
Abstract
Global emergence of multidrug resistant (MDR) strains limits therapeutic efficacy in Pseudomonas aeruginosa corneal ulcers. Identifying the primary causal factors of resistance shall improve clinical management. In this study, we sought to identify the underlying mechanisms of fluoroquinolone and aminoglycoside resistance in MDR, non-MDR, and drug susceptible P. aeruginosa (n = 19) strains obtained from keratitis patients. Phenotypic assays were performed to study the bacterial growth kinetics, efflux, permeability, and biofilm formation. Mutational alteration of target genes (DNA sequencing), relative expression of efflux system genes (real time PCR), and detection of aminoglycoside modifying enzyme (AME) genes (PCR) were done by molecular methods. We repeatedly found the mutations in quinolone resistance determining region of fluoroquinolone target genes, gyrA and parC, and the presence of AME genes, aph(3″)-I and aph(6)-I, in all MDR isolates. Furthermore, the MDR isolates were largely characterized by slower growth, cytotoxic type III secretion system genotype, better biofilm-forming ability, and the presence of additional AME genes. The non-MDR isolates were resensitized upon inhibition of active efflux or enhancement of membrane permeability. Altogether this study highlights target gene alteration and enzymatic drug modification as the major mechanisms of quinolone and aminoglycoside resistance in P. aeruginosa keratitis isolates.
Introduction
Microbial keratitis is characterized by painful inflammation of the cornea, and the ulcers caused by Pseudomonas aeruginosa are often associated with a poor visual prognosis. 1 Development of multidrug resistance by the pathogen to a broad spectrum of antibiotics poses a major challenge for treatment. Inappropriate usage of antibiotics and prolonged duration of treatment have led to a global increase in the development of drug resistance among ocular pathogens. 2 Apart from the innate chromosomally encoded mechanisms, P. aeruginosa has the ability to acquire resistance genes from the same or different species. 3 Various mechanisms of antimicrobial resistance 4 include target alteration, enzymatic drug inactivation, acquisition of resistance genes, decreased drug influx or increased efflux, and biofilm formation. 5
Previous reports from the steroids for corneal ulcers trial (SCUT) as well as our study on P. aeruginosa endophthalmitis isolates have shown increased fluoroquinolone resistance in cytotoxic P. aeruginosa strains encoding exoU gene of type III secretion system (T3SS).6,7 However, not many studies have addressed the underlying mechanisms of drug resistance in ocular P. aeruginosa isolates. A deeper understanding of specific causative factors may help to curb the spreading rate of antibiotic resistance by devising appropriate treatment strategies. In this study, we sought to identify and functionally characterize the drug resistance mechanisms in corneal isolates of P. aeruginosa. We specifically focused on fluoroquinolone and aminoglycoside resistance, since these two classes of antibiotics are commonly used to treat P. aeruginosa corneal infections.
Furthermore, we extended some of our investigations to drug-sensitive isolates as well to understand the prevalence of these mechanisms in general populations of P. aeruginosa. We found that the mutational alteration of target genes and the acquisition of aminoglycoside modifying enzyme (AME) genes were the predominant causes for quinolone and aminoglycoside resistance in multidrug resistant (MDR) P. aeruginosa isolates. To the best of our knowledge, this is the first comprehensive study to characterize multiple resistance mechanisms in a select group of ocular P. aeruginosa isolates.
Materials and Methods
Bacterial isolates and reagents
The P. aeruginosa isolates were obtained as part of the diagnostic procedure by culturing either the corneal buttons or scrapings and identified based on growth characteristics, Gram staining, standard biochemical tests, and 16s rRNA sequencing. 8 Nineteen isolates were selected based on clinical outcomes and antibiotic sensitivity pattern. The study was approved by the Institutional Review Board of Aravind Medical Research Foundation. P. aeruginosa (ATCC® 27853™), PAO1 (kindly gifted by Dr. Eric Pearlman, University of California, Irvine, USA), and PA14 (kindly gifted by Dr. K. Balamurugan, Alagappa University, Karaikudi, India) were used as the reference strains.
Moxifloxacin,
Growth curve
To study the growth kinetics, overnight cultures adjusted to 0.5 McFarland standards were added to a 96 well plate and the growth was monitored at 600 nm using Spectramax M3 Multimode Microplate reader (Molecular Devices, San Jose, CA). ATCC 27853 was used as the reference strain and uninoculated broth was maintained as a negative control. For each isolate, the time taken to reach an optical density (OD) of 0.6 (Δt) was noted and a growth curve was generated.
Antimicrobial susceptibility testing
Minimum inhibitory concentration (MIC) was determined using broth dilution method 9 and VITEK 2. 10 In brief, in broth dilution method, serially diluted antibiotics were inoculated with test organisms in cation-adjusted Mueller–Hinton broth 9 and incubated for 18–24 hr at 37°C with appropriate controls. For VITEK 2, overnight cultures in blood agar (5% sheep blood mixed with molten nutrient agar) 8 were resuspended in 0.45% NaCl and adjusted to 0.5 McFarland standard 9 to fill the card (AST-N281, BioMerieux, Marcy-l'Étoile, France) according to manufacturer's instructions. The turbidity signal was automatically measured by the system up to 18 hr.
MDR was defined as the resistance to at least one agent in ≥3 different antibiotic classes. 11 The multiple antibiotic resistance (MAR) index was calculated using the formula, MAR index = a/b, where a is the number of antibiotics to which the isolate showed resistance and b is total number of antibiotics it has been tested against. 12 MAR index of ≤0.2 and >0.2 indicates “low-risk” and “high-risk” source of contamination, respectively.
Genotypic detection of T3SS genes
The T3SS genotype of the P. aeruginosa isolates was determined by PCR with primers specific for exoS, exoT, and exoU genes. The primers and reaction conditions are reported earlier 7 (Supplementary Table S1).
Real-time accumulation and efflux assays
The intrinsic accumulation and efflux activities of the bacterial isolates were determined under defined conditions by a semiautomated fluorimetric assay using the common efflux pump substrate ethidium bromide (EtBr). 13 For accumulation assay, bacteria grown till mid-log phase (OD600 of ∼0.6) were adjusted to an OD600 of 0.3 using 1× phosphate-buffered saline (PBS). CCCP (15 μg/mL), a proton conductor that has been shown to inhibit active efflux, was added to achieve maximum accumulation. 14 After a baseline measurement, EtBr (2 μg/mL) was added and fluorescence was measured in the multimode reader for 1 hr at 25°C (excitation, 518 nm; emission, 605 nm). Control wells had EtBr and CCCP in PBS to determine background fluorescence.
Efflux was measured as a decrease in fluorescence over time under optimal conditions. After 1 hr accumulation, the cells were harvested and resuspended in 1× PBS along with 0.4% (V/V) glucose to achieve maximum efflux. The fluorescence was again measured for 1 hr at 37°C and the normalized fluorescence was plotted against time in a graph.
MIC reversal assay
Overexpression of efflux pumps was detected by a CCCP-based microplate assay for the MDR and non-MDR strains. 15 Overnight cultures of P. aeruginosa (0.5 McFarland) were added to a mixture of serially diluted antibiotics and CCCP (25 μg/mL) in a microtiter plate. The MIC of CCCP was determined using microbroth dilution method. Totally 100 μg/mL was the MIC for ATCC strain as well as the lowest and most common MIC of our isolates. Hence we used one-fourth of that concentration (25 μg/mL) for the MIC reversal assay. Untreated bacterial suspension served as a positive control, whereas uninoculated broth with antibiotics and CCCP served as negative control. The plate was then incubated at 37°C overnight.
At least fourfold decrease in MIC in the presence of EPI indicates an overexpression of efflux pumps. 16 Efflux-mediated resistance can be confirmed when the addition of CCCP resensitizes the bacteria to the tested drug. To determine whether drug resensitization could be achieved by the addition of a membrane permeabilizer, EDTA 17 was used instead of CCCP, and MIC was recorded in the presence and absence of 0.1 and 0.2 mM EDTA. 15 Uninoculated broth containing EDTA and antibiotics served as negative control. Resensitization of the bacteria to an antibiotic in the presence of EDTA indicates poor membrane permeability as a major mechanism of resistance.
Biofilm formation assay
The ability of P. aeruginosa isolates to form biofilms was determined on microtiter plates by a crystal violet-based assay. 18 Uninoculated broth was maintained as negative control. Based on the density (absorbance550), the biofilm formation was considered to be “strong” if the absorbance was >4× absorbance of the negative control, “moderate” if the absorbance was >2× but <4 × of the negative control and “weak” if <2× but greater than the absorbance of the negative control. If the absorbance is lesser than the absorbance of the negative control, the isolate is considered to be a nonbiofilm producer.
Quantitative real-time PCR
Relative expression of efflux pump genes (mexA, mexB, oprM, mexX, and mexY) was determined by real-time qPCR. In brief, DNase-treated total RNA was reverse transcribed into cDNA using high-capacity cDNA reverse transcription kit (Applied Biosystems, Foster City, CA). Real-time PCR was performed using KAPA SYBR fast qPCR kit (KapaBiosystems, Woburn, MA) in a 7900 HT fast real-time PCR system (Applied Biosystems). The primer sequences are given in Supplementary Table S1. 19 The assays were performed in triplicates and the results were normalized using rplU mRNA levels. The relative expression between PAO1 control and test isolates was calculated by the 2−ΔΔCt method.
PCR amplification and sequencing
The acquired aminoglycoside resistance genes were detected by PCR. Mutational alteration of fluoroquinolone target genes and bacterial 16s rRNA sequences was determined by dye-terminator DNA sequencing. Multilocus sequence typing (MLST) of the MDR isolates (seven housekeeping genes acsA, aroE, guaA, mutL, nuoD, ppsA, and trpE) was performed as described previously in the pubMLST scheme (https://pubmlst.org/paeruginosa/). 20
Bacterial DNA was isolated with QIAamp DNA Mini Kit (QIAGEN, Hilden, Germany) and PCR was carried out in a total volume of 25 μL. The primer sequences are given in Supplementary Table S1.21–23 The reaction conditions included 30 sec denaturation at 95°C, 30 sec annealing at the given temperature (Supplementary Table S1), and 1 min extension at 72°C, followed by a final extension at 72°C for 5 min. The amplicons were further purified using the ExoSAP-IT PCR Product Cleanup kit (Affymetrix, Santa Clara, CA) and cyclic PCR was performed with specific primers. Big dye terminator sequencing was performed using 3130 Genetic Analyzer (Applied Biosystems).
Statistical analysis
Statistical analysis was done using GraphPad Prism software (version 7.00). Differences between two data sets were analyzed by two-tailed unpaired Student's t-test and a p-value <0.05 was considered statistically significant. All values shown are mean ± SEM values of three independent experiments, unless otherwise stated.
Results
Antibiotic susceptibility and growth kinetics
Antibiotic susceptibility testing (Table 1) was done based on the common clinical use of the tested antibiotics in bacterial keratitis.6,24 Eight isolates were MDR among which three were resistant to all the antibiotics tested except colistin, whereas the other five were resistant to at least three classes of antibiotics. In the non-MDR group (n = 11), five isolates were sensitive to all the tested antibiotics, whereas the other six showed resistance to maximum two classes of antibiotics. Since our focus is on fluoroquinolone and aminoglycoside resistance, isolates PA9 and PA10 that showed resistance to moxifloxacin and tobramycin were considered as non-MDR strains for further analysis. The other non-MDR strains were grouped along with sensitive isolates.
Minimum Inhibitory Concentration
MIC was determined using broth dilution method and VITEK 2. The values are represented in μg/mL.
R, resistant; I, intermediate; S, sensitive; MAR, multiple antibiotic resistance; MIC, minimum inhibitory concentration.
MAR index values of MDR isolates varied from 0.6 to 0.93, suggesting significant pre-exposure to antibiotics (Table 1). Higher MAR index values indicate the origin of isolates from a source where antibiotics are frequently used, as in the hospital environment. 12
The MDR isolates generally had a slower growth pattern and, therefore, higher ΔT values (mean ± SEM = 12.06 ± 1.4) than sensitive isolates (mean ± SEM = 9.55 ± 0.44) (Fig. 1 and Table 2). Notably, bacteria obtained from healed ulcers were also relatively slow growing (mean ± SEM = 11.78 ± 1.26) compared with isolates obtained from patients who had undergone therapeutic penetrating keratoplasty (mean ± SEM = 9.72 ± 0.54). However, these differences were not statistically significant owing to the small sample size and high standard deviations. Furthermore, most MDR isolates lacked the exoS gene of T3SS and had an exoUT genotype, whereas the sensitive isolates were predominantly exoS positive (Table 2).

Growth kinetics of Pseudomonas aeruginosa isolates was monitored by measuring absorbance of bacterial suspension (OD600) for 24 hr. ATCC 27853 was used as the reference strain.
Phenotypic and Genotypic Characteristics of Pseudomonas aeruginosa Isolates
TPK, therapeutic penetrating keratoplasty; UT denotes the presence of exoU and exoT genes; T denotes the presence of exoT gene; ST represents the presence of exoS and exoT genes; UST denotes the presence of exoU, exoS, and exoT genes; N represents the absence of all three genes.
Active efflux of antibiotics
Constitutive or inducible expression of the efflux systems may confer resistance to a broad spectrum of antimicrobials. 25 An EtBr-based fluorimetric assay was used to study the accumulation and efflux capabilities of bacterial isolates.13,26 EtBr emits weak fluorescence when outside the cell in aqueous medium, whereas within the cell it accumulates in the periplasm and fluoresces strongly. 13 The kinetics of EtBr transport will reflect the overall efflux capacities of the bacterial isolates, allowing bulk measurements in a single experiment.
Under minimum efflux conditions, EtBr uptake occurred rapidly during the first 5 min after which it increased gradually to reach a plateau at 1 hr. Maximum accumulation was seen in sensitive isolates PA16 and PA19 (Fig. 2a) and MDR isolates PA3 and PA6 (Fig. 2b). After accumulation, EtBr efflux was monitored at different time intervals and was found to be highest in PA10, followed by PA9 and PA5. The other isolates had low to moderate efflux activities and the results were comparable between sensitive and resistant isolates (Fig. 2c, d).
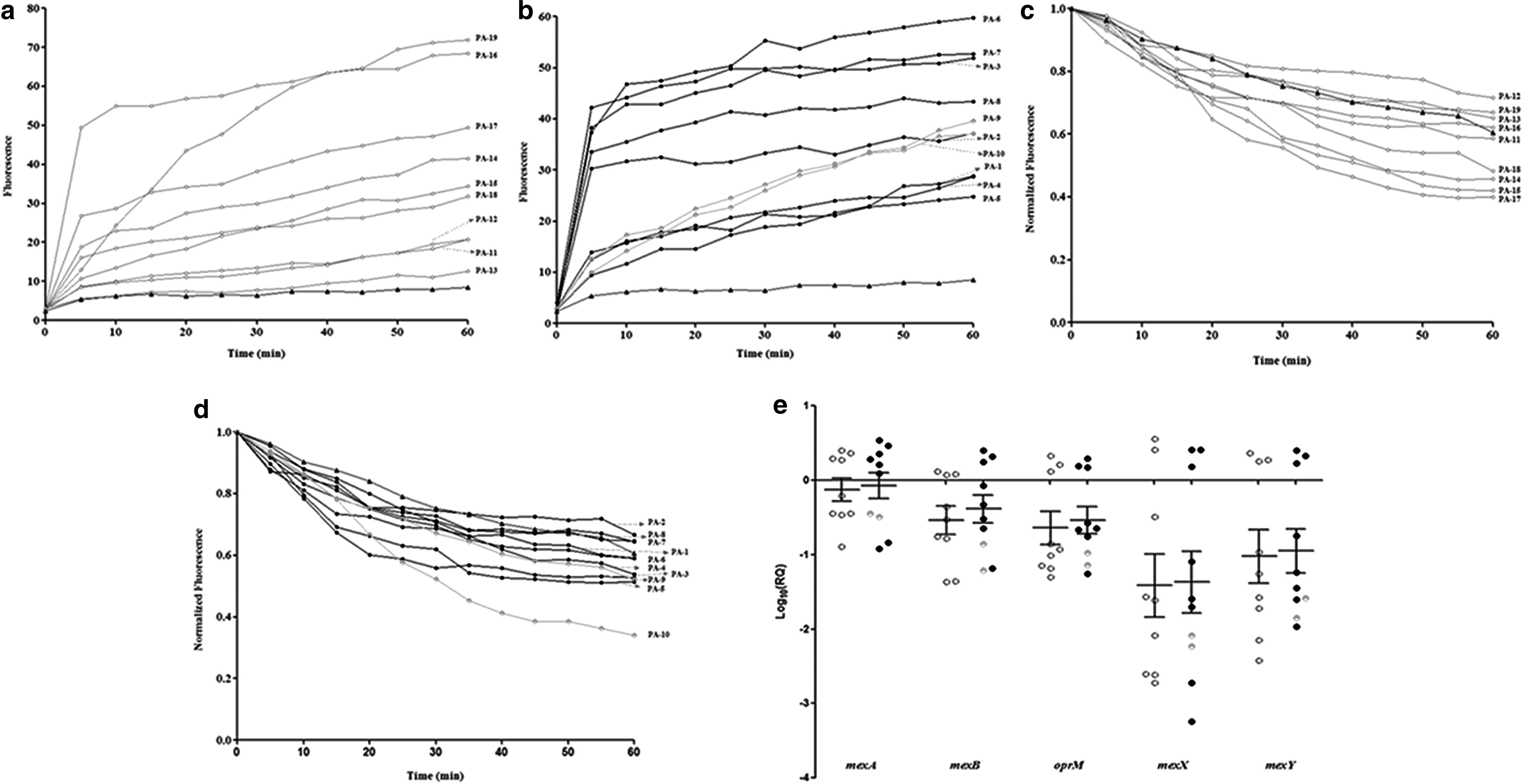
Accumulation and efflux activities of P. aeruginosa isolates. Fluorescence produced by the accumulation of EtBr (2 μg/mL) was measured in the presence of CCCP (15 μg/mL) at 25°C for 60 min in
To check whether inhibiting active efflux would resensitize the bacteria, MIC reversal assay was performed in the presence of CCCP (Table 3). Isolate PA9, which was initially resistant to moxifloxacin, regained its sensitivity, whereas PA3 and PA4 showed four- to eightfold reduced moxifloxacin MIC in the presence of CCCP. The MIC of moxifloxacin toward all other MDR isolates remained largely unchanged, indicating normal efflux activities in these cases. Similarly, PA10 that initially showed resistance only to tobramycin was also resensitized in the presence of CCCP. Except PA4, the MIC of tobramycin against all other MDR isolates reduced by two- to fourfold upon treatment with CCCP.
Minimum Inhibitory Concentration Reversal Assay
MIC was determined in the presence and absence of CCCP (25 μg/mL) and EDTA separately.
ND, not done; S, sensitive; CCCP, carbonyl cyanide
The relative expression levels of mexAB-oprM and mexXY genes were highly downregulated in most of the isolates when compared with PAO1. MDR isolates PA1, PA2, and PA6 and sensitive isolates PA11 and PA12 had increased transcript levels of all five efflux genes, whereas PA5 (MDR) had highest expression of mexA gene (Fig. 2e). However, the differences among the groups were not statistically significant.
Poor membrane permeability
To determine the role of membrane permeability in resistance, MIC reversal assay was done in the presence of a membrane permeabilizer, EDTA (Table 3). Similar to the CCCP-based assay, isolates PA9 and PA10 that were initially resistant to moxifloxacin and tobramycin, respectively, regained their sensitivity in the presence of EDTA. Three MDR isolates showed ≥8-fold decrease in MIC to both moxifloxacin and tobramycin at 0.2 mM concentration of EDTA. However, none of the MDR isolates regained their drug susceptibility, suggesting that decreased permeability may act as an additional mechanism of resistance in these organisms.
Target site alterations
We sequenced the gyrA and parC genes of all isolates, and found single amino acid substitutions of Ile for Thr-83 in gyrA and Leu for Ser-87 in parC in seven MDR isolates. PA1, the only fluoroquinolone-sensitive MDR isolate, did not possess these mutations. In addition, one more substitution of Thr for Pro-752 in parC was present in six MDR isolates including PA1 (Table 4). The mutations were not present in the sensitive and non-MDR isolates.
Target Site Alterations and Multilocus Sequence Types in Multidrug Resistant Isolates
Amino acid changes caused by mutations in gyrA and parC genes are represented with PAO1 as the reference strain. ND, not determined due to sequencing errors.
Aminoglycoside resistance in P. aeruginosa can occur due to active efflux, decreased permeability, or through acquisition of resistance genes. 5 Two MDR isolates were found to possess the rmtB gene that code for 16S rRNA methyltransferase protein (Table 2). These results suggest that 16s rRNA methylases play a minimal role in mediating aminoglycoside resistance in P. aeruginosa isolates.
Enzymatic inactivation of aminoglycosides
The isolates were screened for the presence of common AME genes, namely aph(3′)-IIb, aph(3″)-I, aph(6)-I, and aac(3)-Id. The aph(3′)-IIb gene was more prevalent and was found in both resistant and sensitive isolates (n = 13). All MDR isolates possessed aph(3″)-I and aph(6)-I genes, whereas four of them had aac(3)-Id gene (Table 2). The non-MDR strain PA9 that showed resistance to gentamicin possessed three (except aph(3′)-IIb) AME genes.
Biofilm formation
The isolates were tested for their ability to form biofilms by a crystal violet-based biofilm formation assay. Density-based grading revealed that stronger biofilms were formed by four MDR, two non-MDR, and one sensitive isolate. Moderate biofilm densities were noted with one MDR and seven sensitive isolates, whereas the others formed weak biofilms (Table 2). There was no correlation between the extent of biofilm formation and clinical outcome. Although stronger biofilms are known to be associated with increased drug resistance, the MDR phenotype was also exhibited by a couple of weak biofilm-forming organisms that had caused worst clinical outcomes (Table 2) in patients. 27
Phylogenetic analysis and MLST typing
The evolutionary and genetic relationship among the 19 isolates was studied by constructing a phylogenetic tree with the 16s rRNA sequences using MEGA7 software. 28 The isolates were grouped into two different clusters based on their sequence homologies (Supplementary Fig. S1). Both the clusters had resistant and sensitive isolates, with seven MDR isolates in the same cluster indicating genetic relatedness. Further MLST typing of the MDR isolates revealed presence of sequence types ST357, ST308, ST2579, ST3083, and ST2607 among our isolates (Table 4).
Discussion
Corneal ulcers cause severe visual disability and P. aeruginosa is the leading cause of Gram-negative bacterial corneal ulcers among our patients. 29 Continuous exposure of pathogenic organisms such as P. aeruginosa to antimicrobials and heavy metals in the environment leads to the selection of MDR organisms that pose a global threat. 30 In this study, we have investigated the underlying mechanisms of resistance in P. aeruginosa isolates with varying susceptibility profiles that were obtained from corneal ulcer patients. Fluoroquinolones such as moxifloxacin and aminoglycosides such as tobramycin are commonly used to treat P. aeruginosa corneal infections. 31 Hence we sought to identify the mechanisms behind quinolone and aminoglycoside resistance in a select group of clinical isolates.
At the outset, the MDR strains had slower growth rates that might imply that acquisition of resistance genes had happened at the cost of fitness to the bacterium. The incompetent bacterial replication could be a reason that some of these ulcers could heal despite the reduced antibiotic susceptibility of the isolates. Thus, the acquisition of a MDR phenotype by the bacterium could even prove beneficial to the host by reducing bacterial viability. One such typical example was the MDR strain PA5 that possessed all the resistance determinants, yet cleared easily (healed ulcer) perhaps due to its slower growth pattern (ΔT, 14 hr) (Table 2).
The MDR isolates were also predominantly exoU positive, and such an association between multidrug resistance and T3SS genotype has previously been reported from our group with endophthalmitis cases, where all the MDR isolates were found to possess the exoU gene. 7 Another report from the SCUT has shown elevated fluoroquinolone resistance in cytotoxic exoU encoding P. aeruginosa strains and suggested association with worse clinical outcomes. 6
Being an opportunistic pathogen, P. aeruginosa has developed selective outer membrane permeability and an array of efflux systems that maintain cell homeostasis during hostile conditions. 26 Among the various efflux systems present in P. aeruginosa, the clinically relevant resistance-nodulation-division (RND) efflux systems have broad substrate specificity and mediate active efflux of various antibiotics such as β-lactams, chloramphenicol, trimethoprim, fluoroquinolones, tetracyclines, and aminoglycosides.32,33 MAR in P. aeruginosa has been shown to be associated with overexpression of RND efflux pumps along with target site mutations. 34 MexAB-OprM dysregulation alone has been reported to decrease the antibiotic susceptibility of clinical isolates. 35
In PAO1, expression of MexXY has been shown to be induced by tetracycline, erythromycin, or gentamicin. 36 It functionally associates with OprM and contributes to intrinsic resistance to these antimicrobials. In this study, the RND efflux system genes were upregulated at the transcript level only in few isolates. Notably, the expression of mexX and mexY was highly downregulated in most of the isolates, which could be due to the absence of an inducing agent in culture. Even in isolates with an increased expression of mexAB-oprM- and mexXY genes, active efflux does not seem to play a major role in conferring drug resistance, as suggested by the MIC reversal data. However, the non-MDR strains were readily resensitized in the presence of CCCP, indicating active efflux as the predominant mechanism of resistance in these cases.
It is quite possible that increased efflux is the initial step in the development of drug resistance that is later taken over by other more powerful mechanisms such as target site mutations or acquisition of resistance genes. Increased efflux activity was also noted in few sensitive isolates, but the mechanism that protects them against developing resistance is unclear. Similar to efflux inhibition, enhancing membrane permeability with EDTA also resensitized only the non-MDR isolates that had lower MIC levels. In these isolates, active efflux along with reduced permeability may prevent drug accumulation inside the cells. Increasing the membrane permeability of these strains could reverse efflux-mediated resistance and vice versa. The existence of synergy between these two resistance mechanisms has already been noted in Gram-negative bacteria. 37
Fluoroquinolones primarily target DNA gyrase and topoisomerase IV, bacterial enzymes that are involved in DNA replication, transcription, and recombination. 38 ParC is a subunit of topoisomerase IV, and the quinolone resistance determining regions (QRDRs) of parC and gyrA have high sequence similarities. 39 Mutations in DNA gyrase lead to quinolone resistance, whereas parC mutants increase resistance levels by acting as a prerequisite for gyrA mutations. 40
The predominant amino acid substitutions noted among our MDR isolates, Thr-83→Ile in GyrA and Ser-87→Leu in ParC, have previously been reported in ciprofloxacin-resistant P. aeruginosa isolates and an association was found between ciprofloxacin MIC and the number of target site alterations.40,41 In Gram-negative bacteria, mutations first arise in the most susceptible DNA gyrase gene, followed by additional mutations that further increase resistance. 32
Apart from QRDR, the 16S rRNA subunit is also subject to alterations such as methylation that confers panaminoglycoside resistance. 42 The limited frequency of rmtB gene among our isolates is consistent with an earlier report, where only one strain of VIM-4 MBL-producing P. aeruginosa isolated from the burn patients possessed this gene. 43 In contrast, enzymes that catalyze modification of aminoglycosides are more prevalent among Gram-negative bacteria. AME could be acetyltransferases (AACs), phosphotransferases (APHs), or nucleotidyltranferases (ANTs) that modify the hydroxyl or amino groups of the 2-deoxystreptamine nucleus or the sugar moieties. 44
In this study, aph(3″)-I and aph(6)-I genes were detected in all our MDR isolates and the presence of AME genes was associated with widespread resistance to gentamicin, tobramycin, and amikacin. Altogether, our data imply that aminoglycoside resistance among the ocular P. aeruginosa isolates is largely mediated through the enzymatic modification of drugs.
Finally, we tested the biofilm-forming ability of our isolates, since these surface-attached bacterial communities show increased resistance to antimicrobials, which could be 1,000-fold more than in planktonic cells. 45 Although our resistant isolates had better biofilm-forming abilities, two MDR isolates that were associated with worst clinical outcomes formed only weaker biofilms. Hence biofilm formation may not be an important cause of drug resistance in ocular infections. However, we cannot rule out the possibility that by creating a physical barrier to antimicrobials, biofilms could have facilitated development of drug resistance by decreasing the clinical efficiency of antibiotics. Recent studies have shown that exposure of bacteria to intermittent concentrations of antimicrobials and formation of persisters often precede development of drug resistance. 46
To conclude, we have found the mutational alteration of QRDR and the presence of AME genes as the most common and predominant mechanisms of quinolone and aminoglycoside resistance in corneal ulcer isolates of P. aeruginosa. Mutations are selected under continuous antibiotic pressure and passed onto subsequent generations as an adaptive strategy for survival. Dysregulations in membrane permeability and efflux also develop as a result of previous antibiotic exposure and confer resistance during subsequent challenges with the drug.
The smaller sample size of this study has undermined the significance of certain findings. Although we aimed for a much larger sample size, the poor growth pattern of many resistant isolates made them unsuitable for a complete analysis. Still, the study throws light on the antibiotic resistance pattern and the associated mechanisms of resistance in a select group of clinical isolates. In the long run, pertinent information on various drug–pathogen interactions will aid in the development of adjunctive therapies or other improvised methods to prevent the spread of multidrug resistance.
Footnotes
Acknowledgments
We thank Sangeetha Ramalingam, Kathirvel Kandasamy, and Rameshkumar Gunasekaran for their assistance in this work. The support provided by Aurolab, Madurai, in procuring the antibiotics is gratefully acknowledged. This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Disclosure Statement
All of the authors declare that they have no competing interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.