
Abstract
Colistin is considered a last-hope antibiotic against extensively drug-resistant isolates of Acinetobacter baumannii. Resistance to colistin has been rarely reported for A. baumannii. Genetic alterations in the PmrA-PmrB two-component system and lipid A biosynthesis genes may be associated with colistin resistance. We investigated molecular mechanisms of colistin resistance in three clinical colistin-resistant (ColR) and two colistin-susceptible (ColS) A. baumannii isolates. A ColR mutant was generated in vitro by repetitive drug exposure. The pmrA, pmrB, lpxA, lpxC, and lpxD genes were amplified and sequenced. To evaluate association between colistin resistance and upregulation of pmrCAB operon, transcriptional level of the pmrC gene encoding for lipid A phosphoethanolamine (PEtN) transferase was quantified by reverse transcription quantitative PCR (RT-qPCR) analysis. All clinical and in vitro-selected ColR isolates harbored at least one point mutation in the pmrB gene, including A142V, P233S, T235I, and A227V substitutions as well as duplication of H325. No alteration was found in the pmrA and other amino acid substitutions identified in the pmrB as well as lpx genes did not seem to be involved in colistin resistance as they were found in both ColS and ColR isolates. RT-qPCR analysis revealed a correlation between colistin resistance and pmrC overexpression. Specific alterations in the PmrB, linked to overproduction of PEtN transferase, triggered colistin resistance in the studied A. baumannii isolates.
Introduction
A
Materials and Methods
Bacterial strains and antimicrobial susceptibility testing
Three clinical colistin-resistant (ColR) and two colistin-susceptible (ColS) A. baumannii strains isolated from separate patients hospitalized in Imam Khomeini hospital, the largest medical center of Iran with 1400 beds, were investigated. Isolates of A. baumannii were identified by microscopic examination, conventional biochemical methods, sequencing of the 16S-23S rRNA gene intergenic spacer (ITS) region using universal primers 1512F (5-GTCGTAACAAGGTAGCCGTA-3) and 6R (5-GGGTTCCCCCATTCGGAAAT-3), 15 and detection of blaOXA-51-like carbapenemase gene by PCR. 16 Drug susceptibility testing for colistin as well as other antimicrobials was performed by using the disk diffusion method. Minimal inhibitory concentrations (MICs) of colistin were determined by the broth macrodilution method.17,18 Isolates with MICs of colistin >2 mg/L were categorized as resistant according to guidelines described by the European Committee on Antimicrobial Susceptibility Testing (EUCAST) (www.eucast.org). Pseudomonas aeruginosa ATCC27853 was used as quality control for antimicrobial susceptibility testing.
Molecular characterization of colistin resistance
To investigate the leading causes of colistin resistance, genomic DNA was extracted from the bacterial strains, and PCR amplification of the entire region of the pmrB, pmrA (encoding for PmrB sensor kinase and PmrA response regulator, respectively), lpxA, lpxC, and lpxD (coding for essential enzymes involved in lipid A biosynthesis) genes was carried out using primers listed in Table 1. PCR products were analyzed by gel electrophoresis and were sent to SEQLAB Sequence Laboratories (Göttingen GmbH, Germany) for sequencing. The obtained gene sequencing results were compared to those of A. baumannii ATCC 17978.
RT-qPCR, reverse transcription quantitative PCR.
Laboratory-induced development of resistance to colistin
To assess whether colistin nonsusceptibility in A. baumannii occurs after colistin exposure, we performed an in vitro exposure experiment using a method described previously, with minor modifications. 19 To in vitro induction of colistin resistance we subcultured clinical ColS A. baumannii isolate (AB424S), which had a low MIC value in media containing increasing concentration gradients of colistin sulfate (Product Number: C4461; Sigma-Aldrich). Fresh Mueller–Hinton agar media supplemented with a colistin concentration of half the MIC values (0.25 μg/ml) and higher concentrations (0.5, 1, 2, 4, 8, 16 μg/ml) were inoculated with a 3 × 105 cfu/ml bacterial suspension and incubated for 48–72 hr. Mutants were selected in a stepwise manner by picking colonies growing at the highest antibiotic concentration at each stage and again subculturing on media with a higher colistin concentration. Seventeen selection cycles were required to obtain one resistant mutant isolate. Colistin MIC in the selected resistant isolate was determined by the broth macrodilution method.
Transcriptional analysis of pmrC by reverse transcription-quantitative PCR analysis
To investigate whether colistin resistance is associated with pmrCAB upregulation, the expression level of the pmrC gene (which encodes the protein that adds PEtN to lipid A) was evaluated. Bacterial cells were grown aerobically in Luria-Bertani (LB) broth until midlog phase. Bacterial cell wall was broken by several freeze/thaw cycles and total RNA was extracted using a GeneAll RiboEx total RNA extraction kit (GeneAll Biotechnology, Seoul, Korea). Reverse transcription (RT) was performed using the RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific) with 1 μg of DNase I (Takara Biotechnology, Dalian, China)-treated RNA. The real-time RT-PCR amplification was performed on an Eco Real-Time PCR system (Illumina) using Power SYBR green PCR master mix (Applied Biosystems). Primers ((Table 1) (Metabion, Martinsried, Germany)) were added to the appropriate master mix at concentrations of 200 nM (pmrC) or 140 nM (16S rRNA) and 2 μl of 1:15 diluted cDNA product was used as a template in 12.5 μl reaction volume. Briefly, real-time RT-PCR was performed under the following conditions: 1 cycle of 95°C for 10 min, 45 cycles of 95°C for 15 sec, 56°C for 30 sec, and 72°C for 30 sec, followed by final extension at 72°C for 2 min. The relative gene expression levels were calculated using the 2−ΔΔCT formula with 16S rRNA housekeeping gene as internal control. 20
Nucleotide sequence accession numbers
The nucleotide sequences of the pmrA, pmrB, lpxA, lpxC, and lpxD genes as well as 16S-23S rRNA gene ITS region from ColR and ColS A. baumannii isolates obtained in this study have been deposited at GenBank nucleotide sequence databases under the accession numbers MF673412 to MF673424, MG870548 to MG870560, and MG870543 to MG870547.
Results
Characteristics of the laboratory-induced and clinical ColR isolates
The three clinical ColR A. baumannii isolates were obtained from separate patients with respiratory infection hospitalized in the Intensive Care Unit of Imam Khomeini hospital. Based on ITS fragment analysis, all the studied isolates belonged to A. baumannii species and blaOXA-51-like carbapenemase gene were detected by PCR in all isolates. All three ColR isolates were found to be resistant to all tested antimicrobials, including tigecycline and colistin (shown in Table 2), and patients expired during their stay at the hospital due to the severity of the infection. A ColR mutant AB424R was generated in vitro by iterative subculturing of isolate AB424S, which was resistant to all antimicrobials except for colistin. We chose an XDR isolate for colistin-resistant induction experiment to unravel mechanisms mediating colistin resistance when the bacterium was resistant to other antimicrobials. MIC of the in vitro-selected ColR isolate was found to be 8 mg/L compared with 0.5 mg/L, which was found for progenitor-susceptible isolate AB424S. No significant changes were observed in susceptibility to any other tested antimicrobials listed in Table 2.
MIC, minimal inhibitory concentration; GM, gentamicin; TN, tobramycin; AK, amikacin; NET, netilmicin; C, chloramphenicol; IMI, imipenem, MEM, meropenem; DOR, doripenem; CIP, ciprofloxacin; OFX, ofloxacin; TM, trimethoprim; PTZ, piperacillin/tazobactam; TC, ticarcillin; TIM, ticarcillin/clavulanic acid; CRO, ceftriaxone; CTX, cefotaxime; CAZ, ceftazidime; CPM, cefepime; AP, ampicillin; SAM, ampicillin/sulbactam, ATM, aztreonam; CT, colistin; PB, polymyxin; T, tetracycline; DXT, doxycycline; MN, minocycline; TGC, tigecycline.
All isolates harbored two SNPs (A138T and A444V).
Specific PmrB alterations are the leading causes of acquired colistin resistance
The entire region of the pmrA, pmrB, lpxA, lpxC, and lpxD genes of the three clinical, one in vitro-selected ColR, and two clinical ColS A. baumannii strains, was amplified by PCR. Two substitutions were identified in the pmrB gene in each of the laboratory-induced and clinical ColR isolates AB8, AB40, AB67, AB424R, and ColS AB424S, including A138T (GCT> ACT) and A444V (GCG>GTG), which were absent from ColS AB429 and A. baumannii ATCC 17978. All 4 ColR isolates revealed at least one point mutation in the pmrB gene. Strain AB8 (MIC >512 mg/L) harbored two point mutations, including A142V (GCG>GTG) and P233S (CCT>TCT). Strain AB40 with colistin MIC values of 256 mg/L contained A142V (GCG >GTG) substitution. Also, this isolate was characterized with another genetic alteration in which codon 325 (CAT, standing for histidine) was duplicated, resulting in production of protein with 445 amino acids instead of 444 amino acids. Both AB67 and AB424R with MIC values of 8 mg/L were found to have T235I (ACT> ATT) and A227V (GCT>GTT) substitutions in the pmrB gene, respectively (Table 2). Compared to sequences of A. baumannii ATCC 17978, all ColS and ColR isolates revealed Y131H substitution in the lpxA, and C120R + N287D substitution in the lpxC. Also, two substitutions were found in the lpxD gene, including V63I (in ColS AB429) and E117K (in ColS AB424 and all ColR isolates). No differences were found in the pmrA gene of any of the ColR isolates compared with ColS isolates and A. baumannii ATCC 17978.
Colistin resistance is associated with pmrC overexpression and decoration of LPS with PEtN
To evaluate the association between the colistin-resistant phenotype and overexpression of the pmrCAB operon, the transcription levels of the pmrC gene were analyzed by RT quantitative PCR (RT-qPCR) method. In general, laboratory-induced and all clinical ColR isolates showed an elevation in the transcription level of the pmrC from 3- to 254-fold compared with the levels obtained for our ColS wild-type isolates. Isolates AB40 and AB8 containing several genetic alterations in different loci of the pmrB gene were characterized with an extremely high level of pmrC expression (254-fold and 233-fold, respectively). Approximately 7.4- and 3-fold increases in the expression of pmrC gene were observed for AB67 and AB424R respectively (Fig. 1).
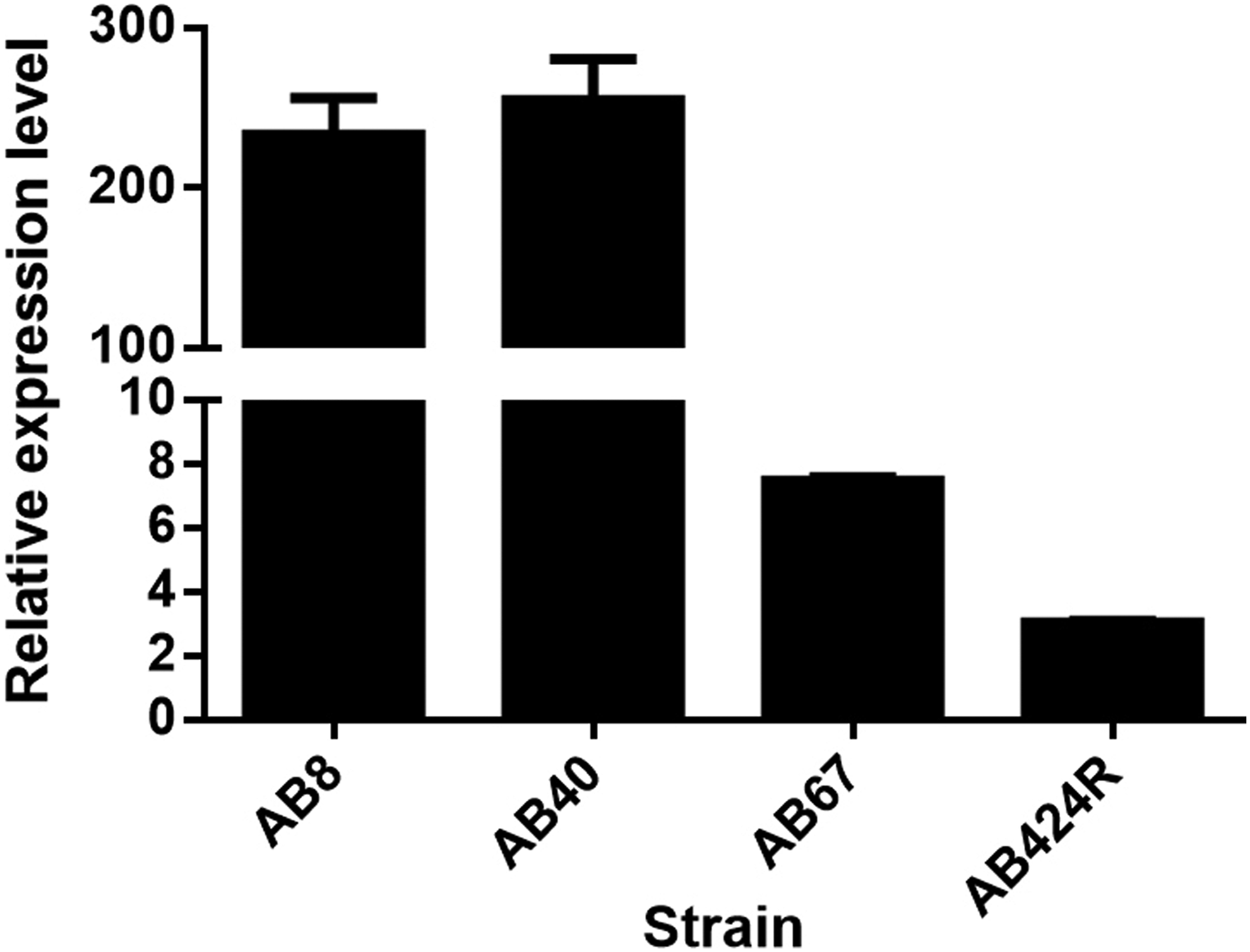
Transcriptional analysis of pmrC gene in laboratory-induced and clinical colistin-resistant isolates compared with that of colistin-susceptible strain.
Discussion
While colistin and tigecycline are often considered as the only available antimicrobials against XDR A. baumannii infections, some isolates have developed resistance to these last-line therapeutic options. 3 Here, we report the emergence of pandrug-resistant A. baumannii isolates from Imam Khomeini Hospital the largest hospital of the country. All three ColR clinical A. baumannii isolates were obtained from separate patients and represented a high case-fatality rate (100%). The laboratory-induced ColR isolate AB424R had a 16-fold change in colistin MIC, obtained by 17 selection cycles (45 days), indicating that long-time drug exposure is involved in the development of resistance. We found that genetic alterations in the pmrB facilitated resistance to colistin. The genetic alterations found within the pmrB gene were A142V, P233S, T235I, A227V, and H325 duplication. According to domain prediction results provided by InterPro a and NCBI databases, b P233S,T235I, and A227V substitutions were found to be located at an essential position for dimerization and phosphoacceptor domains (positions 218–280), which includes the site of histidine autophosphorylation and phosphate transfer reactions. During the activation of PmrB sensor kinase, the phosphoryl group is transferred to the Asp acceptor residue of response regulator PmrA protein. These substitutions have been found in ColR clinical strains or laboratory mutants in previous studies12,21,22 and the positions seem critical for PmrB activity. In isolate AB40, codon 325 (histidine), which was duplicated, lies in a position next to ATPase domain (positions 326–437) of the histidine kinase PmrB protein and may exert some effects on phosphorylation of PmrB. Moreover, isolates AB8 and AB40 revealed a novel A142V substitution in the PmrB, which was found to have a deleterious effect on protein structure as determined by the PROVEAN prediction tool. 23 However, the exact role of this novel alteration in resistance conferring requires to be further studied by confirmatory tests. Altogether, these alterations most likely result in some activation of PmrB in the absence of the physiological activating signals and the subsequent interaction with PmrA response regulator. Upon activation, the phosphorylated PmrA binds to the promoter region of the pmrCAB operon, and results in the upregulation of genes in the operon, including pmrC. To examine the accuracy of this hypothesis, the transcription level of the pmrC gene was evaluated using the RT-qPCR method. We observed a significant correlation between colistin resistance and overexpression of pmrC. All ColR isolates revealed overexpressed pmrC with AB40 and AB8 showing the highest expression levels. Also, we found that expression of pmrC gene was irrespective of the absence or presence of colistin during the culturing of isolates for RNA extraction (data not shown). Since A138T and A444V substitutions were found in ColS isolate AB424S before resistance induction and have been reported previously in several ColS isolates,21,22,24 they do not seem to be involved in colistin resistance in A. baumannii and most likely do not have an essential impact on PmrB activity. It is hypothesized that only specific PmrB alterations, which happen in more conserved domains, cause PmrB activation and subsequently pmrCAB upregulation and colistin resistance. Therefore, evaluation of pmrC expression level can provide more accurate data about genotypic susceptibility of A. baumannii to colistin compared to sequencing of genes coding for PmrA-PmrB TCRS. Beceiro et al. 22 did not detect any genetic alterations in the pmrA and pmrC genes in ColR A. baumannii isolates and they found several point mutations in the pmrB gene (M145K, P233S, L87F, S14L, F387Y, S403F, A227V, and N353Y), which were associated with overexpression of pmrCAB operon. Park et al. 25 identified six kinds of amino acid alterations in the pmrB (T192I, I121F, A184V, Q228P, A183T, and P190S) in some of the in vitro-selected ColR mutants with no mutations in pmrA and pmrC genes. They found that pmrB mutations linked with pmrA and pmrB overexpression. Similar to a study performed by Oikonomou et al., 24 sequence analysis of the lpxA, lpxC, and lpxD genes revealed some amino acid substitutions that did not seem to be related to colistin resistance as they were detected in ColS isolates as well. It has been demonstrated that LPS-deficient A. baumannii isolates harboring deleterious genetic alterations in lipid A biosynthesis genes remain susceptible to other antimicrobials due to altered permeability of the outer membrane, which commonly works as a main barrier against penetration of many antimicrobials. 13 Moffat et al. demonstrated that an lpxA mutant strain, highly resistant to colistin, displayed a significant increase in sensitivity to the other clinically relevant antibiotics, including a Gram-positive antibacterial teicoplanin. 13 Since the isolates studied in the current work were resistant to all tested antimicrobials, colistin resistance due to loss of LPS was unlikely.
In summary, we report here the isolation of pandrug-resistant isolates of A. baumannii with high case-fatality rate. We found that colistin resistance in ColR isolates obtained from Iranian patients was mediated by PmrA-PmrB TCRS but not loss of LPS. A laboratory-derived mutant characterized with mutated PmrB was obtained during the repetitive drug exposure over a period of 17 cycles, indicating that long-term colistin exposure could be an important risk factor for the development of resistance. Not all pmrB alterations can lead to colistin resistance as A138T and A444V substitutions were found in a ColS isolate with a low colistin MIC value and pmrC expression level. Colistin-resistant phenotype linked with pmrC overexpression indicating that overproduction of LPS-modifying enzyme PEtN transferase, and therefore LPS modification with positively charged PEtN moiety, is involved in colistin resistance conferring in the studied A. baumannii isolates from Iran.
Footnotes
Acknowledgment
This work was supported by the University of Tabriz.
Ethical Approval and Consent to Participate
The research has been approved by the Ethics Committee of Tehran University of Medical Sciences under the reference number R.TUMS.REC.1394.2166.
Disclosure Statement
No competing financial interests exist.