
Abstract
Objectives:
Mupirocin is a useful antibiotic against superficial skin infections. We compared the impact of mupirocin with a cephalosporin, a fluoroquinolone, an aminoglycoside, and a macrolide on planktonic cell growth and biofilm formation of methicillin-susceptible Staphylococcus aureus (MSSA) and methicillin-resistant S. aureus (MRSA).
Materials and Methods:
Minimum inhibitory concentration (MIC) of mupirocin was determined against S. aureus strains used in this study. Biofilm formation of S. aureus strains exposed to mupirocin was quantified by crystal violet staining assay. Moreover, biofilm structure and viability of the biofilm cells were visualized by Live/Dead staining assay. Biofilm-related gene expression was investigated by quantitative real-time PCR.
Results:
MRSA USA300 clone was resistant to mupirocin with MIC of 1,024 mg/L, while MRSA ATCC-43300 and MSSA ATCC-29213 were susceptible with MICs of 0.03 mg/L. Planktonic cell growth of the S. aureus strains was inhibited by mupirocin in a dose-dependent manner. However, some of the low concentrations of mupirocin less than the MICs promoted biofilm formation. Confocal laser scanning microscopy of the biofilm structures and cell viabilities showed established biofilms of slightly higher cell density in the mupirocin treated groups, especially in the MRSA USA300 clone. Gene expression of RNAIII in planktonic cells and biofilms of MRSA USA300 clone showed the highest upregulation after initial exposure to sub-MIC of mupirocin followed by downregulation, whereas the other antibiotics showed various fluctuations.
Conclusion:
The results showed that subinhibitory concentrations of mupirocin promoted biofilm formation of S. aureus, in particular the MRSA USA300 clone.
Introduction
S
Generally, bacteria exist in either of two states as planktonic or adherent cells. Planktonic or free-floating cells are important for proliferation and expansion, whereas adherent cells are bacteria colonized on biotic or abiotic surfaces. Bacteria are embedded within an extracellular polymeric substance matrix that plays an essential role in the initial forming of complex differentiated communities called biofilms. 3 Biofilms can survive in a hostile environment by resisting some antibiotics through innate properties or adapting to the defense mechanisms of the human body. In addition, biofilms give rise to chronic bacterial infections if the adherent cells are not removed. 4
Prevention and treatment of biofilm infections with topical or systemic antibiotics can be used in some biofilm infections. 5 Mupirocin or pseudomonic acid A is a fermentation product of Pseudomonas fluorescens. The mechanism of action is protein synthesis inhibition in bacteria by binding to the isoleucyl-tRNA synthetase enzyme that prevents the incorporation of isoleucine into protein. This mechanism has no in vitro incidence of cross-reactivity with other antimicrobial agents. 6 Mupirocin is highly effective against aerobic Gram-positive cocci such as S. aureus- and S. epidermidis–associated wound infections, including MRSA. The indications are for prophylaxis and treatment in ulcers, operative or burn wounds, skin infection, and eradication of nasal carriage S. aureus. 7
Few studies have demonstrated the efficiency of mupirocin against S. aureus or Pseudomonas aeruginosa bacterial biofilm infection in single or combination therapy8,9 even in subinhibitory concentrations of mupirocin against P. aeruginosa.10,11 However, when bacteria are exposed to supra-inhibitory concentrations of antibiotics followed by sub-minimum inhibitory concentrations (MICs) in vivo, the bacteria appear as antibiotic-resistant mutants. 12 Several studies have shown that subinhibitory concentrations of antibacterial substances modulated the biofilm growth.13–16 Moreover, antiseptic agents such as ethanol synergized the antibacterial activity of the formulations and are well known for their applications such as stabilizers, solvents, or disinfectants. However, a wide range of ethanol concentrations in different applications might eventually result in low levels of ethanol, which have also been reported to affect the viability and growth of biofilm development.17–19
Therefore, the objectives of this study were to investigate bacterial biofilm formation of S. aureus in response to various concentrations of mupirocin compared with ethanol by evaluation of drug susceptibility testing, quantification of biofilm formation, visualization of biofilm structure and viability of biofilm cells, and determination of biofilm gene expression compared to four other classes of antibiotics.
Materials and Methods
Bacterial strains and chemicals
S. aureus strains (MRSA ATCC BAA-1556 or USA300 clone, MRSA ATCC-43300, MSSA ATCC-29213) were purchased from American Type Culture Collection (ATCC, Manassas, VA). Bacteria were routinely cultured in brain heart infusion (BHI) agar (BD Diagnostics, NJ) at 35°C for 12 hr and then a single colony was inoculated into BHI broth and cultured for 12 hr. Mupirocin was obtained from TEVA Pharmaceutical Works Private Ltd., Co. (Petah Tikva, Israel). Cefazolin, levofloxacin, gentamicin, and erythromycin were purchased from Sigma-Aldrich (St. Louis, MO).
Mupirocin preparation
Concentrations of mupirocin from 0.001 to 1,024 mg/L were prepared by dissolving the drug in ethanol and diluting by tryptic soy broth (BD Diagnostics) supplemented with 0.5% (w/v) glucose (TSBG) medium. As a control group without mupirocin, ethanol was diluted by TSBG medium to detect the possible effects of ethanol on planktonic cell growth and biofilm formation.
Determination of mupirocin MIC
One hundred microliters of serial-diluted mupirocin and ethanol solutions were added to a 96-well round bottom polystyrene plate. For bacterial preparation, S. aureus strains were cultured at 35°C for 12 hr in BHI agar. The bacterial colonies were then suspended in 4 ml of normal saline solution (NSS). The turbidity of bacterial suspensions was adjusted to 0.5 McFarland using a nephelometer (PhoenixSpec; BD Diagnostics) and then diluted 10 times with NSS. Finally, 3 μl of bacterial suspensions were inoculated into the wells containing sample solutions and incubated at 35°C for 18 and 24 hr. Bacterial growth in the wells was visually observed and compared with a positive control. The lowest concentration which was able to inhibit bacterial growth was identified as the MIC.
Quantification of biofilms
S. aureus strains incubated in BHI broth were washed twice in TSBG medium, and the bacterial suspensions were diluted to an OD600 of 0.02. Fifty microliters of serial-diluted solutions of mupirocin and ethanol were added to a 96-well flat bottom polystyrene plate followed by 50 μl of S. aureus cultures. The plates were then incubated at 35°C for 6 hr. Following incubation, OD595 of bacterial cultures was measured by a microplate reader for the analysis of planktonic cell growth. The plates were then washed twice with 150 μl of deionized water (DW) to remove planktonic cells and stained with 150 μl of 0.2% (w/v) crystal violet solution for 30 min. The solutions were then discarded, and the stained biofilms were washed twice with 150 μl of DW and air-dried for 30 min. Finally, the biofilms were solubilized by 150 μl of absolute ethanol for 30 min before 100 μl was used to measure A595 for the analysis of biofilm formation.
Visualization of biofilm cell viability
Biofilm structures and the viability of biofilm cells were visualized using a confocal laser scanning microscope (LSM 710; Carl Zeiss, Jena, Germany) equipped with a 63x/1.40 numerical aperture Plan-Apochromat objective. One hundred microliters of the overnight cultures of S. aureus strains diluted to an OD600 of 0.02 were added into an eight well cover glass chamber (IWAKI, Tokyo, Japan) together with 100 μl of mupirocin and ethanol solutions. The chambers were then incubated at 35°C for 6 hr. Following incubation, biofilms were stained using a FilmTracer™ LIVE/DEAD® Biofilm Viability Kit (Invitrogen, Carlsbad, CA). After washing with DW, the SYTO9-stained cells (green) and propidium iodide-stained cells (red) in biofilms were excited with an argon laser and a helium-neon laser, respectively. Green and red fluorescence were detected within the range of 493–580 and 548–797 nm, respectively. Live/Dead image stacks of biofilms were captured at the same horizontal level of 30 μm height by a Carl Zeiss software (ZEN 2010). Moreover, the live biomass that occupied the biofilms in layers was calculated using a computer program for quantitative analysis (COMSTAT2; www.comstat.dk). 20
Measurement of gene expression
RNA extraction from planktonic cells and biofilms of MRSA USA300 clone was performed using the RNeasy Mini Kit (Qiagen, Hilden, Germany). Fifty microliters of the overnight cultures of MRSA USA300 clone were diluted to an OD600 of 0.02 and then added into a 96-well flat bottom plate together with 50 μl of 1/16x MIC of four antibiotics against MRSA USA300 clone. The antibiotics were mupirocin (64 mg/L), cefazolin (0.125 mg/L), levofloxacin (0.25 mg/L), gentamicin (0.0625 mg/L), and erythromycin (4 mg/L). The 96-well plate was then incubated at 35°C for 3, 4, 5, and 6 hr. After incubation for a specific time, the planktonic cell cultures were transferred to a 2 ml tube, and 0.1 ml of RNAprotect (Qiagen) was then added to the wells. Biofilms that formed in the wells were scraped by pipetting. Following vortex and incubation for 5 min, the scraped biofilms were transferred to a 2 ml tube. In the planktonic cell cultures, 0.5 ml of the transferred cultures were added to a 2 ml tube with 1 ml of RNAprotect followed by vortexing and incubation for 5 min. RNAprotect mixtures were centrifuged at 5,000 g for 10 min. After the supernatants were removed, the pellets were kept at −80°C. The pellets were lysed using 0.1 ml of Tris-EDTA buffer supplemented with 0.2 mg/ml of lysostaphin and then incubated at 37°C for 30 min. After extraction with RNeasy Mini Kit, RNA samples were treated with TURBO DNase (Ambion, Darmstadt, Germany). RNA concentration was determined using a BioSpec-nano (Shimadzu, Tokyo, Japan). cDNA was prepared using the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA) according to the protocol. Finally, quantitative real-time PCR was performed to measure the expression of agrA, RNAIII, and sarA genes using an Applied Biosystems 7500 Fast Real-Time PCR System. The primers are shown in Table 1. cDNA samples were treated with the Fast SYBR Green Master Mix (Applied Biosystems). The PCR condition was 95°C for 20 sec and then 40 cycles at 95°C for 3 sec and 60°C for 30 sec. Ct values were calculated by a 7500 Fast software version 2.3. The data were analyzed using the ΔΔCt method. 16S rRNA gene was used as an internal standard.
Statistical analysis
All experiments were repeated at least thrice. Data are presented as mean ± standard deviation. The significance of differences was calculated using a Student t test with a p-value <0.05 considered as statistically significant.
Results
Antimicrobial activity of mupirocin against planktonic S. aureus cells
The MICs of mupirocin against S. aureus strains are presented in Table 2. The MRSA USA300 clone was resistant to mupirocin at an MIC of 1,024 mg/L at 18 hr and more than 1,024 mg/L at 24 hr, whereas mupirocin-susceptible strains of MRSA ATCC-43300 and MSSA ATCC-29213 showed potent antimicrobial activity of mupirocin at a lower MIC of 0.03 mg/L at 18 and 24 hr. Ethanol in a range of 0.0001–102.4 ml/L (0.00001–10.24% v/v) had no antimicrobial effect against S. aureus.
MIC, minimum inhibitory concentration; MRSA, methicillin-resistant Staphylococcus aureus; MSSA, methicillin-susceptible Staphylococcus aureus.
Effect of mupirocin and ethanol on planktonic cell growth and bacterial biofilm formation
Figure 1 shows the planktonic cell growth and biofilm formation of the tested S. aureus after exposure to mupirocin or ethanol at the exponential or log phase of bacterial growth (6 hr). The results showed that the growth of planktonic cells in MRSA strains was inhibited by mupirocin in a dose-dependent manner once the concentration approached the MIC. However, methicillin-susceptible S. aureus (MSSA) indicated that the inhibition of growth occurred only at the MIC or higher concentrations (Fig. 1A). Interestingly, it was found that some of the low concentrations of mupirocin less than MIC promoted biofilm formation compared to the ethanol group (Fig. 1B). Mupirocin at concentrations of 16–64 mg/L in MRSA USA300 clone significantly increased the biofilm, as well as mupirocin in MRSA ATCC-43300 at a concentration of 0.008 mg/L and MSSA ATCC-29213 at concentrations from 0.002 to 0.03 mg/L. Nevertheless, the antibiofilm effect of mupirocin against MRSA USA300 clone, MRSA ATCC-43300, and MSSA ATCC-29213 was observed at higher concentrations near or above MIC in the ranges of 256–1,024, 0.125–1, and 0.125–1 mg/L, respectively.
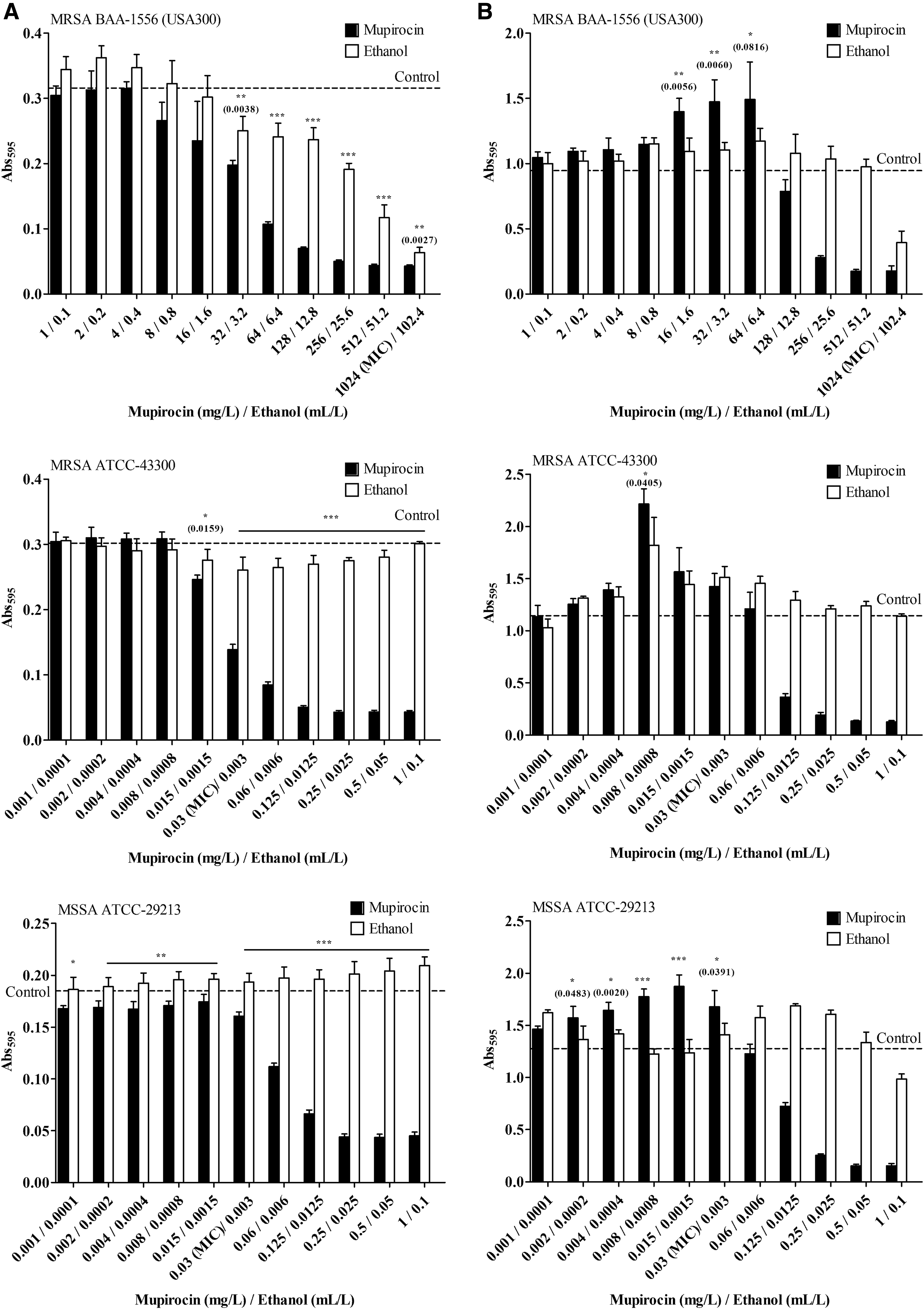
Planktonic cell growth
Low concentrations of ethanol treatment from 3.2 to 102.4 ml/L in the MRSA USA300 clone showed a weak antibacterial effect against planktonic cells in a concentration-dependent manner (Fig. 1A). However, no antibacterial effect of ethanol was found in mupirocin susceptible strains in either MRSA ATCC-43300 or MSSA ATCC-29213. However, ethanol treatment in the MSSA group seemed to promote the planktonic cells. The results indicated that ethanol had no antibiofilm effect against the tested S. aureus (Fig. 1B). In contrast, ethanol tended to promote S. aureus biofilm formation at a basal level higher than the control group.
Effect of low concentrations of mupirocin and ethanol on biofilm structure and cell viability
Figure 2 presents the biofilm structure and cell viability of MRSA BAA-1556 (USA300 clone), MRSA ATCC-43300, and MSSA ATCC-29213 exposed to the most significant doses for each strain (Fig. 1), which were 16, 0.008, and 0.015 mg/ml of mupirocin and 1.6, 0.0008, and 0.0015 ml/L of ethanol for 6 hr, respectively. Confocal laser scanning microscopy (CLSM) images showed that spontaneous mat-like biofilms were established with a higher cell density in the mupirocin treated group (Fig. 2D–F) compared to the control (Fig. 2A–C) and ethanol (Fig. 2G–I) groups, even though there were no differences in the biofilm thicknesses. However, the orthometric view of the biofilms showed viable (green) and nonviable (red) bacteria inside the biofilms. Each control group demonstrated green viable bacterial colonies with differences in the bacterial population densities. The greatest density was seen in the MRSA USA300 clone followed by MRSA ATCC-43300 and MSSA ATCC-29213. Once the cells were treated with low concentrations of mupirocin or ethanol, the condensed structures of the biofilms were analyzed. Greater live biomass in the biofilms was observed for each stacked layer compared to the control group, mainly in the MRSA USA300 clone (Fig. 3). The percentage of live biomass in MRSA USA300 clone biofilms remained around 48% in layer 20 after treatment with low concentrations of mupirocin or ethanol. However, in the control group the percentage of live biomass gradually decreased after layer 20. In addition, the ethanol treated group in MRSA ATCC-43300 gave the highest percentage of live biomass in the biofilms, while in MSSA ATCC-29213 the ethanol showed the lowest percentage of live biomass compared to the control and mupirocin treated groups.
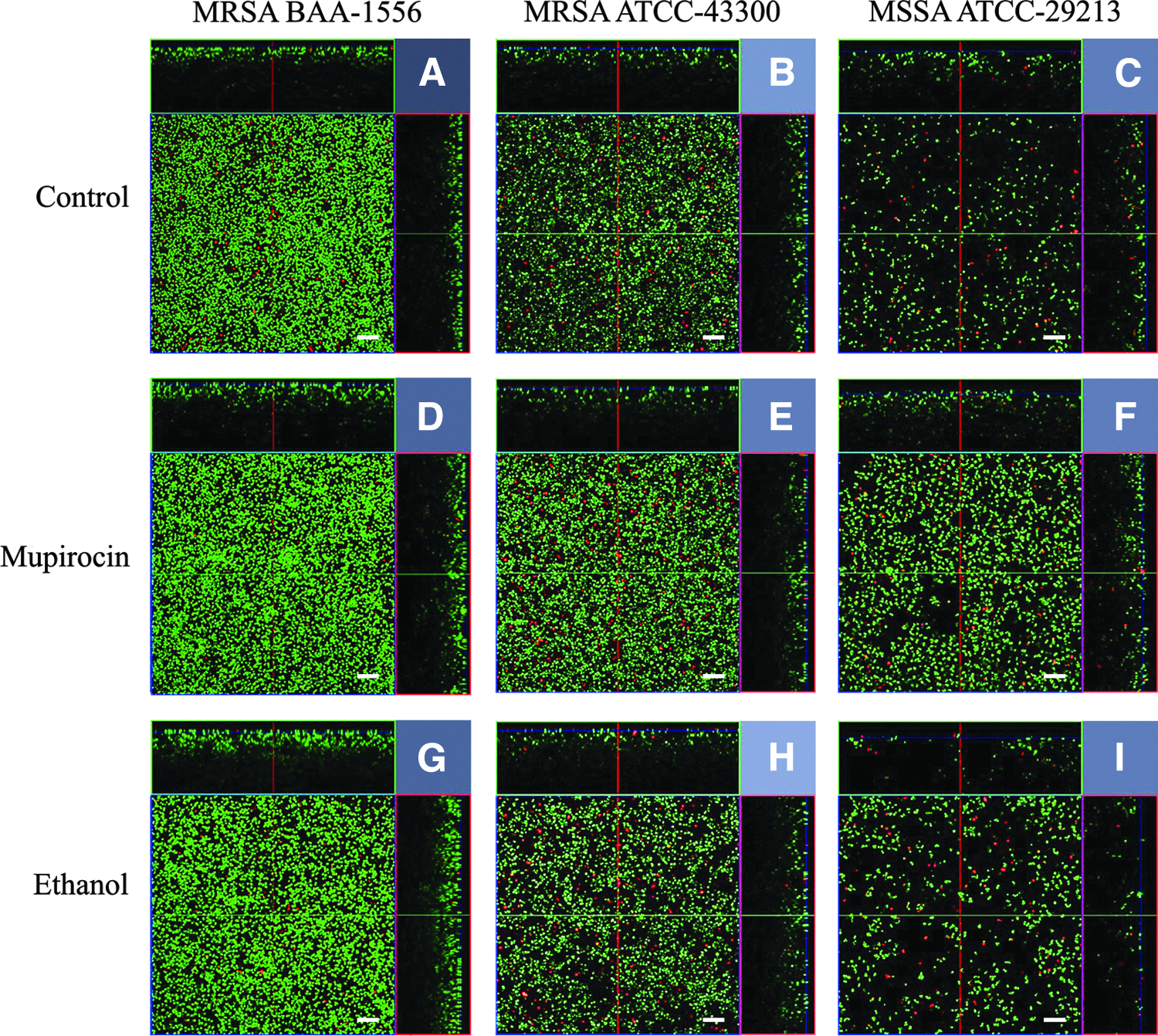
Biofilm structures and cell viability of MRSA ATCC BAA-1556 (USA300 clone), MRSA ATCC 43300, and MSSA ATCC 29213 exposed to the low concentrations of mupirocin and ethanol for 6 hr. LIVE/DEAD images were displayed as orthometric views. Each projection shows fields of 134.7 × 134.7 × 30 μm (xyz), as indicated. Scale bars = 10 μm.

Percentage live biomass in biofilms by layer after exposure to the low concentrations of mupirocin and ethanol for 6 hr analyzed by the COMSTAT2 software; control (-■-), mupirocin (-●-), and ethanol (-▲-) treated groups. MRSA BAA-1556 (USA300 clone) was treated with mupirocin 16 mg/L and ethanol 1.6 ml/L
Effect of low concentrations of antibiotics on biofilm-related gene expression of MRSA USA300 planktonic cells and biofilms
Quorum sensing (QS) related to S. aureus biofilm formation is encoded by the agr system. agrA and RNAIII are two intracellular effectors of the QS system. In addition, the sarA plays a role in the accumulation phase of staphylococcal biofilm formation. The expressions of agrA, RNAIII, and sarA involved in the biofilm formation of MRSA USA300 after exposure to 1/16x MIC of mupirocin, cefazolin, levofloxacin, gentamicin, and erythromycin for 6 hr are shown in Table 3. The results revealed that RNAIII expression in the sub-MIC mupirocin treated group for both planktonic cells and biofilms was the dominant gene showing the highest upregulation at 3 hr of incubation for up to 20 times compared to the control group. This was followed by a slight downregulation until 6 hr. However, agrA and sarA were not significantly different. agrA slightly decreased and sarA slightly increased from the baseline level at 6 hr. The overall analysis of gene expression from planktonic cells of the other four antibiotics indicated that the genes were initially upregulated at 3 hr, and the maximum upregulation was 3.61-fold found in agrA gene expression of cefazolin after 3 hr. The smallest increase in gene expression was 1.01-fold found in agrA gene expression of cefazolin after 5 hr. However, the gene expression of the biofilm showed various fluctuations with an initial low expression level and at the prolonged incubation time.
The significant differences in gene expression from the baseline level (p < 0.01).
N/A, not applicable.
Discussion
Ethanol is usually used as a solvent to dissolve various antibiotics in medical microbiology for in vitro testing. However, the final concentration of ethanol in the culture media depends on the concentration of the antibiotics selected. In this study, 1,024 mg/L of mupirocin contained 102.4 ml/L or 10.24% v/v of ethanol. Ethanol concentrations of 70–85% are commonly used as disinfectants. Ethanol disrupts the membrane structure, 23 interferes with cell division, 24 inhibits nutrient transport, and alters the pH and potential of the membrane.25,26 However, the present study confirmed that low concentrations of ethanol had weak antibacterial effects against S. aureus. Moreover, ethanol increased the production of planktonic cells and biofilms of S. aureus possibly due to bacteria within the biofilm that survived and had raised the metabolic activity, transcription of biofilm-promoting genes, and several antibiotic resistance genes, which resulted in the high production of biofilms. 18
S. aureus is considered to be susceptible to mupirocin at concentrations ≤4 mg/L that was shown in MRSA ATCC-43300 and MSSA ATCC-29213. MICs from 8 to 64 mg/L refer to low-level resistance, which is mediated by a mutation in the native isoleucyl-tRNA synthetase gene or ileS. 27 Bacteria with MICs of 128–256 mg/L are uncommon and also considered to have low-level resistance. The susceptibility testing showed a mupirocin resistance gene in the MRSA USA300 clone. A high level of mupirocin resistance (MIC ≥512 mg/L) is mediated by the expression of pUSA03, which is a conjugative plasmid containing ermC that encodes constitutive resistance to macrolides, lincosamides, and streptogramin B, and the mupA gene (ileS2) encodes an alternated isoleucyl-tRNA synthetase causing bacterial resistance to mupirocin. 28 However, a novel mupB gene was discovered and found that this gene also caused a high level of mupirocin resistance (MIC ≥1,024 mg/L) even if mupA is negative by PCR assay. 29
Crystal violet binding assay is commonly used to quantify biofilm formation. Studies reported that antibacterial activity of antibiotics occurred at high concentrations, whereas low concentrations produced a favorable environment for susceptible bacteria which eventually formed an antibiotic-induced biofilm.14,15 An incubation time of 6 hr represented the adhesion and formation of an early biofilm. This state showed the highest mupirocin-induced biofilm formation in comparison with 24 hr but no significant differences between the mupirocin treated group and positive control (data not shown). However, the antibiofilm effect of mupirocin was observed at the higher concentrations near or above MIC. This observation confirms that the correct use of mupirocin does not pose a threat because the effective concentrations were the same for planktonic cells.
The CLSM technique explained the metabolic activity and provided visualization of the three-dimensional development of bacteria present in biofilms. Many studies have shown the antibacterial effects against the biofilms by CLSM analysis using the LIVE/DEAD BacLight™ Bacterial Viability Kit.30–32 The present study showed an effective stain to differentiate viable and nonviable cells. The red fluorescing propidium iodide permeates into the bacteria with damaged cytoplasmic membranes, whereas SYTO9 can enter all cells and shows a green fluorescent in the total nucleic acid stain. 33 In addition, there are several methods to determine the metabolic activity of biofilms. They include the reduction of tetrazolium or resazurin salts to assess the activity of reductases from the respiratory chain, the use of green fluorescent protein labeled microorganisms, microautoradiography, and measuring the heat production rate by isothermal microcalorimetry (IMC). IMC is a label-free technique that is directly linked to the overall metabolism, including substrate consumption and by-product release. 34 However, COMSTAT2 software was used in this study to quantify the live biomass of the biofilms in each layer by taking image stacks of the biofilm created by CLSM as the source of data. The results indicated that the effect of low concentrations of mupirocin may vary in different strains of S. aureus. The greatest impact was on the increased density, particularly in the MRSA USA300 clone biofilms.
The difference in the number of viable cells and cell density can be described by the expression of biofilm formation and QS related genes. Gram-positive bacteria use oligopeptides as autoinducers, while Gram-negative bacteria use acylated homoserine lactones to communicate. 35 The QS system can be found in the interspecies of bacteria through autoinducers, which might be some of the signal molecules like acylated homoserine lactones, oligopeptides, or even some antibiotics.36,37 Bacteria use QS to regulate gene expression and physiological activities such as virulence, antibiotic production, or biofilm formation. QS in S. aureus is encoded by the agr (accessory gene regulator) system, which consists of two divergent transcription units leading to RNAII and RNAIII. agrA regulates metabolic genes, while RNAIII controls both biofilm formation and accumulation.38,39 Delta-toxin is a virulence factor encoded by the hld gene within RNAIII regulatory molecule that is responsive to the agr-encoded QS system. It was found that S. aureus that failed to produce delta-toxin had a greater tendency to form a biofilm because delta-toxin acts as a surfactant to limit the ability of bacterial aggregation into biofilms.40,41 Studies reported that the role of agr expression in biofilm development and behavior depended on environmental conditions and that the agr effect included both up- and downregulation of biofilms even for isolates within the same lineage.39,42 Recent evidence indicated that sarA (staphylococcal accessory regulator) is also a central regulatory element that controls the production of S. aureus virulence factors. sarA also plays a role in the synthesis of polysaccharide intercellular adhesion consisting of polymeric N-acetyl-glucosamine (PNAG), which is the primary determinant of the accumulation phase of staphylococcal biofilm formation regulated by the icaADBC operon-encoded enzyme.43,44 Reports indicated that sarA might promote biofilm formation by suppressing transcription of a repressor of PNAG synthesis, and the mutation of sarA results in a reduced capacity to produce PNAG and form a biofilm.44–46 The sarA mutant adhered better than the agr mutant or the wild-type strain suggesting that mutation of sarA might enhance at least the initial stages of biofilm formation. 47 Moreover, sarA mutants had a reduced capacity to adhere to immobilized fibronectin and other extracellular matrix proteins due to the increased production of proteolytic enzymes resulting in degradation of the fibronectin-binding protein.48,49 In this study, the results from real-time PCR illustrated the maximum regulation in gene expression at the beginning, especially RNAIII in the presence of subinhibitory concentrations of mupirocin in both planktonic cells and biofilms. This was followed by downregulation resulting in bacteria that had a greater potential to proliferate and form a biofilm as a function of cell density, which conformed to the CLSM images and other reports. 50 Low concentrations of the other four antibiotics (i.e., cefazolin, levofloxacin, gentamicin, and erythromycin) were selected for comparison with mupirocin regarding the gene expression of MRSA USA300 clone. Relative to the mupirocin treated group, a minor increase in gene expression of the planktonic cells occurred after treatment with these four antibiotics during the early incubation period. Gene expression then decreased with the same pattern as mupirocin, while gene expression of the biofilms varied with low expression levels.
In conclusion, the present study demonstrates the increased biofilm formation of S. aureus induced by subinhibitory concentrations of mupirocin. The findings confirmed by CLSM showed that the biofilms were formed with a high cell density, and the maximum upregulated gene of RNAIII was considerable to support the formation. Further studies using clinical strains presenting different mupirocin susceptibility profiles are required for clarification of the key mechanisms involved in the biofilm expression in S. aureus.
Footnotes
Acknowledgments
The authors thank all members and laboratory facilities of the Department of Microbiology and Infectious Diseases, Toho University School of Medicine, Tokyo, Japan. This work was supported by the Thailand Research Fund through the Royal Golden Jubilee PhD Program (PHD/0062/2557 to R.S.).
Disclosure Statement
No competing financial interests exist.