
Abstract
Aims:
Cross-contamination of feed with antibiotics causes pigs to become unintentionally exposed to low concentrations of antibiotics. This study investigates the effect of residues of doxycycline hyclate (DOX) in an ex vivo model of the intestinal tract of pigs, focusing on the microbial community, microbial activity, and the enrichment of resistant bacteria and resistance genes.
Results:
The effect of three concentrations DOX were tested; 1 and 4 mg/L correspond to the intestinal concentrations when pigs are fed a compound feed containing 3% of a therapeutic dose, and a reference concentration of 16 mg/L. These were continuously administered to a chemostat, simulating the microbial ecosystem of the pig cecum and inoculated with cecal content of organically grown pigs. The administration of even the lowest DOX concentration caused a significant decrease in bacterial activity, while the microbial community profile appeared to remain unaffected by any of the concentrations. A concentration of 1 mg/L DOX caused minor selection pressure for tetracycline-resistant Escherichia coli but no other groups enumerated with plate cultivation, while 4 mg/L induced major enrichment of tetracycline-resistant E. coli, Enterobacteriaceae and total anaerobes. High abundances of tet(Q), tet(M), tet(W), tet(O), and tet(B) were detected in the inoculum and also before antibiotic administration in the chemostat and did not significantly increase during administration of 1 and 4 mg/L DOX. Only 16 mg/L DOX caused minor enrichments.
Conclusions:
Cross-contamination concentrations of doxycycline, as a result of cross-contamination, cause a selection pressure for resistant bacteria and negatively affect microbial activity.
Introduction
I
Residual amounts of antimicrobial compounds in the feed will pass the intestinal tract before reaching systemic circulation. Concentrations in the gut compartments depend on the initial concentration in the feed, and the pharmacokinetic properties and bioavailability of the drug. In an in vivo experiment, Peeters et al. studied the intestinal concentrations of chlortetracycline, doxycycline hyclate (DOX), and sulfadiazine-trimethoprim when pigs were fed a compound feed including 3% of the maximum recommended dose (MRD) of the antibiotic, simulating a cross-contamination event. Doxycycline was detected with concentrations in the range of 1 and 4 mg/kg intestinal content. 10 In this study, we aimed to investigate the effect of these concentrations of DOX on the microbial community, microbial activity, and the resistance development in an ex vivo model of the intestinal ecosystem of pigs. Therefore, a chemostat was designed to simulate the microbial ecosystem of the pig cecum, a prominent organ of the pig's gastrointestinal tract characterized by a dense microbial ecosystem for the fermentation of soluble fibers.11,12 While current in vivo studies are restricted by fecal samples or end point sampling after slaughter, the ex vivo model provides the opportunity of easy and standardized sampling to study the effect of DOX on the simulated microbial community of the cecum, without interference of host effects. Although many types of antibiotics are used in the farming industry, we specifically focused on DOX as it remains one of the most frequently used antibiotics in pig husbandry 7 and remains persistent and active in different matrices over extended periods, evidenced by a half-life of 120 days in liquid pig manure 13 and a shelf-life of 3–5 months as part of a medicated feed. 14
Materials and Methods
Ex vivo simulation of the pig cecum
The physical and chemical characteristics of the pig cecum were simulated with an ex vivo continuous fermentation model, as previously described,15,16 with minor modifications. The equipment consisted of similar BioFlo 110 and BioFlo 115 bioreactors, with a 1.3 liter fermenter vessel (New Brunswick Scientific, Enfield, CT). Both units were set up in parallel to perform two experimental runs simultaneously. The reactor vessels were filled with 0.5 liter feed medium, autoclaved, and brought to a pH of 6.5 and a temperature of 37°C. After inoculation, the reactors were initially operated in batch mode for 24 hr, followed by 9 consecutive days of continuous culture (Fig. 1). Fresh medium was added via a peristaltic pump at a rate of 1.8 mL/min and reactor content was removed as waste at the same rate to retain a working volume of 0.5 liter and a transit time of 4.6 hr, reflecting the nominal residence time of cecal content in pigs. The pH was kept constant at 6.5 with 3 mol/L NaOH solution and the temperature was maintained at 37°C. The temperature, pH, and redox were continuously monitored with sensors (Ingold® probes; Mettler Toledo, Zaventem, Belgium). Anaerobic conditions were maintained by flushing the headspace of the vessels with an anaerobic gas mixture (20% CO2, 80% N2) at a flow rate of 20 mL/hr. The reactor content was kept homogeneous through constant agitation (150 rpm). After a 24 hr batch incubation period, continuous fermentation with regular feed medium was carried out during 4 days ( = pretreatment blanc control period). From day 5 onward, the regular feed medium was complemented with 1, 4, or 16 mg/L DOX (Fagron, Waregem, Belgium). The concentrations of 1 and 4 mg/L represent the upper and lower limits of intestinal concentrations in pigs exposed to feed containing 3% of the MRD of DOX. 10 In addition, we tested 16 mg/L DOX as this concentration is above the epidemiological cutoff values (ECOFF) of doxycycline for all species listed in the EUCAST database. 17 This concentration is therefore considered a positive control to investigate a dose that would certainly elicit a response of the wild-type and susceptible species (and some of the resistant species) in the bacterial communities of the bioreactors. This positive control was compared to the two low concentrations, resulting from a cross-contamination event.
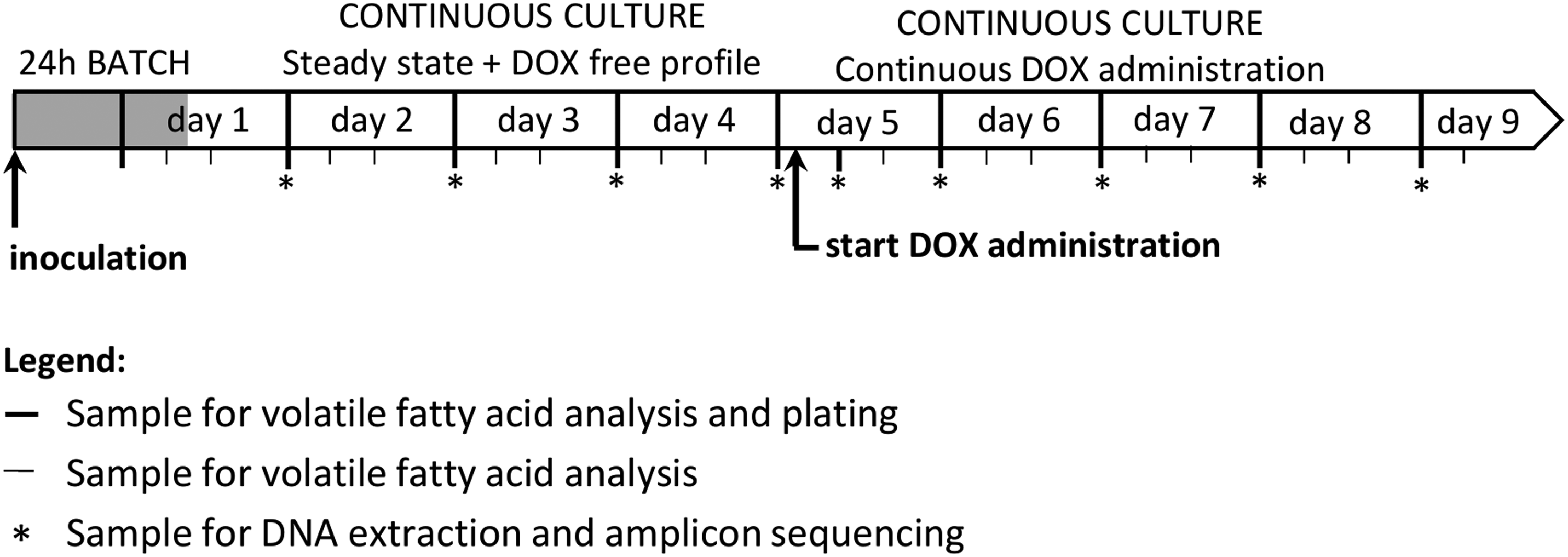
Chronology of a reactor run. Samples for plating and DNA extraction were collected at 9:00 every morning. On day 0, the reactor was inoculated with a pooled cecal inoculum and operated as a batch culture during 24 hr. During day 1, the feed medium was prepared and the reactor changes to continuous culture. After sample collection on day 5, feed medium was supplemented with 1, 4, or 16 mg/L DOX. Six hours after starting continuous administration of doxycycline, an extra sample was collected for plating and DNA extraction. DOX, doxycycline hyclate.
Three replicate reactor runs were executed for each tested concentration of DOX. After a 4 day pretreatment period with regular feed medium, considered as the blanc control period, reactor runs 1, 2, and 3 received 1 mg/L DOX, reactor runs 4, 5, and 6 received 4 mg/L DOX and reactor runs 7, 8, and 9 received 16 mg/L DOX. Ten milliliters of the reactor content was sampled daily at 9 AM for microbial and molecular analysis. Two additional samples were collected in the afternoon (1 PM, 5 PM) for accurate monitoring of the volatile fatty acid composition. On day 5, samples for microbial and molecular analysis were collected just before starting continuous antibiotic administration, and 6 hr thereafter (Fig. 1). In addition, as an extra blanc control, one run (run 10) was conducted without the addition of DOX. After inoculation and the 24 hr batch period as described above, samples were taken daily at 9 AM only for microbial analysis.
Chemicals, growth medium, and inoculum
The feed medium had a complex composition, mimicking the nutritional availability in the pig cecum. The chemicals were obtained from Sigma (Bornem, Belgium), unless stated otherwise. The feed medium consisted of (in g/L) starch from corn (5), casein from bovine milk (10), casein hydrolysate acid (0.5), soybean oil (1) (AD Delhaize, Belgium), anhydrous L-cystein hydrochloride (0.65), pectin from citrus peel (2.7), alphacel (13.8) (MP Biochemicals, Brussels, Belgium), mucin from porcine stomach type II (5), vitamin-mineral premix for pigs (2.35) (Vitamex, Drongen, Belgium), KHPO4 (0.93) (Merck, Overijse, Belgium), and Na2HPO4.12H2O (1.12). The medium was acidified with 4 mL of 37% HCl to pH 2 and stored at 5°C in autoclaved 13 liter pyrex bottles under constant magnetic agitation. Beforehand, a zone of inhibition test confirmed that prolonged incubation at a pH of 2 had no negative influence on the activity of doxycycline (data not shown).
At the onset of each new experimental run, the reactors were inoculated with an identical and homogenous inoculum of cecal bacteria. To obtain a representative inoculum, 10 organically grown pigs were selected from a farm that did not use antimicrobial therapy during growth. The cecal microbiota of these pigs as inoculum allowed to investigate the effect of antibiotics on an ex vivo microbial community that has not yet been into deliberate contact with antimicrobial compounds. At slaughter age, the ceca of these pigs were removed during the slaughter process, tied up, and individually stored in stomacher bags on ice for transport. In the lab, the cecal contents were poured out in a measuring cup and thoroughly stirred. The cecal content of 10 pigs were pooled to obtain enough inoculum for all technical repetitions of the reactor runs and for qPCR analysis of tet genes. Subsequently, the pooled cecal contents were diluted with anaerobic phosphate buffer (8.8 g K2HPO4 + 6.8 g KH2PO4 + 1 g sodium thioglycolate in 1 liter dH2O) to a 1:1 ratio, homogenized for 2 min in a stomacher and centrifuged for 3 min at 500 g. The supernatant was supplemented with 15% w/v glycerol and subsamples of 12 mL were stored at −80°C. Before inoculation, a subsample was thawed in a 37°C water bath.
Selective cultivation
The total number of colony forming units (CFUs) and the number of tetracycline-resistant CFUs of cultivable anaerobes, Escherichia coli and Enterobacteriaceae in the reactor were enumerated on agar plates. In these bioassays, we defined “resistance” as the ability of bacteria to proliferate in the presence of 10.5 mg DOX per liter agar. A 1 mL aliquot of each sample was used to make a 1:10 serial dilution series in 9 mL Maximum Recovery Diluent (Oxoid, Aalst, Belgium). Hundred microliters of each dilution was plated on Reinforced Clostridial Agar (RCA) (Oxoid) for cultivation of Clostridia and other anaerobes, Violet Red Bile Glucose Agar (VRBGA) (Oxoid) for the cultivation of Enterobacteriaceae and RAPID'E.coli2 Agar for the cultivation of E. coli (Bio-Rad, Temse, Belgium) plates without DOX (total colony count) and with 10.5 mg/L filter-sterilized DOX (colony count of tetracycline-resistant bacteria). The plates were prepared according to the manufacturer's instructions. RCA plates were anaerobically incubated at 37°C for 3 days, VRBGA plates were incubated for 24 hr at 37°C and RAPID'E.coli2 plates at 44°C for 24 hr before counting the colonies. The samples taken from the blanc run (run 10) were only plated on VRBGA with and without DOX, this was to check the stability of a reactor run and the Enterobacteriaceae were chosen as indicator organisms.
Fatty acid quantification
C2-C8 (including isoforms C4-C6) fatty acid analysis was performed according to Andersen et al. 18 In short, liquid reactor samples were conditioned with sulfuric acid, sodium chloride, and 2-methyl hexanoic acid as internal standard. Short chain fatty acids (SCFA) and branched chain fatty acids (BCFA) were extracted with diethyl ether and quantified with gas chromatography.
DNA extraction
Nine reactor runs were executed and DNA was extracted from nine samples per run. The samples were collected on day 2, 3, 4, 5 (before and 6 hr after starting DOX administration), 6, 7, 8, and 9. DNA extractions were performed with the repeated bead beating and column protocol (RBB+C), as previously described 19 but with minor adaptations. Five-hundred microliters of a homogenous reactor sample was transferred to a cryovial with 0.4 g autoclaved zirconia beads (ø = 0.1 mm). Cells were ruptured in a FastPrep®-24 (MP Biomedicals) (two times 45 sec, 6 m s–1) in the presence of a lysis buffer, adopted from Vilchez-Vargas et al. 20 DNA quality and quantity was subsequently checked with Nanodrop™ (NanoDrop ND-1000; Thermo Scientific) and the Quantus™ double-stranded DNA assay (Promega, Madison, WI).
Quantitative PCR
qPCR analysis was performed on a LightCycler® 480 Real-time PCR system (Roche). Total 16S rRNA gene abundance, as a proxy for bacterial abundance, was quantified using SYBR Green technology. For each DNA extract, a 1,000-fold dilution was made and analyzed in duplicate. The qPCR was carried out with a reaction mixture and PCR conditions described in Desloover et al. 21 and eubacterial primers suggested in Ovreas et al. 22 (Table 1). The abundances of five tetracycline resistance genes tet(Q), tet(O), tet(M), tet(W), and tet(B) were quantified using TaqMan qPCR assays, using a 10 or 100-fold dilution of the DNA extracts. The primer/probe sets were reported in Knapp et al. 23 (Table 1). Each reaction mixture contained, in a total volume of 25 μL, 12.5 μL TaqMan® Environmental Master Mix 2.0 (Applied Biosystems, Foster City, CA), 300 nmol/L of each primer, 100 nmol/L probe, and 5 μL of template DNA. The PCR program was carried out in a thermal cycling process consisting of a hot start activation step of 10 min at 95°C, followed by 40 cycles of 15 sec at 95°C and 1 min at 60°C. Within each run, a standard curve was constructed using a 10-fold dilution series of plasmid DNA (IDT, Coralville, IA) to determine PCR efficiency (Table 1). The total number of gene copies was calculated by converting the quantification cycle values (Cq) to gene copy abundances, using the standard curve and taking the PCR efficiency into account.
The TaqMan probes were dual-labeled with 5′-FAM (fluorescein) and 3′-BHQ1 (Black Hole Quencher®).
qPCR efficiency of the SYBR assay was quantified with a dilution series of the commercial vector pIDTSMART_AMP (IDT, Coralville, IA), including the target sequence of the 16S gene fragment of Prevotella ruminicola (NCBI: NC_014033).
qPCR efficiency of the TaqMan assays were quantified with a dilution series of the commercial vector pIDTSMART_AMP (IDT), including the concatenated target sequences of each tested resistance gene, flanked by “TATA.” The target sequences were obtained from NCBI: JQ966986.1 for tet(B), KF408178.1 for tet(M), M18896.2 for tet(O), X58717.1 for tet(Q), AF202986.1for tet(W).
Amplicon sequencing and processing of sequenced reads
Amplicon sequencing of the eubacterial V3-V4 variable region of the 16S rRNA gene was done on 81 samples from nine reactor runs and on three replicate samples of the inoculum to determine the taxonomic profiles of the microbial communities. Library preparation for Illumina amplicon sequencing and processing of the sequenced reads was carried out as previously described. 24 The raw sequence reads are available on the NCBI Sequence Read Archive under the project accession number PRJNA351773. Instead of using the Usearch “fastq_mergepairs” command, the forward and reverse reads were merged using PEAR 0.9.8 with a minimum overlap length of 120 bp, a minimum and maximum resulting length of 400 bp and 450 bp and a quality threshold of 30 with a minimum length of 200 bp after trimming. 25 The final OTU table was normalized using CSS 26 to account for variable library sizes, using the QIIME command “normalize_table.py”. 27 Processing the data resulted in an average library size of 57 199 reads per sample. Rarefaction analysis was done to ascertain that the library size of each sample was sufficient to analyze the bacterial community (Supplementary Fig. S1). Simpson diversity and observed richness were calculated with the Phyloseq package in R. 28 The OTU table was used to calculate relative abundances and summarize the table to family level, thus obtaining a table with the relative abundances of each eubacterial family in samples collected from nine reactor runs and three replicate samples of the inoculum (i.e., cecal samples). The family level table was used to calculate pairwise Bray-Curtis dissimilarity indices and differences between communities were visualized with nonmetric multidimensional scaling (NMDS) using the Vegan package in R. 29
Statistical analysis
Significant changes in fatty acid concentrations, abundances of specific tetracycline resistance genes, total number of CFUs and number of resistant CFUs of E. coli, Enterobacteriaceae, and cultivable anaerobes as a response to DOX administration were analyzed using a linear mixed effects model 30 including “treatment” as fixed effect and “reactor run” as random effect, with Tukey adjustment for post hoc testing. The short-term effect of DOX treatment on the microbial activity was analyzed by comparing the SCFA and BCFA data from 2 days before DOX administration with data from samples during the first 2 days of DOX administration. Significant changes in the number of CFUs of resistant species (plate cultivation) and resistance gene abundance (qPCR assays) as result of DOX administration were tested in a similar way but using data from all sampling points. Significant results obtained with the linear mixed effects model were graphically verified: a pretreatment mean and a 95% confidence interval (CI95) were calculated for each parameter. Only parameter values that surpassed the CI95 boundaries during DOX administration were accepted as a true significant response to DOX administration.
Results
Data analysis of samples from reactor run 2 (1 mg/L DOX) revealed unexpected problems at the startup phase of this run. During the first 2 days of chemostat, the microbial community had a 10-fold lower concentration of main and BCFA and metabarcoding revealed no Bacteroidetes. Concomitantly, the abundance of tet(Q) gene copies, mainly associated with Prevotella and Bacteroides,31,32 were much lower during the first 2 days compared with other reactor runs. Because reactor run two deviated from all other runs, it was not considered representative and thus excluded from further analysis.
Microbial activity
Main SCFA and BCFA concentrations serve as markers for bacterial metabolic activity and were used to assess steady-state after reactor startup and stabilization. Upon administration of 1, 4, and 16 mg/DOX, average BCFA concentrations significantly decreased with 43% (p < 0.001), 36% (p < 0.001), and 28% (p < 0.001) in isovalerate concentration and 17% (p < 0.001), 20% (p < 0.001), and 14% (p < 0.01) in isobutyrate concentration, respectively (Fig. 2). DOX administration also exerted an influence on the main SCFA concentrations. Propionate and butyrate concentrations significantly decreased with 18% (p < 0.01) and 33% (p < 0.001), respectively, during the administration of 4 mg/L DOX. Administration of 16 mg/L DOX resulted in a significant decrease of propionate and butyrate with 14% (p < 0.05) and 18% (p < 0.001), respectively, while 1 mg/L DOX administration resulted in an average 35% (p < 0.001) decrease of butyrate concentration. Acetate was the main SCFA of anaerobic fermentation but did not significantly decrease upon continuous administration of either one of the DOX concentrations (Fig. 2).
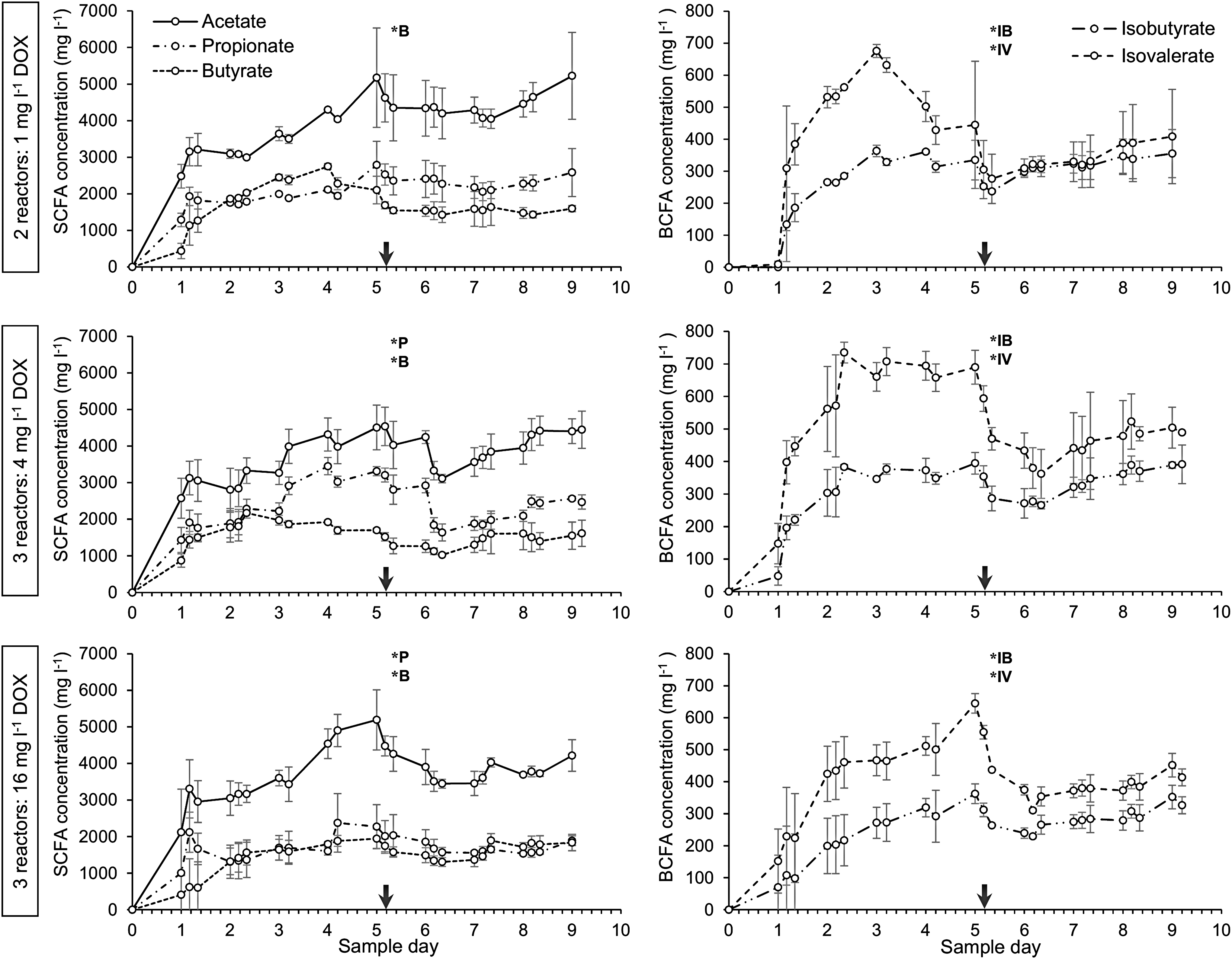
Mean concentrations (mg/L) of main short chain fatty acids (left): acetate, propionate, and butyrate and branched chain fatty acids (right): isobutyrate and isovalerate. Means and standard deviations were calculated for 2–3 replicate reactor runs. Group 1 (top): reactor runs 1, 3 received 1 mg/L DOX. Group 2 (middle): reactor runs 4, 5, 6 received 4 mg/L DOX. Group 3 (bottom): reactor runs 7, 8, 9 received 16 mg/L DOX. The starting point of antibiotic administration is indicated by a vertical arrow. Significant short-term decrease of fatty acid concentrations, due to continuous administration of DOX, is indicated by an asterisk (*). P, propionate; B, butyrate; IB, isobutyrate; IV, isovalerate.
Microbial community
The bacterial community structure of the ex vivo ecosystem was investigated with metabarcoding. Changes in richness (observed number of OTUs) and diversity (Shannon and Simpson index) measures could not be consistently attributed to DOX administration (Supplementary Fig. S2). On average, the main phyla of Bacteroidetes, Firmicutes, and Proteobacteria represented 47% (±6%), 39% (±5%), and 6% (±5%) of the reactor community before antibiotic treatment. The Enterobacteriaceae, a family within the Proteobacteria, making up 0.9% (±0.6%) of the community was mainly represented by the genus Escherichia-Shigella. Taxonomic analysis of the bacterial communities identified a core set of eight families: the Prevotellaceae, Ruminococcaceae, Lachnospiraceae, Bacteroidaceae, Acidaminococcaceae, Erysipelotrichaceae, Enterobacteriaceae, and Succinivibrionaceae. This dominant population was consistent between all reactor runs and represented 85% of the reads in all samples. The remaining families were prone to day-to-day fluctuation within each reactor run (Supplementary Fig. S3) and had variable relative abundances between reactor runs. To study differences between family level composition of the reactor communities, taking into account the relative abundances, samples were ordinated in a two-dimensional plot using NMDS analysis (Fig. 3). Samples of the inoculum and samples collected on the second day of each reactor run clustered closely together. Samples of subsequent days were spread in the NMDS plot, where independent reactor runs displayed different positioning. Overall, there was no consistent increase or decrease of any taxonomic group as a result of 1, 4, or 16 mg/L DOX administration, which was observed in all of the replicate runs per DOX treatment.

NMDS profile of pairwise community dissimilarity (Bray–Curtis) indices of family level abundance data of samples from eight reactor runs and three replicates of the inoculum. The symbols make a distinction between inoculum samples and samples collected before DOX administration, and during the continuous administration of 1, 4, or 16 mg/L DOX. The clustering of inoculum samples (1) and eight reactor samples collected on day 2 (2) are indicated with spider graphics. 95% confidence ellipses were constructed for each of the reactor runs. NMDS, nonmetric multidimensional scaling.
Abundances of total and doxycycline-resistant anaerobes, Enterobacteriaceae and E. coli
The blanc reactor run without the addition of DOX revealed that the number of sensitive and resistant Enterobacteriaceae remained quite stable during the experiment of 9 days (data not shown). This indicates that effects on microbial counts in the other runs where DOX was administered can be attributed to the addition of DOX and not to the reactor run itself. To determine the effect of 1, 4, and 16 mg/L DOX on the proliferation of resistant bacteria compared to the pretreatment period without DOX, reactor content was plated on agar for the enumeration of total and resistant CFUs of E. coli, Enterobacteriaceae and cultivable anaerobes (Fig. 4). During continuous administration of the lowest DOX concentration (1 mg/L DOX), only minor effects were observed compared to the control period of the reactors. The total E. coli count halved from 4.6 × 10 7 to 2.1 × 10 7 CFUs (p < 0.001) while the resistant E. coli count increased from 1.2 × 10 4 to 5.6 × 10 4 CFUs (p < 0.001). No significant effects were found for the enumerations of resistant and total Enterobacteriaceae and anaerobes (averages of two reactor runs). Administration of 4 or 16 mg/L DOX had a more pronounced effect on the proliferation of resistant bacteria. Resistant E. coli significantly increased from 1.1 × 10 2 to 1.1 × 10 5 CFUs (p < 0.001) and from 8.0 × 10 2 to 5.9 × 10 5 CFUs (p < 0.001), respectively, while resistant Enterobacteriaceae significantly increased from 4.4 × 10 5 to 2.5 × 10 6 CFUs (p < 0.001) and from 8.5 × 10 4 to 1.1 × 10 6 CFUs (p < 0.001), respectively (averages of three reactor runs). Total cultivable anaerobe counts were more variable between reactor runs. The resistant cultivable anaerobes slightly increased from 7.9 × 10 7 to 8.1 × 10 8 CFUs (p < 0.001) and from 1.3 × 10 8 to 4.3 × 10 8 CFUs (p < 0.01), respectively (averages of three reactor runs).
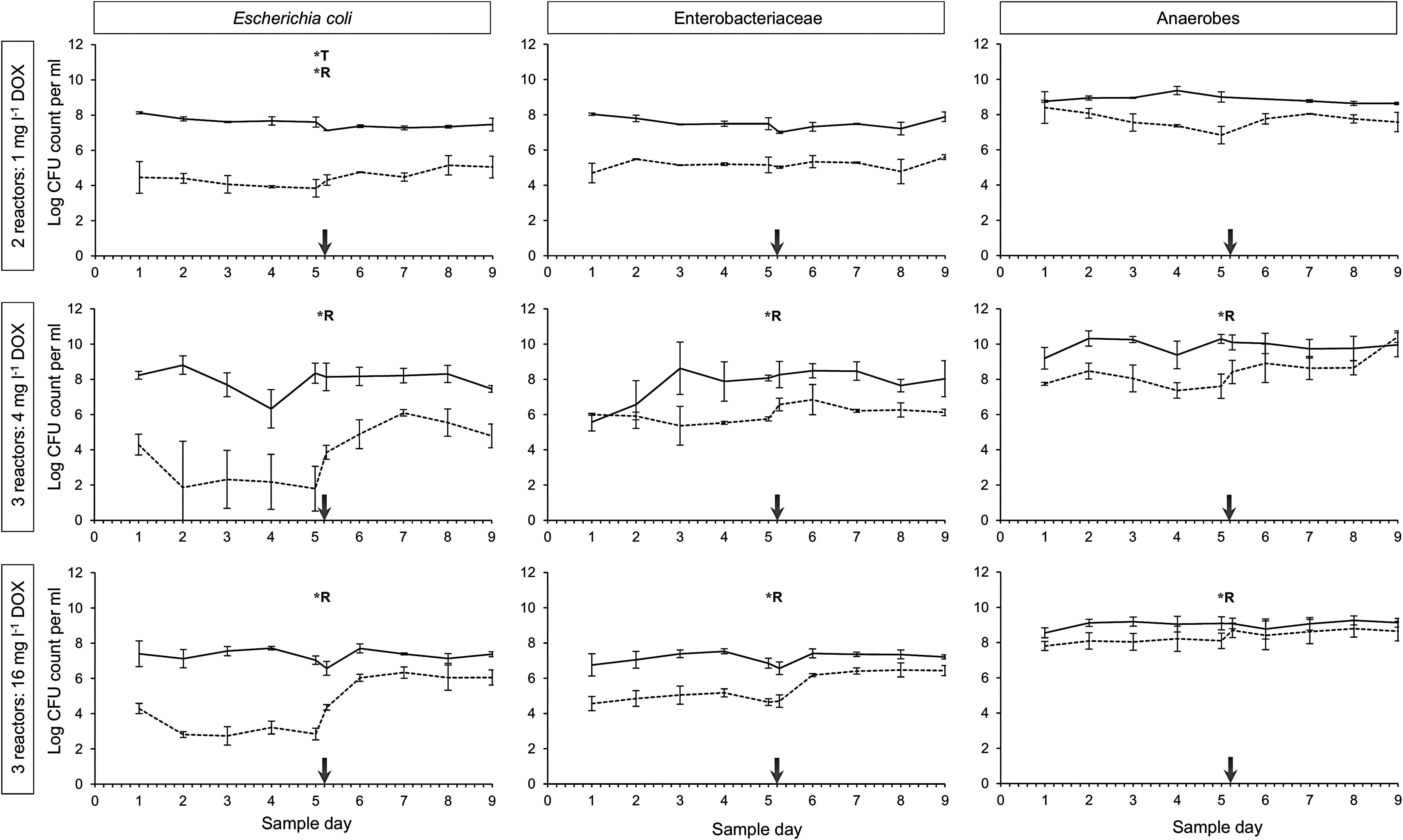
Mean enumerations of total (full line) and doxycycline-resistant (dashed line) Escherichia coli, Enterobacteriaceae, and cultivable Clostridia given in log10 CFU counts per milliliter reactor content. Means and standard deviations were calculated for two to three replicate reactor runs. Group 1 (top): reactor runs 1, 3 received 1 mg/L DOX. Group 2 (middle): reactor runs 4, 5, 6 received 4 mg/L DOX. Group 3 (bottom): reactor runs 7, 8, 9 received 16 mg/L DOX. The starting point of antibiotic administration is indicated by a vertical arrow. A significant increase/decrease in colony count per milliliter (T = total colony count, R = doxycycline-resistant colony count on agar containing 10.5 mg/L DOX) due to continuous administration of DOX and determined with a linear mixed-effects model, is indicated with a asterisk (*). CFU, colony forming units.
Abundances of specific resistance genes
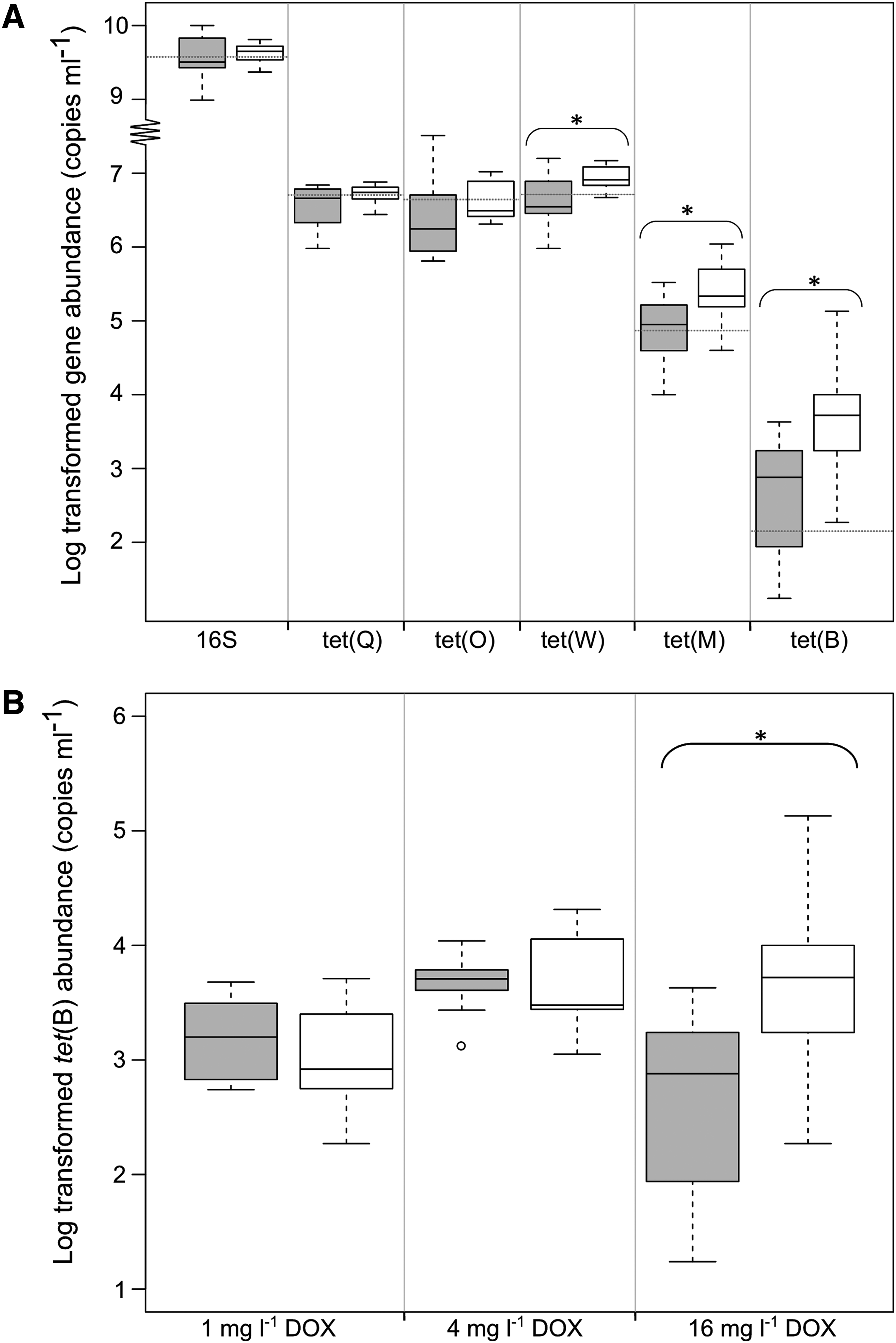
The quantification of tetracycline resistance genes and total 16S rRNA gene abundances was carried out to evaluate the influence of DOX administration on the abundance of specific resistance genes and on the total bacterial load in samples collected during each reactor run and in the cecal inoculum. The tetracycline resistance genes tet(Q), tet(O), tet(W), tet(M), and tet(B) were chosen because they are widespread and often detected with high abundances in agricultural environments23,33–35 and occur in several bacterial groups of the gut microbiome. 31 The samples collected during the pretreatment control period of the reactor runs had similar copy numbers of 16S genes and tet genes as detected in the inoculum samples (Fig. 5A): an average (±SD) bacterial load of 7.3 (±2.5) × 10 8 16S gene copies mL–1, 3.5 (±2.0) × 10 6 tet(Q) genes mL–1, 2.3 (±8.6) × 10 6 tet(O) genes mL–1, 4.5 (±4.7) × 10 6 tet(W) genes mL–1, 7.5 (±9.4) × 10 4 tet(M) genes mL–1, and 1.4 (±2.6) × 10 3 tet(B) genes mL–1. The administration of the highest concentration (16 mg/L DOX) resulted in a near doubling of tet(W) copy numbers, that is, an increase with 3.7 × 106 gene copies mL–1 (p < 0.01). The copy numbers of tet(M) significantly increased with 1.7 × 105 gene copies mL–1 (p < 0.01) and tet(B) gained 3.6 × 103 gene copies mL–1 (p < 0.001), representing a 3- and 10-fold increase, respectively. No significant changes were observed for tet(Q) and tet(O). The administration of lower concentrations (1 or 4 mg/L DOX) did not induce a significant increase of tet(Q), tet(O), tet(W), tet(M) (data not shown), or tet(B) (Fig. 5B).
Discussion
Carry over of antimicrobial compounds from medicated feed to nonmedicated feed results in the presence of subtherapeutic concentrations of antimicrobials in the nonmedicated feed. When pigs consume these cross-contaminated feed, such an antimicrobial compound can accumulate in the compartments of the intestinal tract where it might exert an influence on the microbial community and activity and impose a selection pressure for resistant bacterial species or strains, which could spread to the environment and the wider public via the use of fecal material as manure, animal handling, and carcass contamination. The objective of this study was to investigate the effects of such cross-contamination doses of DOX. One and 4 mg/L DOX correspond to the in vivo intestinal concentration range upon feeding with a compound feed containing 3% of a MRD. 10 These concentrations, and a positive control concentration of 16 mg/L DOX, were administered to the complex microbial community of an ex vivo continuous fermentation model, mimicking the chemical and physical characteristics of the pig cecum. The bioreactors were inoculated with the cecal content of organically grown pigs (who did not receive antibiotic treatments during growth) to investigate the effect of DOX on a gut microbial community that has not yet been into contact with antimicrobial compounds. The use of bioreactors to study the microbiota of animals is an interesting alternative for in vivo experiments as these are hampered by high costs and ethical concerns. Obviously, in vitro or ex vivo models are limited in their capacity to mimic an animal in all his facets, for example, the role of the gut epithelia, or the influence of the immune system on the gut flora. Therefore, it could be questioned if bioreactors are a worthy alternative for in vivo experiments and if the community in the bioreactor reflects the true microbiome of the pig's gastrointestinal tract. Although, bioreactors were already used in the past, it is only recently that the microbiome of bioreactors and pigs can extensively be compared by means of sequencing the DNA of all members in both communities. For this particular experiment, De Mulder 36 demonstrated that the majority of the OTUs were shared between the reactor communities and the in vivo ceca of the pigs that were used as inoculum. The OTUs not included in the reactor microbiome represented less than 20% of the community. Further, the relative abundance of the communities in the bioreactor was in agreement with previous in vivo studies. 37 For example, we found the reactor microbiome dominated by Bacteroidetes and Firmicutes (>90%) and only found a low abundance of the Enterobacteriaceae (<1%) which was also reported in several in vivo studies. 37 This demonstrates that the bioreactor is a worthy alternative for in vivo trials.
Doxycycline inhibits protein synthesis of susceptible bacteria by binding on the 30S subunit of the ribosome and preventing the attachment of aminoacyl-tRNA. Consequently, doxycycline is a broad-spectrum antibiotic, exerting a bacteriostatic effect. The fatty acid profiles suggest that even the administration of low concentrations of DOX (1 mg/L) can cause inhibition of protein synthesis of sensitive bacteria, resulting in an overall reduced metabolic activity and lower production of metabolic end products. The steep reductions of BCFA concentrations, compared to SCFA, are likely due to the 10-fold lower concentrations of BCFAs causing a more pronounced reduction. Another possible explanation might be the variable effect of doxycycline on the solid adherent and the liquid environments of the ecosystem. The presence of solid adherent bacteria is evidenced by the detection of known 38 cellulolytic genera such as Ruminococcus (1.8% ± 3.3%) and Fibrobacter (2.3% ± 3.7% of the reads) using metabarcoding. Microbial attachment and the development of biofilms on the surface of insoluble fibers is the driving factor behind carbohydrate fermentation 39 that accounts for a large fraction of the acetate, propionate, and butyrate concentrations. 40 Presumably solid adherent bacteria are, to a certain extent, protected against doxycycline (this hypothesis is further explored in Supplementary Data S4). On the other hand, proteolytic activity is primarily attributed to free-living bacteria, proliferating on soluble nitrogen sources in the feed medium and generating a complex mixture of metabolic end products, including SCFAs (acetate, butyrate, and propionate) and BCFAs (isobutyrate and isovalerate). 41 These free-living bacteria are more susceptible to antibiotics, 42 which could also have contributed to the steep decrease of BCFA production following DOX administration.
In contrast to the effects of DOX on the metabolic activity, alpha diversity calculations and ordination of the samples based on OTU abundance profiles suggest there was no influence of DOX administration on the microbial community structure. In line with these results, no taxonomic group was found to significantly increase or decrease during continuous DOX administration. Similarly, Holman and Chénier demonstrated that the microbial diversity and community structure of fecal samples were not affected by administering a subtherapeutic dose of 5.5 mg chlortetracycline per kilogram feed to weaning pigs. 43 Presumably, selection pressure imposed by doxycycline will cause resistant bacteria to proliferate and replace sensitive species within the major taxonomic groups. In addition, adherent bacteria and bacterial aggregates could contribute to the stability of the community during antibiotic stress. Therefore, our findings suggest that doxycycline administration only exerts a minor influence on the bacterial community composition. However, that does not mean that DOX has no influence on the bacterial community, as metabarcoding can only detect shifts in relative abundance on OTU level and is not able to detect shifts on strain level, nor in absolute abundance. When resistant strains outcompete susceptible strains within the same species, this will not be picked up by metabarcoding.
Plate cultivation on agar with DOX indicated a presence of around 102–104 tetracycline-resistant E. coli per milliliter (representing 0.0001–0.02% of the total enumerated E. coli counts) and around 107–108 tetracycline-resistant cultivable anaerobes per milliliter (representing 0.7–13% of the total enumerated anaerobes) in the reactors during the pretreatment control period. This was remarkable as there was no selective pressure imposed by antibiotics during the pretreatment period nor during the growth of the organically grown pigs whose cecal content were used as inoculum of the bioreactors. In comparison, 40.4% of E. coli isolated from fecal samples from organically grown Tibetan pigs (with a complete absence of antibiotic use) displayed resistance against tetracycline. 44 It would seem that tetracycline-resistant species are an inherent part of the microbial communities in the pig's GIT. Despite the high occurrence of tetracycline resistance in the intestinal microbial ecosystem of pigs (even in organically grown pigs), selective plate cultivation in our experiments provided strong and consistent evidence of the influence of DOX on the enrichment of tetracycline-resistant species in the complex and dynamic microbial communities of the reactor system. Notably, when focusing on a single species such as E. coli, a generally recognized indicator organism for tracking microbial resistance 45 we observed a profound effect of DOX administration on the enrichment of resistant CFUs. In contrast, the effects of DOX administration appeared smaller when larger taxonomic groups were enumerated on less specific agar plates, such as the family of Enterobacteriaceae on VRBGA and cultivable anaerobes on RCA, as these plates support the growth of multiple species and each species and even strains belonging to the same species can respond differently to the effects of DOX. Nevertheless, plate cultivation suggest that low DOX concentrations, can cause significant enrichment of tetracycline-resistant species, especially species that form a potential threat to human and animal health, for which E. coli is an indicator. This corresponds to the results of Peeters et al. where the same low DOX concentrations, that is, 1 and 4 mg/L, were able to select for a DOX-resistant E. coli strain and to promote the rate of conjugation in a similar bioreactor setup. 46
In accordance with the high prevalence of resistant CFUs during the pretreatment period of the reactor runs, qPCR assays also indicated high concentrations of specific tet genes during the pretreatment period. Within a community of around 109 gene copies per milliliter of eubacterial 16S rDNA, around 106 gene copies per milliliter of tet(Q), tet(O), and tet(W), 105 gene copies per milliliter of tet(M) and 10 3 gene copies per milliliter of tet(B) were detected. Furthermore, the bioreactors contained similar levels of tet(Q), tet(O), tet(W), and tet(M) during the pretreatment period of the reactors as detected in the inoculum prepared from the pooled cecal content of organically grown pigs. These results suggest that tetracycline resistance is omnipresent in a gut environment. Long-term persistence and ubiquity of tetracycline resistance genes in the absence of antibiotic use was also previously suggested.47,48 However, in contrast to plate cultivation, the copy numbers of tet genes did not increase upon DOX administration of 1 and 4 mg/L, and only 16 mg/L caused statistically significant increases of tet(W), tet(M), and tet(B). These findings indicate a discrepancy between classic plate cultivation of viable bacteria and the molecular quantification of genes. This can have several explanations; first, on plates only those cultivable organisms can grow expressing the resistance gene, whereas qPCR detects resistance genes even when they are not expressed. Second, one isolate can harbor several copies of a certain resistance gene, for example, when located on both the chromosome and/or different plasmids, or located on a plasmid that is present in a low-, medium-, or high copy number. Third, this could also be attributed to the limited number of tet genes investigated in this study. Five tet genes were selected and quantified because of their frequent occurrence in agricultural environments with antibiotic use.33–35 Tet(Q), tet(O), tet(W), and tet(M) encode ribosomal protection proteins and are found in both Gram-positive and Gram-negative bacteria. Tet(B) encodes a widely distributed Gram-negative tetracycline efflux pump 49 but is mainly associated with species of the Proteobacteria.31,32 However, there is a large number of known (and yet unknown) tet genes that were not included in this study. The effect of DOX administration on the abundance of a single tet gene might be more limited because numerous other resistance genes and/or mutations may contribute to resistance development, thus spreading the effect across a large number of genes. It is therefore possible that DOX administration only induces a limited individual increase in the abundance of a large number of tet genes.
Plate cultivation and qPCR are commonly used techniques in research on antimicrobial resistance, but our results emphasize some shortcomings of these methods. Agar plate cultivation enumerates total and resistant cultivable bacteria, but lacks in information about the taxonomy and the antimicrobial resistance determinants they possess. Furthermore, only a small fraction of the community is cultivable. On the other hand, each qPCR assay is specific for a resistance gene or a group of genes, which may or may not be expressed. The researcher must make a reasoned selection of which genes to study, with the risk of overlooking other important contributors. In addition, qPCR only provides quantitative information of gene levels, but not about host association. Developments in the field of metagenomics can address these shortcomings. Hi-C sequencing, based on DNA cross-linking in living cells before NGS library preparation, is able to reliably associate plasmids with each other and with the chromosomal DNA of the host cell. 50 A recently described technique called epicPCR isolates single bacterial cells in emulsion beads and uses fusion PCR to physically link specific functional and phylogenetic genes before amplicon sequencing. 51 In the near future, these novel techniques could be used to correlate taxonomic classification and resistance mechanisms of the species in the community.
In conclusion, we investigated the influence of subtherapeutic DOX concentrations, as a consequence of cross-contamination in the feed, on the microbial ecosystem of the pig cecum simulated with an ex vivo bioreactor model. When cross-contamination results in a concentration of 3% or more of the therapeutic dose, there is a significant effect on the enrichment of resistant bacteria and specifically resistant E. coli. As specific tet genes were already abundant in the ceca of organically grown pigs used as inoculum and in the blanc pretreatment control phase of each reactor run, only the highest DOX concentration tested led to a small increase in abundance of the investigated tet genes. On the other hand, the microbial activity, indicated by the fatty acid concentration, decreased significantly for each DOX concentration tested, whereas the taxonomic profile of the simulated bacterial cecal community was not influenced by DOX administration.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.