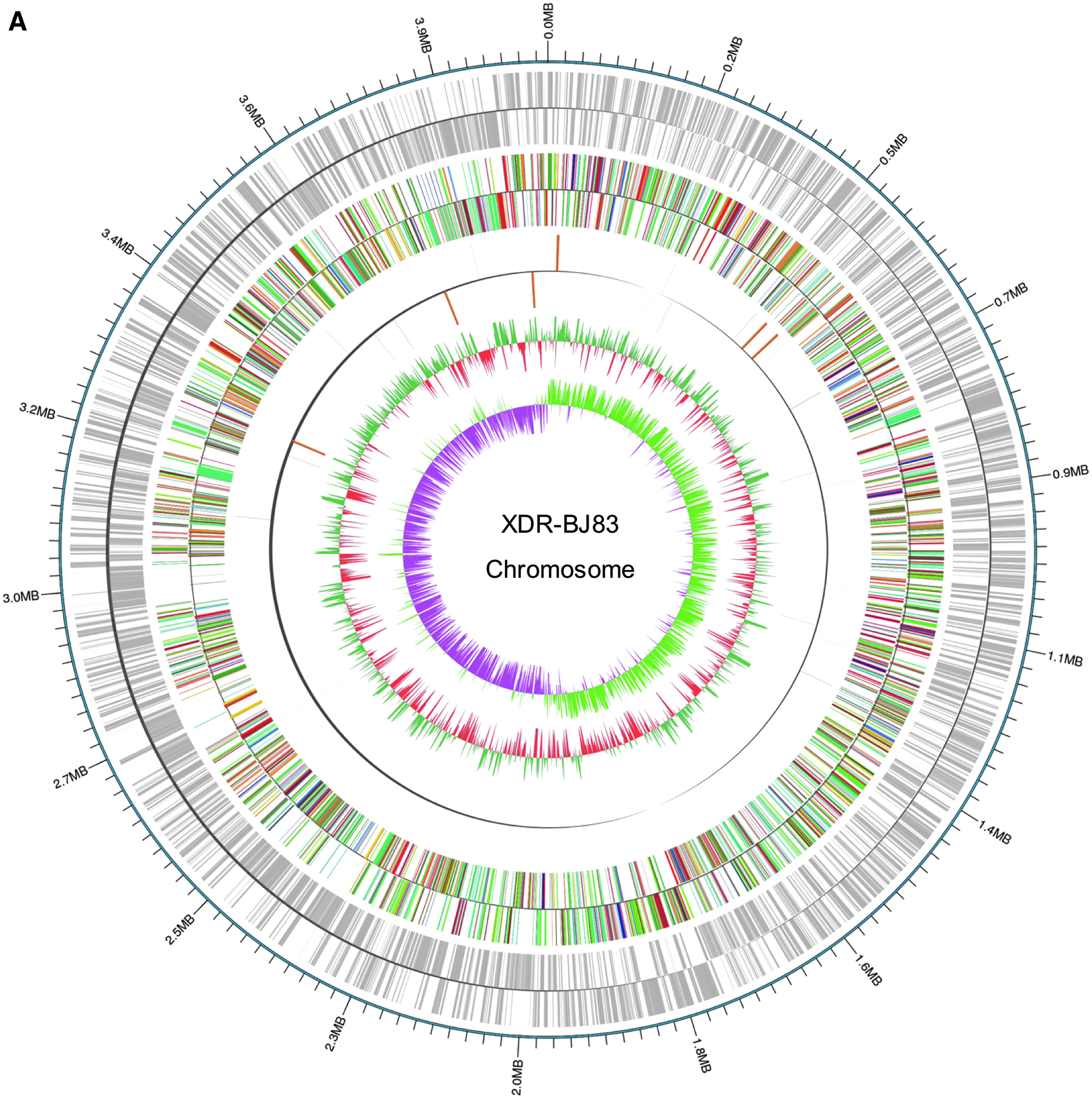
Circular genome representation of the Acinetobacter baumannii strain XDR-BJ83 chromosome (
CRISPR, clustered regularly interspaced short palindromic repeat.
MLST and phylogenetic analysis
The sequence type of XDR-BJ83 was ST368 (1-3-3-2-2-140-3) under the Oxford MLST scheme, which is classified as clonal complex 92 (CC92). Under the Pasteur MLST scheme, the sequence type was ST2 (2-2-2-2-2-2-2), which belongs to the largest and most widespread group, International Clone-II (IC-II).
Based on the seven housekeeping genes of the Oxford MLST scheme, the phylogenetic tree revealed that XDR-BJ83 was closest to the A. baumannii strain BJAB07104, which is also an ST368 strain. These two strains were clustered together with many other strains, such as BJAB0868, DU202, KBN10P02143, AC29, AC30, MDR-TJ, MDR-ZJ06, YU-R612, TCDC-AB0715, XH386, TYTH-1, ACICU, and D36, which all belong to group IC-II.
Genes related with antibiotic resistance
A total of 19 genes related to antibiotic resistance were found in the genome of XDR-BJ83 (Table 3). The cephalosporinase (class C β-lactamase), encoded by blaADC of XDR-BJ83, was a new variant, and the gene sequence showed a nucleotide mutation (A1022C) when compared with blaADC-25 (EF016355.1). A copy of ISAba1 was located upstream of the blaADC gene, but no IS existed upstream of blaOXA-66, which is also an intrinsic β-lactams gene. Five copies of the blaOXA-23 gene were found and each of them has an ISAba1 in their upstream sequence. Mutation in the gyrA gene (Ser83Leu) could explain the resistance profile to fluoroquinolones. For the parC gene, there were three mutations (Met32Ile, Glu88Gly, and Ser467Gly) when compared with the antibiotic-sensitive strain SDF. All resistance genes were located on the chromosome of XDR-BJ83, and the plasmid pBJ83 did not carry a resistance gene.
Drug efflux pumps also play important roles in antibiotic resistance. Five major efflux pump families, including resistance-nodulation-division family (RND), major facilitator superfamily (MFS), multidrug and toxic compound extrusion family (MATE), small multidrug resistance family (SMR), and ATP-binding cassette superfamily (ABC), are considered to be associated with A. baumannii antibiotic resistance. 30 In Table 4, the known drug efflux pump genes are listed. In addition, there were many putative drug efflux pump genes.
ABC, ATP-binding cassette superfamily; MATE, multidrug and toxic compound extrusion family; MFS, major facilitator superfamily; RND, resistance-nodulation-division family; SMR, small multidrug resistance family.
The two-component regulatory genes of XDR-BJ83, adeR and adeS, are adeABC operon regulatory genes that were structurally intact and had no IS in their upstream sequences. The amino acid sequences of AdeR and AdeS were compared with the antibiotic-sensitive strain AB307-0294, 31 indicating the presence of multiple mutations (Fig. 2). In the genome of XDR-BJ83, the target site of the regulator protein AdeR was a 10-bp direct repeat sequence with the base T interval (AAGTGTGGAGTAAGTGTGGAG) that was located in the intergenic region between the adeR gene and adeABC operon, and no mutation was observed.

Alignment of AdeR
The XDR-BJ83 genome carried a variety of genes encoding outer membrane proteins (OMPs), such as carO, ompA, ompW, omp33, omp25, dcap-like, oprB, and oprD. Interruption of the carO gene is associated with carbapenem resistance.32,33 However, the carO gene carried in the XDR-BJ83 genome was intact and the amino acid sequence showed 100% identity with CarOb, which has a high affinity for carbapenems (Fig. 3).

Alignment of amino acid sequences of CarO from A. baumannii strain XDR-BJ83 with CarOa (AKL79738), CarOb (ADQ27797), and CarO-undefined (AFH56947), which were obtained from GenBank. The asterisk represents the same amino acid as the reference sequence.
Insertion sequences
In the genome of XDR-BJ83, 8 types of ISs were identified, and a total of 31 ISs were annotated (Table 5). Four types of these ISs belong to A. baumannii, and the other four types are thought to originate from other bacterial species. The number of ISAba1 was the largest in these ISs (21, 67.7%). IS26 and IS15DI all belong to the IS6 family, and there are only 3-bp differences for their nucleotide sequence. The sequence between 1333264 and 1334083 bp was 100% identical with IS26, and the sequence between 1333264 and 1334083 bp was 100% identical with IS15DI. However, sequences between 1331378 and 1332197 bp and 1351517 and 1352336 bp had 2-bp differences with IS26 and 1-bp difference with IS15DI, respectively. In this study, we classified these four ISs as IS26 according to previous studies. 34 The positional relationship between ISs and antibiotic resistance genes will be shown in the next section.
IS, insertion sequence.
Structures of resistance islands
Five putative resistance islands (PRIs) were predicted on the chromosome of XDR-BJ83 using GIPSy. No resistance island was found on the plasmid pBJ83. PRI3 and PRI5 carried several resistance genes. PRI1, PRI2, and PRI4 carried many genes with unclear function (hypothetical genes) and did not carry any resistance genes. The atr gene, encoding N-acetyltransferase, was interrupted by PRI3 with an 8-bp direct repeat sequence formation (Fig. 4). The main structure of PRI3 comprised Tn6179 and Tn6180, which contained five copies of ISs, including a copy of ISEc28, a copy of ISEc28, and three copies of IS26. Tn6180 carried a copy of class I integron, which includes three gene cassettes possessing aacA4, catB8, and aadA1 genes. PRI5 was inserted into the znuA gene, which encoded the zinc ABC transporter solute-binding protein and formed a 9-bp direct repeat sequence (Fig. 4). PRI5 possessed five copies of Tn6206, which carried a copy of ISAba1 and the blaOXA-23 gene. The truncated genes and direct repeat sequences suggested that PRI3 and PRI5 were acquired by transposition.
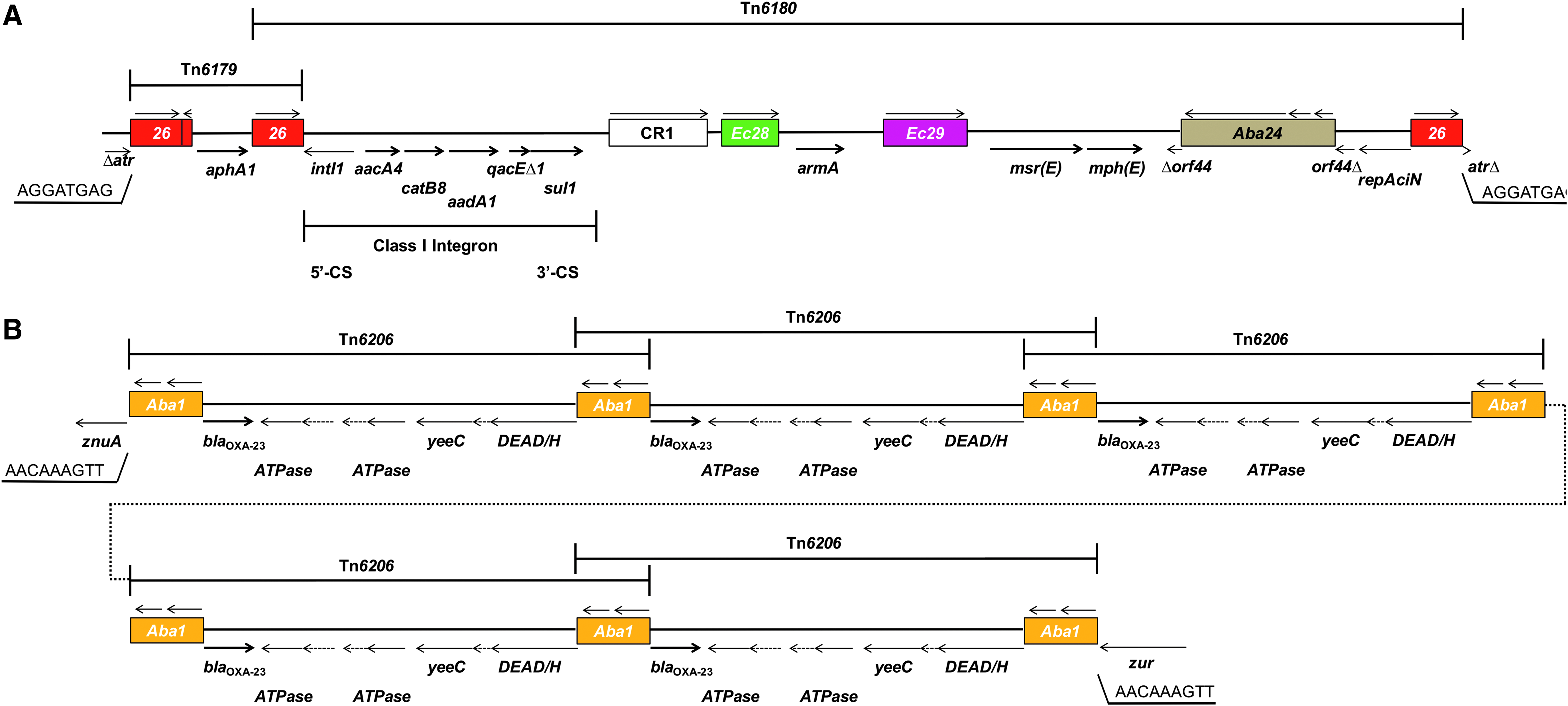
Structures of PRI3
In addition to the above PRIs, the genome of XDR-BJ83 contained an AbaR-type RI (designated AbaRXDR-BJ83) (Fig. 5). The AbaRXDR-BJ83 was 43,080 bp in size and carried two transposons (Tn6022 and Tn6022▵), two copies of ISAba1, and four resistance genes (sul2, tetA(B), strA, and strB). The other ISs mentioned in the previous section were not located in any PRI or adjacent to any antibiotic resistance gene.
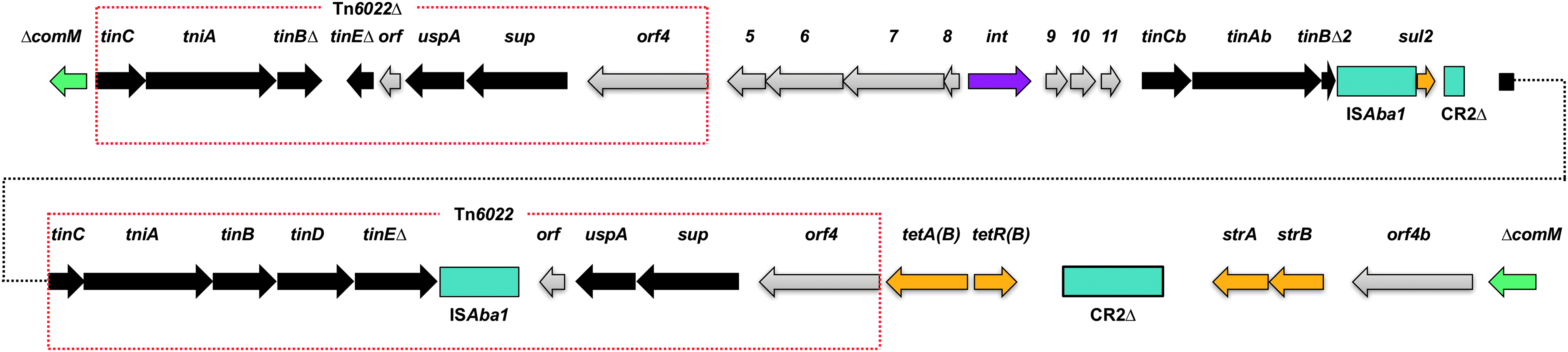
Structure of AbaRXDR-BJ83 in the A. baumannii strain XDR-BJ83 genome. Arrows represent the extent and orientation of genes. Among them, green arrows represent truncated comM gene, blue arrows represent transposase genes, yellow arrows represent resistance genes, purple arrows represent integrase genes, and gray arrows represent genes that encode the hypothetical protein. These genes encoding hypothetical proteins were drawn based on the previous literature. 45 Colored boxes represent ISs or ISCRs. Red dotted-line boxes indicate the range of transposons.
Type IV secretion system in plasmid pBJ83
Plasmid pBJ83 was categorized as an F-plasmid because it contained a 33-kb region that carried 20 genes encoding the type IV secretion system (T4SS) apparatus. The 20 T4SS-related genes included traD, traI, finO, traM, traA, traL, traE, traK, traB, trbG, traV, traC, trbI, traW, traU, trbC, traN, traF, traH, and traG. Except for the finO gene, all the other T4SS-related genes were located in the negative strand. The replicase of plasmid pBJ83 could not be classified by the current replicon typing method.
Discussion
In this study, we performed a WGS of the A. baumannii strain XDR-BJ83 by SMRT sequencing, which was associated with an outbreak at the Chinese PLA General Hospital. The sequence type of XDR-BJ83 was ST368, and our unpublished study showed that this A. baumannii outbreak was mainly caused by ST368 strains. In previous studies, ST368 was the predominant sequence type in several hospitals in China.35–37 According to the phylogenetic analysis, we observed that another ST368 strain, BJAB07104, had the closest relationship with XDR-BJ83, which was isolated 5 months earlier than XDR-BJ83 in the same hospital. 38 Thus, the spread of ST368 A. baumannii strains in this hospital was further confirmed.
XDR-BJ83 was an XDR A. baumannii. A variety of antibiotic resistance and efflux pump genes carried by the genome were the basis of antibiotic resistance of XDR-BJ83. The chromosome of XDR-BJ83 contained several β-lactamase genes, but did not include genes for class A and class B β-lactamases. A copy of ISAba1 was inserted upstream of the blaADC gene as a promoter to enhance ADC's expression and might be associated with resistance to expanded-spectrum cephalosporins. Further investigation is needed to determine whether the mutation (A1022C) in the blaADC gene affects the cephalosporinase hydrolysis activity. The blaOXA-66 gene belongs to the blaOXA-51-like gene cluster, which has a low level of carbapenemase activity and contains no ISs in its upstream sequence in the XDR-BJ83 genome. As the most widespread carbapenemase in CRAB, OXA-23 is a class D β-lactamase that has a strong carbapenemase activity. 9 All of the upstream blaOXA-23 genes were inserted by a copy of ISAba1 in the genome of XDR-BJ83, and each blaOXA-23 gene and ISAba1 were identified as part of Tn6206. The ISAba1 is also a strong promoter of the blaOXA-23 gene. In addition, the efflux pump operons, adeABC and adeIJK, might be associated with the resistance to β-lactams in XDR-BJ83. Overexpression of AdeABC and AdeIJK could reduce the concentration of β-lactam drugs in A. baumannii, but no effect of AdeIJK on carbapenem resistance has been previously observed. 30 The mutations of adeABC operon regulatory genes, adeR and adeS, in XDR-BJ83 genome might affect the expression of AdeABC. Some OMPs may be associated with decreased susceptibility or resistance to carbapenems, such as CarO, Omp33, or OprD. 32 Loss of the carO gene or disruption by IS can lead to resistance to carbapenems. 33 However, for XDR-BJ83, CarO has no function in resistance to carbapenems.
Resistance of XDR-BJ83 to aminoglycosides may be dependent on the genes encoding aminoglycoside-modifying enzymes, 16s RNA methyltransferase, and extended-spectrum pump AdeABC and AbeM, shown in Tables 3 and 4. Mutations in gyrA and parC genes may produce resistance to fluoroquinolones. Among them, a mutation resulting in an Ser-83-Leu substitution in GyrA increases the MICs of fluoroquinolones and has been confirmed by previous studies. 39 Another quinolone resistance mechanism might involve active efflux by AdeABC, AdeIJK, AdeFGH, AbeS, and AbeM. These efflux pumps could pump out fluoroquinolones and result in low-level resistance to all of these antibiotics. 30 The tetA(B) gene carried by the XDR-BJ83 genome encoded the specific efflux pump protein of tetracycline, which could explain the resistance to tetracycline. No tetB gene could be found in XDR-BJ83 genome, and only active efflux by AdeABC, AdeIJK, and AdeFGH was not sufficient to cause resistance to tigecycline and minocycline.30,40 Resistance of XDR-BJ83 to chloramphenicol may depend on genes encoding the chloramphenicol-modifying enzymes, CraA, AbeM, and AbeS. 41 The phosphotransferase encoded by the mph(E) gene and the macrolide-specific efflux pumps encoded by the msr(E), macA, and macB genes could produce resistance to macrolides in XDR-BJ83. In addition, products encoded by the sul1 and sul2 genes in the XDR-BJ83 genome could lead to resistance to sulfonamides.
Except for intrinsic resistance genes and gene mutations, the main genomic basis for antibiotic resistance is acquired resistance genes, which are obtained by horizontal gene transfer mediated by mobile genetic elements. The XDR-BJ83 genome carries a variety of mobile elements. ISAba1 is the most common IS in the genome of A. baumannii and is a basic component of multiple RIs and transposons. ISAba1 can enhance the expression of downstream resistance genes and can insert into some specific target genes, such as the adeS gene, resulting in resistance to tigecycline. 42 The genome of XDR-BJ83 contained 21 copies of ISAba1, which might play an important role in the horizontal gene transfer of acquired resistance genes.
The XDR-BJ83 genome had three RIs that carry antibiotic resistance genes. PRI3 is an AbGRI3-type RI that was first identified in the MDR-TJ and TYTH-1 strains, consists of only one copy of Tn6180, and is named as AbGRI3-1. A similar structure was also found in the YU-R612 and NCGM_237 strains.43,44 A new class of AbGRI3-type islands consists of Tn6180 and Tn6179, is designated AbGRI3-2, and was found in the UH9907 strain. 34 The structure of PRI3 was almost the same as that of AbGRI3-2. PRI5 was formed by five copies of Tn6206, which carries a copy of the blaOXA-23 gene and is the most common transposon in CRAB. In a previous study, the Tn6206 in the BJAB07104, BJAB0868, and BJAB0715 strains was different in target genes and direct repeat sequences. The Tn6206 in PRI5 also showed the same differences in comparison with the abovementioned strains. 38 The first AbaR-type RI was found in the AYE (designated AbaR1) strain and was characterized as inserted into the comM gene. 45 To date, there have been dozens of AbaR-type RIs in the NCBI database, and some do not carry resistance genes. The structure of AbaRXDR-BJ83 is similar to that of the MDR-ZJ06 (AbaR22) and MDR-TJ (RIMDR-TJ) strains. 46
The plasmid pBJ83 was a conjugative plasmid with a type IV secretion system and did not carry any resistance genes, ISs, transposons, or RIs. T4SS is a multisubunit bacterial cell membrane structure that is involved in a mechanism of conjugation that can transfer nucleoproteins and proteins across the bacterial cell membrane. 47 Currently, there have only been a few reports on the T4SS of A. baumannii. The T4SS of pBJ83 was classified as type A and was previously identified in the plasmid of the ACICU, TYTH-1, and KBN10P02143 strains. 48
In conclusion, the complete genome sequence of an ST368 A. baumannii strain, XDR-BJ83, which was associated with a nosocomial outbreak, was determined by SMRT sequencing. The XDR-BJ83 genome carried multiple antibiotic resistance genes, antibiotic efflux pump genes, and mobile genetic elements that might be associated with antibiotic resistance acquired by horizontal gene transfer. This work may lay the foundation for the future study of resistance mechanisms of A. baumannii.
Footnotes
Acknowledgments
This research was supported by the Key Project of Military Logistics Research (grant number: BWS14J040). The authors would like to thank Guoxin Mo, PhD, Department of Respiratory Medicine, Chinese PLA General Hospital, for providing the A. baumannii strain XDR-BJ83.
Disclosure Statement
No competing financial interests exist.