
Abstract
Quinolones show excellent antibacterial activity against Salmonella isolates. Recently, however, quinolone resistance has been increasing in bacteria. This study aimed to examine in vitro, and compare the activity of DC-159a against Salmonella enterica serovar Typhimurium with that of ciprofloxacin and nalidixic acid. Inhibitory effects of quinolones were assessed by the drug concentration needed to inhibit the supercoiling activity of recombinant DNA gyrases by 50% (IC50). Dilution methods were used to determine the minimum inhibitory concentration (MIC) of quinolones against two different strains, Salmonella Typhimurium and Salmonella Enteritidis. The IC50s of DC-159a against mutant DNA gyrases were much lower than those of nalidixic acid and ciprofloxacin. In particular, the IC50 of DC-159a against DNA gyrase with double mutation was less than 1/50 that of ciprofloxacin and nalidixic acid. MICs of DC-159a were higher than those of ciprofloxacin but lower than those of nalidixic acid. However, the estimated MICs of DC-159a against Salmonella strains with mutant DNA gyrase were lower than those of ciprofloxacin and nalidixic acid. Therefore, DC-159a can be suggested as an antibiotic candidate for treating salmonellosis caused by quinolone-resistant S. Typhimurium.
Introduction
T
DNA gyrase is a bacterial enzyme composed of A and B subunits encoded by gyrA and gyrB, respectively.9,10 Two GyrA and two GyrB form a functional tetrameric protein, and DNA gyrase, essential for protein synthesis and DNA replication, unwinds bacterial DNA by introducing negative supercoils.10,11 Inhibition of DNA gyrase functions is, therefore, one of the most important bactericidal effects of quinolones.9,12 However, mutations arising from the binding sites of quinolones in gyrA, known as the quinolone resistance determining regions (QRDRs), mediate quinolone resistance.13–15 Indeed, amino acid substitutions in QRDR induce conformational changes and decreased binding affinity of quinolones.15,16 Quinolone-resistant mutations frequently occurring at positions 83 and 87 in QRDR suggest that these residues may be involved in quinolone binding.17–19 For example, amino acid substitutions in GyrA at positions 83 (Ser83Phe) and 87 (Asp87Asn, Asp87Gly, and Asp87Tyr) have been frequently reported as quinolone resistance-related mutations.20–22 Moreover, a clinical isolate with double mutation Ser83Phe-Asp87Asn was reported to have a higher level of resistance to quinolones than did those with single mutations. 23 Hence, it is imperative to find an effective compound against bacteria carrying DNA gyrases with the aforementioned amino acid substitutions.
Nalidixic acid is the first-generation quinolone antibiotic, but it only shows modest activity against gram-negative bacteria. 24 Quinolones are synthetic antibiotics with various structural modifications. 25 Since the original production of nalidixic acid, a series of subsequently produced quinolones led to the introduction of fluoroquinolones. 26 Fluoroquinolone is a quinolone that contains a fluorine atom at position C6 of the central ring (Fig. 1). This attachment drastically improves its antibacterial activity and as a result, its antibacterial spectrum becomes much wider than that of nalidixic acid. 27 Ciprofloxacin is a second-generation fluoroquinolone with a broader spectrum of bactericidal activity. 28 This antibiotic is used for treatment of infectious diarrhea, and was selected by the WHO as an essential medication for both adults and children. 29 Nonetheless, although ciprofloxacin is a useful antibiotic with high effectiveness, strains resistant to ciprofloxacin have also been recently isolated in many countries.8,30,31
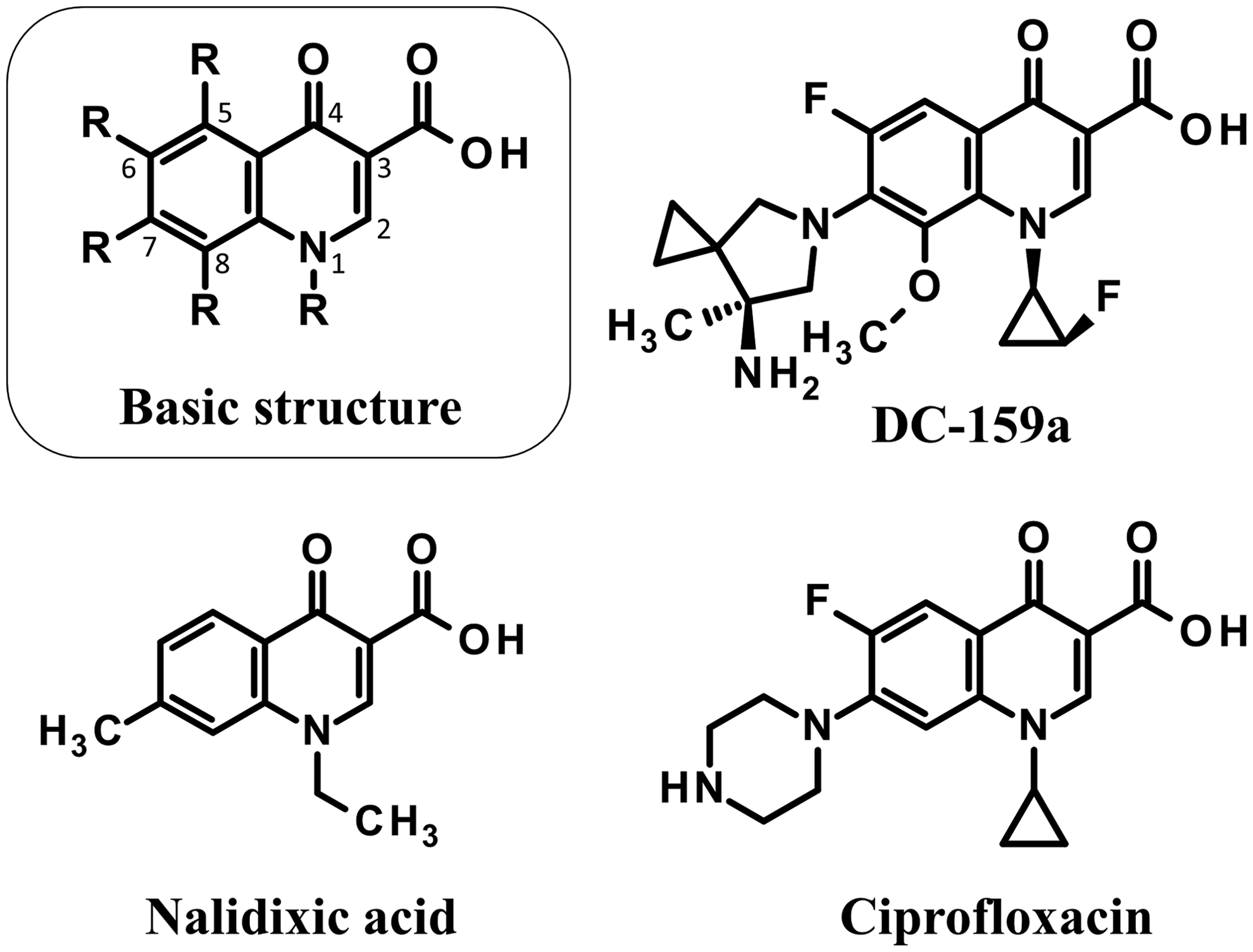
Chemical structures of quinolones used in this study.
DC-159a is a fluoroquinolone developed in Japan. The potency of DC-159a has already been examined in previous studies and proven effective against various pathogens.32–35 Therefore, this study sought to confirm an expected inhibitory activity of DC-159a against Salmonella Typhimurium exhibiting quinolone resistance. In addition, this study aimed to examine in vitro, and compare the antibacterial activity of DC-159a against nontyphoidal Salmonella with that of ciprofloxacin and nalidixic acid.
Materials and Methods
Quinolones
In a previous assay of multiple quinolones, 36 ciprofloxacin, a very important antibiotic from a clinical standpoint, 29 showed lower IC50 than other quinolones. In addition, nalidixic acid is routinely used for screening quinolone-resistant bacteria. 37 Based on this evidence, ciprofloxacin and nalidixic acid were used in this study to evaluate the antibacterial activity of DC-159a.
DC-159a was provided by Daiichi Sankyo Co., Ltd. (Tokyo, Japan). Ciprofloxacin and nalidixic acid were purchased from LKT Laboratories, Inc. (St Paul, MN) and Wako Pure Chemical Industries, Ltd. (Osaka, Japan), respectively. Together with the basic structure of quinolones, the chemical structures of the aforementioned quinolones are shown in Figure 1.
Protein expression and purification
Recombinant DNA gyrase was obtained as separate subunits, GyrA and GyrB. Seven subunits in total were produced in this study. Expression plasmids, including the coding regions of GyrA and GyrB of S. Typhimurium, were previously constructed. 36 Mutations Ser83Phe, Asp87Asn, Asp87Gly, Asp87Tyr, and Ser83Phe-Asp87Asn were introduced into QRDR. The plasmids were then introduced into Escherichia coli BL21(DE3) (Merck KGaA, Darmstadt, Germany), and protein expression was induced in the bacterial cells. A colony of transformed E. coli was inoculated into Luria-Bertani broth with ampicillin (1 μL/mL) and incubated at 37°C until the optical density value reached 0.60. Protein expression was induced by adding 1 mM isopropyl-β-D-thiogalactopyranoside (IPTG), purchased from Wako Pure Chemical Industries, Ltd. The incubation period of recombinant E. coli carrying the GyrA expression plasmid was 40 hr, and the temperature after adding IPTG was 16°C. Separately, recombinant E. coli carrying GyrB was incubated for 13 hr at 18°C. Harvested E. coli was centrifuged, and then sonicated at a 30% duty cycle with 10 cycles of 40 sec on and 40 sec off (Sonifier® 250; Branson, Danbury, CT) to release the expressed protein. The recombinant DNA gyrase subunits were then purified by Ni-NTA agarose column chromatography and dialyzed against DNA gyrase dilution buffer (50 mM Tris-HCl, pH 7.5; 100 mM KCl, 2 mM dithiothreitol [DTT], 1 mM ethylenediaminetetraacetic acid [EDTA]). To avoid denaturation, glycerol was added to the obtained protein. Finally, the mixture was stored in small aliquots at −80°C until further use. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was used to assess the purity of proteins.
DNA supercoiling activity and inhibitory assay
Supercoiling activity of purified DNA gyrases was detected using agarose gel electrophoresis.36,38 Relaxed pBR322 (302.4 ng) was incubated with DNA gyrases (GyrA: 30.6 ng, GyrB: 28.3 ng) in gyrase assay solution (35 mM Tris-HCl, 6 mM MgCl2, 1.8 mM spermidine, 24 mM KCl, 5 mM DTT, 0.36 mg/mL BSA, and 6.5% glycerol [w/v]). After 40-min incubation at 35°C, the reaction was stopped with 8 μL of a stop solution (5% SDS, 25% glycerol, and 0.25 mg/mL bromophenol blue). 39 Next, 10 μL of the reaction mixture was loaded onto a 1% agarose gel in 0.5 Tris-borate-EDTA (TBE) buffer for 2-hr electrophoresis at 40 mA. Gels were stained using 0.5 TBE buffer with 5 μg/mL of ethidium bromide, and the supercoiled DNA bands were visualized under UV light.
A DNA supercoiling inhibitory assay was conducted to identify and quantify quinolone inhibition of DNA gyrases. The procedure was almost the same as that described above for the supercoiling activity assay, except that this time it was conducted in gyrase assay solution with serially diluted quinolones. To examine the supercoiling inhibitory effects of quinolones on recombinant DNA gyrases, brightness of bands was measured by software ImageJ (http://rsbweb.nih.gov/ij). All enzyme assays were run in triplicate to eliminate experimental bias and to confirm reproducibility. After that, the obtained data were first plotted by the quinolone concentration, and subsequently the brightness of the quinolone-free sample was defined as 100% assay control. The relationship between brightness and the quinolone concentration was explained by fitting a four-parameter logistic model. 40 Inhibitory effects were evaluated by IC50, which was defined as the drug concentration necessary to reduce by 50% the supercoiling activity observed in agarose gels in comparison with that in quinolone-free samples.32,36,38 Thus, IC50 was determined from the aforementioned model. The above calculation was conducted using statistical software R version 3.2.5.
Antimicrobial susceptibility testing
Minimum inhibitory concentration (MIC) was determined by the broth dilution method. S. Typhimurium NBRC 13245 and S. Enteritidis NBRC 3313 were tested for antimicrobial susceptibility in accordance with the procedure of the Clinical and Laboratory Standards Institution (CLSI). 41 The suspension of each strain was transferred to a 96-well tray containing the serially diluted quinolones. After overnight incubation of the suspensions at 37°C, the MIC was defined as the lowest concentration of quinolone that completely inhibited all visible bacterial growth in a well. This examination was run in triplicate for each strain.
Estimation of MICs against mutant DNA gyrase carrying S. Typhimurium and S. Enteritidis
Based on the data from a previous study of Salmonella Typhimurium DNA gyrase, it was estimated that there was a correlation between the different quinolones. 38 Moreover, as DC-159a showed the same logarithmic correlation between the IC50 and the corresponding MIC, it was possible to estimate the MICs of DC-159a against mutant strains with single- or double-mutant DNA gyrases from the results of the supercoiling inhibitory assay. Regression lines were calculated based on log(IC50) and log(MIC) of the three quinolones against wild types (WTs) using S. Typhimurium and S. Enteritidis in a separate manner. Next, log(MIC) was calculated from the regression coefficient and log(IC50), and the estimated MIC was determined by converting the variable log(MIC) from logarithm to integer.
Results
Supercoiling activity of recombinant DNA gyrases
The GyrA and GyrB subunits of S. Typhimurium were purified separately by affinity chromatography. Successful purification of GyrA and GyrB at 97 and 89 kDa, respectively, was confirmed by SDS-PAGE (Fig. 2 and Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/mdr). Supercoiling activity was observed when GyrA and GyrB were combined and in the presence of adenosine triphosphate (ATP). This activity was not observed when GyrA, GyrB, or ATP was absent (Fig. 3).

SDS-PAGE analysis of purified DNA gyrase subunits of Salmonella Typhimurium. GyrA and GyrB were purified separately by Ni-NTA affinity chromatography. Three hundred nanograms of each purified protein was loaded onto a 5–20% gradient polyacrylamide gel. Lanes: M, protein marker; 1, WT-GyrA; 2, S83F-GyrA; 3, D87N-GyrA; 4, D87G-GyrA; 5, D87Y-GyrA; 6, S83F-D87N-GyrA; 7, WT-GyrB. SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis. WT, wild type.
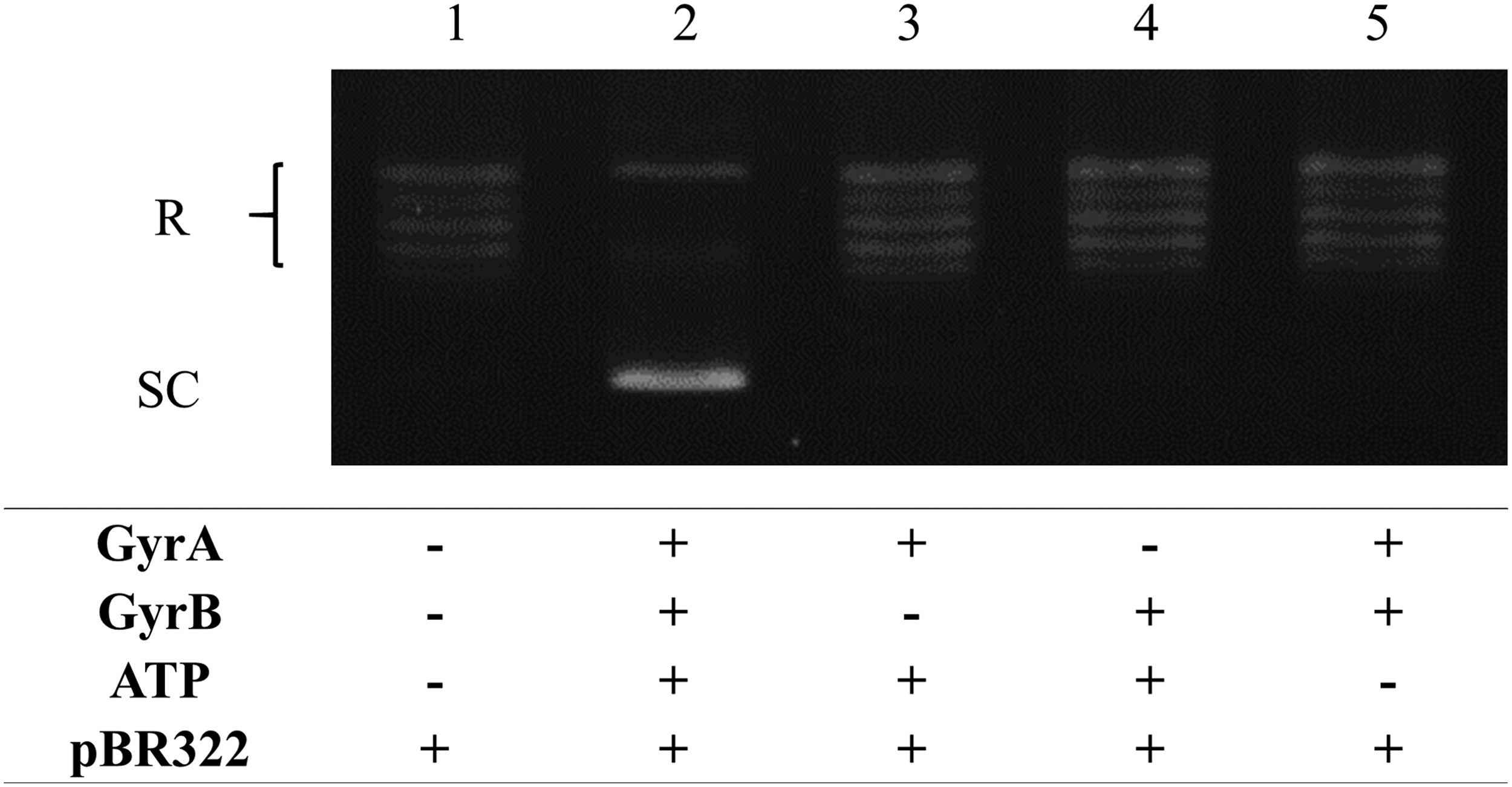
ATP-dependent supercoiling activity of WT GyrA and GyrB proteins of S. Typhimurium. R; relaxed pBR322 DNA. SC; supercoiled pBR322 DNA.
IC50s of quinolones
Inhibition activity of DC-159a against WT and mutant DNA gyrases of S. Typhimurium was estimated by the IC50, and compared with that of ciprofloxacin and nalidixic acid. Dose-dependent inhibition activity of every quinolone within the 0.25–600 μg/mL range was confirmed. The inhibitory effects of tested quinolones are shown in Figure 4. The IC50s of each quinolone are summarized in Table 1. The IC50s of DC-159a and ciprofloxacin against WT DNA gyrases were 0.32 μg/mL (95% confidence interval [95% CI]: 0.25–0.40 μg/mL) and 0.25 μg/mL (95% CI: 0.19–0.32 μg/mL), respectively. There was no significant difference between them. However, DC-159a showed more potent activity against mutant DNA gyrases than did ciprofloxacin. Furthermore, the IC50s of DC-159a against mutant DNA gyrases were significantly different from those of ciprofloxacin. The biggest differences between the IC50s of DC-159a and ciprofloxacin were observed in double-mutant DNA gyrases. For example, the IC50 of DC-159a against the DNA gyrase bearing double mutation Ser83Phe-Asp87Asn was less than 1/55 that of ciprofloxacin. Moreover, while the IC50 of DC-159a against double-mutant DNA gyrases showed a 27.5-fold increase in comparison with that against WT DNA gyrase, the IC50 of ciprofloxacin against double-mutant DNA gyrase showed a 1,920-fold increase when compared with that against WT. In contrast, the concentration of nalidixic acid required to achieve supercoiling inhibition activity against every WT and mutant DNA gyrase was much higher than those of DC-159a and ciprofloxacin.

Supercoiling inhibitory assay. It was observed that as the concentration of quinolone increased, the density of the supercoiled DNA band became less bright. These figures are randomly selected from a triplicate assay.
IC50s of Quinolones Against WT and Mutant DNA Gyrases
(n), the value in brackets, is the n-fold increase in IC50s against mutant DNA gyrases compared with that against WT DNA gyrase.
WT, wild type.
Minimum inhibitory concentrations
To examine and compare the bactericidal effect of DC-159a with that of ciprofloxacin and nalidixic acid, MICs against S. Typhimurium and S. Enteritidis standard strains were measured. The results of antimicrobial susceptibility testing are shown in Table 2. MICs of DC-159a against these two strains were different, with the growth inhibitory activity of DC-159a being higher than that of nalidixic acid but lower than that of ciprofloxacin.
Minimum Inhibitory Concentrations for Salmonella Typhimurium and Salmonella Enteritidis
Dilution method was used to determine the MIC of DC-159a, ciprofloxacin, and nalidixic acid against S. Typhimurium and S. Enteritidis.
MIC, minimum inhibitory concentration.
Estimated MIC
MICs of DC-159a, ciprofloxacin, and nalidixic acid against mutant S. Typhimurium and S. Enteritidis were estimated from the results of IC50 and MIC against standard strains. The estimated values are summarized in Table 3. In CLSI standards, the breakpoint of ciprofloxacin for Salmonella spp. is classified as follows: ≤0.06 μg/mL, susceptible; 0.12–0.5 μg/mL, intermediate; and ≥1.0 μg/mL, resistant. 37 In this work, MICs of ciprofloxacin were estimated to range from 0.33 to 0.60 μg/mL for strains with single-mutant DNA gyrases. Thus, single-mutant strains were categorized as intermediate. Elsewhere, a wider range of MICs were observed for Salmonella isolates having single-mutant DNA gyrases.42,43 The MIC of DC-159a for single-mutant strains was less than half that of ciprofloxacin. Moreover, although the concentration of ciprofloxacin was estimated to be exceedingly high for strains having double-mutant DNA gyrases, the MIC of DC-159a against the same DNA gyrases was 52- to 71-fold lower than that of ciprofloxacin.
Estimated Minimum Inhibitory Concentrations for S. Typhimurium and S. Enteritidis
MIC of each quinolone for WT was determined by broth dilution method in accordance with the procedure of the CLSI. Estimated MICs were calculated based on the logarithmic correlation between IC50 and MIC.
CLSI, Clinical and Laboratory Standards Institution.
Discussion
The emergence of nontyphoidal Salmonella infections caused by quinolone-resistant pathogens has become one of the most significant public health concerns worldwide. DC-159a is a newer fluoroquinolone that has already been shown to have potent activity against various microorganisms including some with amino acid substitutions in DNA gyrases.32,33,35 In this study, we examined the potential bactericidal effect of DC-159a against S. Typhimurium by (1) measuring its IC50 and MIC, and (2) comparing its antibacterial activity with that of ciprofloxacin and nalidixic acid.
IC50s of every investigated quinolone against mutant DNA gyrases were higher than those against WT DNA gyrases. Consequently, observed IC50s against double-mutant DNA gyrases were higher than those against single-mutant DNA gyrases. Amino acids 83 and 87 have previously been described as being strongly related to the binding of quinolones to DNA gyrase.17,19 Therefore, the binding affinities of ciprofloxacin and nalidixic acid were expected to drastically decrease due to IC50 differences between WT and mutant DNA gyrases. Nonetheless, the binding affinity of DC-159a was maintained even when amino acids were substituted. Thus, of all quinolones tested by the supercoiling inhibitory assay, DC-159a showed the most potent inhibitory activity against mutant DNA gyrases of S. Typhimurium, especially double-mutant DNA gyrases.
As described above, it is well known that fluorination at C6 of the quinolone ring improves the antibacterial activity. In addition to fluorination at C6, the structure of ciprofloxacin at positions N1 and C7 is different from that of nalidixic acid. N1 belongs to the enzyme–DNA complex, and a cyclopropyl at position N1 in ciprofloxacin is currently considered the most optimal substituent for the formation of an enzyme–DNA binding complex. 44 Position C7 is believed to directly interact with the DNA gyrase, and a bulky side chain at C7 plays an important role in the prolongation of half-lives and the direct interaction with DNA gyrase. 27 Likely due to these mechanisms, the inhibitory activity of ciprofloxacin against DNA gyrases was greater than that of nalidixic acid, and its observed IC50 lower than that of nalidixic acid. In contrast, DC-159a possesses a 3-aminopyrrolidyl substituent at C7. 35 Apart from DC-159a, sitafloxacin is the only quinolone that presents a similar structure at C7.36,45 A previous study reported that the inhibitory activity against mutant DNA gyrases of S. Typhimurium showed by sitafloxacin was relatively stronger than that of ciprofloxacin. 36 Since the IC50 values showed by DC-159a against Mycobacterium leprae DNA gyrases 32 are similar to those shown by sitafloxacin, a substituent at C7 in DC-159a can be considered a key factor for maintaining a strong affinity to mutant DNA gyrases.
Our results from the supercoiling inhibitory assay were in agreement with previous antimicrobial susceptibility testing against clinical isolates.23,46,47 For example, in our supercoiling inhibitory assay, the IC50 of ciprofloxacin against double-mutant DNA gyrases was greater than that against WT DNA gyrases. This result is in concordance with previous work reporting an association between double mutation in GyrA and a high level of resistance in clinical isolates.23,46 In addition, regardless of the mutation in GyrA, a concentration of nalidixic acid higher than that of ciprofloxacin was required to inhibit the activity of DNA gyrases. Nonetheless, the detected IC50s of ciprofloxacin were significantly lower than those of nalidixic acid. These results were also in agreement with previous studies, as it was reported that the MIC of nalidixic acid against clinical isolates of Salmonella Typhimurium was higher than that of ciprofloxacin.46,47 MIC values are considered to be strongly related to IC50s.38,48 The IC50s of DC-159a were lower than those of ciprofloxacin; hence, low MICs of DC-159a against ciprofloxacin-resistant Salmonella strains were also expected.
In the antimicrobial susceptibility testing, MICs of DC-159a against S. Typhimurium and S. Enteritidis were 0.13 and 0.06, respectively (Table 2). According to National Center for Biotechnology Information (NCBI) databases, S. Typhimurium and S. Enteritidis have the same amino acid sequences in GyrA, and this sequence was determined to be a WT. Thus, the susceptibility of their DNA gyrase against quinolones was presumed to be the same. These results suggested that the rate of DC-159a accumulation was different between S. Typhimurium and S. Enteritidis. In the data from MIC estimation, although it was considered that DC-159a permeability and/or accumulation were lower than those of ciprofloxacin, the MIC of DC-159a was lower than that of ciprofloxacin. Interestingly, the MIC of DC-159a against strains with double-mutant DNA gyrases was less than 1/50 that of ciprofloxacin. Salmonella isolates carrying double-mutant DNA gyrases showed a higher level of quinolone resistance than those with a single mutant. It was also estimated that MICs of DC-159a against each mutant strain were much lower than those of nalidixic acid. Therefore, it was clear that DC-159a showed potential to be used as an effective therapeutic agent against salmonellosis caused by quinolone-resistant bacteria.
Quinolones exhibit dose-dependent bactericidal effects. 49 Thus, the maximum concentration of a drug that a living organism can tolerate (Cmax) must be considered. The Cmax of DC-159a in murine models has previously been reported.50,51 Based on those findings, the Cmax values of DC-159a were similar to those of ofloxacin in serum of male juvenile rats receiving a single oral administration. In a separate study, 52 the Cmax of ofloxacin in a healthy human male was examined. The recommended oral dosage of ofloxacin is 200–400 mg twice daily. 53 When a 200 mg tablet of ofloxacin was administered, the Cmax was 1.74 μg/mL. 52 It is therefore expected that if a similar amount of DC-159a (200 mg) was administered to human, the Cmax would be close to 1.74 μg/mL. This DC-159a concentration would suffice to exert a bactericidal effect because the MIC of DC-159a was estimated to be 1.34 μg/mL at most (Table 3). Moreover, since fluoroquinolones are absorbed from the intestine, 54 the concentration of fluoroquinolone in the intestinal tract would be higher than it would be in serum. Based on this evidence, it is believed that DC-159a could exert a potent bactericidal effect against nontyphoidal Salmonella in the intestinal tract even at a lower dose.
Since quinolones are dose-dependent antibiotics, usually high doses of quinolones exert excellent therapeutic effects. 55 However, as the adverse effects are also dose dependent, 56 a quinolone with fewer adverse effects was required. It is believed that the bactericidal activity and the adverse effects are strongly related to the structure of quinolones as well. In addition to the aforementioned structure–activity relationship of quinolones, a substituent at position C8 has been found to be responsible for adverse effects.56,57 Nonetheless, the substituent at C8 in DC-159a is a methoxy moiety, and C8-methoxy quinolones have been reported to have less adverse effects such as low phototoxicity. 56–59 Therefore, a high dose of DC-159a should be readily available, and the antimicrobial activity against quinolone-resistant S. Typhimurium and S. Enteritidis sufficient.
This study demonstrated that DC-159a has two advantages over ciprofloxacin and nalidixic acid: (1) it is more effective at inhibiting the activity of DNA gyrases with reported mutations in QRDR, especially double-mutant DNA gyrases, and (2) it shows a potent antimicrobial activity at lower doses against quinolone-resistant S. Typhimurium and S. Enteritidis. In addition, it will likely cause less adverse effects even if administered at high doses. It can be concluded that DC-159a is a promising antibiotic candidate that is safe and with potent in vitro activity for treating infectious human diseases such as those caused by quinolone-resistant nontyphoidal salmonellae.
Footnotes
Acknowledgments
This work was supported in part by a grant from the Ministry of Education, Culture, Sports, Science and Technology (MEXT), Japan, for the Joint Research Program of the Research Center for Zoonosis Control, Hokkaido University to Y.S., and in part by the Japan Initiative for Global Research Network on Infectious Diseases, Grant No. 15fm0108008h0001 from the Japan Agency for Medical Research and Development (AMED) to Y.S. We are also grateful to Daiichi-Sankyo Co., Ltd. for providing DC-159a.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.