
Abstract
NetF-producing type A Clostridium perfringens, a pathotype of C. perfringens, causes necrotizing enteritis in neonatal foals and necrotizing and hemorrhagic enteritis in dogs. Recent core genome multilocus sequence typing study revealed that netF+ C. perfringens strains belong to two distinct clonal populations (clonal complexes I and II). There are no reports on susceptibility to antimicrobial drugs of isolates from this pathotype. The susceptibility to 13 different antimicrobial drugs of 49 netF+ strains recovered from foals or dogs with necrotizing enteritis in Canada, the United States, and Switzerland was assessed using a commercial microdilution panel designed for anaerobic human pathogens. All isolates were highly susceptible to 12 antimicrobial agents, including all beta-lactams tested, such as penicillin G and ampicillin, as well as clindamycin, chloramphenicol, and metronidazole. The isolates consistently presented a reduced susceptibility or resistance to tetracycline, which was associated with previously described tetracycline resistance genes. Clonal complex I isolates (n = 41) possessed the tetA408(P) gene, whereas clonal complex II isolates (n = 8) possessed the tetA(P)-tetB(P) genes and were more likely to be fully resistant.
Introduction
C
NetF, a pore-forming toxin within the Leukocidin/Hemolysin superfamily, is preferentially produced by C. perfringens strains from cases of acute canine hemorrhagic and necrotizing enteritis and from necrotizing enteritis in neonatal foals, some of which are fatal. 3 NetF+ C. perfringens strains belong to two distinct clonal populations (clonal complexes I and II). 4 Disease in dogs or foals can be associated with either clonal population.
Nothing is known of the antimicrobial susceptibility of netF+ C. perfringens, although whole genome analysis of 32 netF+ strains showed that they harbor tetracycline resistance determinants.4,5 The aim of this study was, therefore, to assess the in vitro antimicrobial susceptibility of netF+ strains isolated from dogs and foals with necrotizing enteritis. In addition, the genetic organization of tetracycline resistance genes in each clonal population was investigated.
Materials and Methods
Bacterial isolates
A total of 49 netF+ C. perfringens isolates from our culture collection (Department of Pathobiology, University of Guelph) were analyzed in this study. These isolates were from unrelated individual clinical cases recovered from foals with necrotizing enteritis (n = 29) or dogs with acute hemorrhagic enteritis (n = 20) between 1999 and 2018 in Canada (foal, n = 7; dog, n = 10), the United States (foal, n = 22; dog, n = 7), and Switzerland (dog, n = 3).
DNA extraction
To isolate genomic DNA, three to five single bacterial colonies from overnight blood agar plates culture were resuspended in 500 μL of autoclaved water and then centrifuged at 12,000 g for 1 min. The supernatant was removed and DNA of the samples was extracted using InstaGene matrix (Bio-Rad Laboratories, Mississauga, ON, Canada). In brief, 200 μL of InstaGene matrix was mixed with each bacterial cell pellet and incubated at 56°C for 25 min, following with additional incubation in boiling water for 8 min. The samples were then centrifuged at 12,000 g for 3 min and the supernatants were kept at −20°C until use.
Clonal relationship analysis and detection of tetracycline resistance genes by PCR
Whole genome sequencing has shown that netF+ C. perfringens may harbor either the tetA(P) and tetB(P) genes or the tetA408(P) gene. NetF+ C. perfringens strains were examined by PCR for the presence of the tetA(P) and tetB(P) genes (overlapping PCR product) and for the tetA408(P) gene. PCR amplification was carried out in a 25 μL total volume reaction containing 1 × PCR buffer with Mg2+ (New England BioLabs, Pickering, ON, Canada), 0.2 mM dNTP mixture, 1.25 units of TaqDNA polymerase (New England BioLabs), 10 pmol of each primer, and 5.0 μL of each DNA template. The PCR program for tetA(P)-tetB(P) and tetA408(P) was 94°C for 3 min, 29 cycles of 94°C for 30 sec, 45°C for 30 sec, extension at 72°C for 1 min, and finally, 72°C for 7 min.
To determine whether the netF+ isolates tested in this study belong to clonal complex I or clonal complex II, unique primers were designed for each clonal population based on previous genome sequencing of netF+ strains.4,5 The PCR amplification and programs for clonal complex identification were similar to those already described with the exception of the annealing temperature, 53°C for 30 sec/cycle. The amplified PCR products were separated by 1.5% agarose gel electrophoresis and observed by Ez-Vision Two DNA dye marker (AMRESCO, Inc., Solon, OH) and photographed under ultraviolet light by GelDoc (Bio-Rad Laboratories). The following primers were used in this study:
tet(A/B)-F: GCTACTGTACTTTCTATAAATG tet(A/B)-R: ATGTGTCAAATAATATTCTTGT (predicted product size = 550 bp)
6
tetA408-F: CATGATATTTTTTTATCCATTATC tetA408-R: GCCAGAAAATAATTTTAAACC (predicted product size = 685 bp) Clade I-F: ACCGTTGAAGATCAACTTGGGA Clade I-R: TTGCCCTCTCTTCAGAACCTG (predicted product size = 397 bp) Clade II-F: ATTTGTTCTGGGTGTGGTGC Clade II-R: GCTGCAACCTGACAAGGTGT (predicted product size = 476 bp)
Antibiotic susceptibility testing
Minimal inhibitory concentrations (MICs) were determined by microdilution broth method using the commercially available Sensititre plate ANO2B (TREK Diagnostic Systems, Cleveland, OH). The plate, which is designed for human anaerobic pathogens, consists of 15 antibiotics and is based on the guideline M11 of the Clinical and Laboratory Standards Institute (CLSI, Wayne, PA). The plate included two carbapenems, imipenem and meropenem, which are not only rarely used in veterinary medicine but also should be avoided. 7 Therefore, these antibiotics are not included in this study. The susceptibility test was performed according to the manufacturer's instructions. The microdilution plates were incubated for 24 hr at 37°C under anaerobic conditions and test results were read using a Sensititre manual viewer (TREK Diagnostic Systems). C. perfringens ATCC 13124 and Bacteroides fragilis ATCC 25285 were used as quality controls for each test run. Breakpoints were determined using the M100-S28 guideline of CLSI-2018 8 for anaerobic bacteria.
Results
The MIC distributions and the MIC50 and MIC90 of 13 antibiotics tested against 49 netF+ C. perfringens strains are presented in Table 1. With the exception of tetracycline, all isolates showed susceptibility to the antimicrobial tested. For tetracycline, 4 (8%) isolates were resistant (MIC ≥16 μg/mL) and 23 (47%) strains displayed reduced susceptibility (MIC only a twofold dilution below the defined breakpoint) (Table 1).
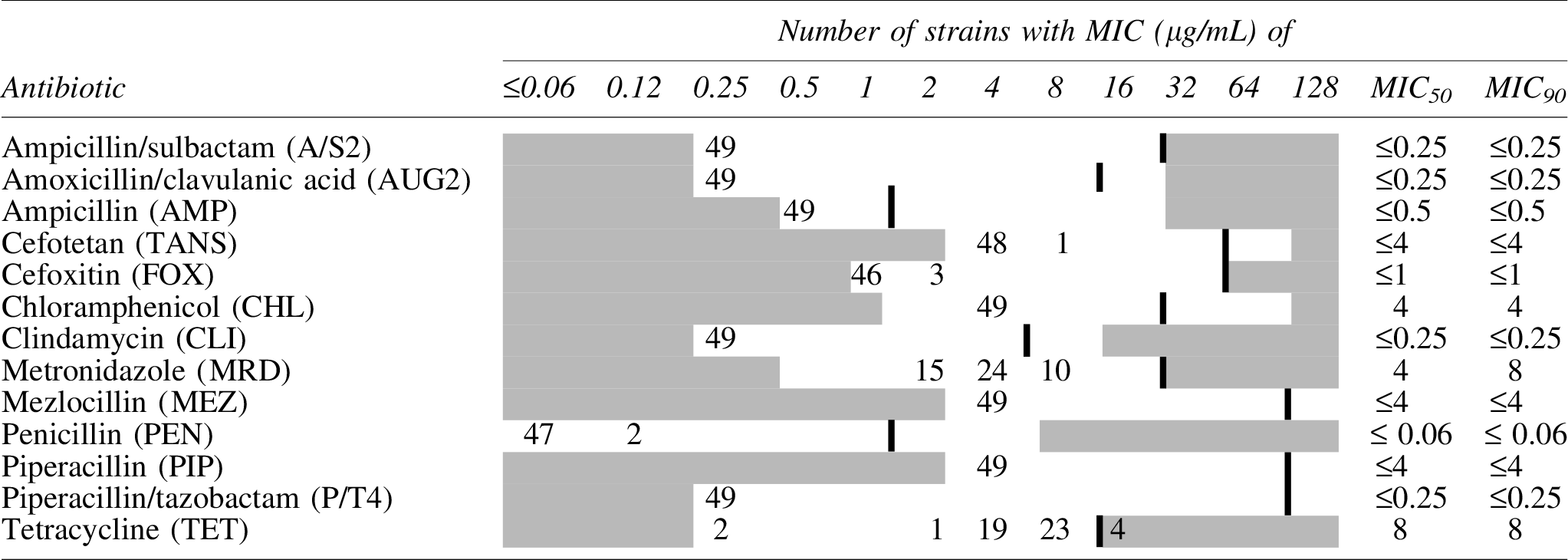
Areas in white represent the concentration ranges tested for each antimicrobial agent. Areas in gray denote concentrations outside of the dilution range of the antimicrobial. Bolded vertical bar lines indicate the clinical “breakpoint” for resistance, Clinical and Laboratory Standards Institute, 2018.
MIC, minimal inhibitory concentration.
PCR analysis for clonal population identification showed that the majority of isolates (foal, n = 22; dog, n = 19) belonged to clonal complex I, with the remaining isolates (foal, n = 7; dog, n = 1) being clonal complex II.
The genetic organization and distribution of tetracycline determinants in netF+ strains were evaluated by PCR (Table 2). With the exception of two isolates (one canine clonal complex I isolate, strain number JFP718 [GenBank Accession No. MARD00000000] and one equine clonal complex II isolate, strain number JFP55 [GenBank Accession No. CP010993]), all isolates harbored either the tetA408(P) or both the tetA(P) and tetB(P) genes. The two isolates without tetracycline resistance genes were highly susceptible to tetracycline with an MIC of ≤0.25 μg/mL (Table 1). Interestingly, all other members of clonal complex I carried the tetA408(P) gene, whereas the tetA(P) and tetB(P) genes were only found in clonal complex II isolates (Table 2). No apparent correlation between the source of isolates and genetic organization of tetracycline genes was detected. The tetA408(P) gene was located chromosomally, based on the 26 isolates for which genome sequencing data were available,4,5 and the tetA(P) and tetB(P) genes were located on a plasmid, based on the 6 isolates for which genome sequencing data were available.4,5 Contig data were insufficient to determine whether the plasmid(s) was or was not tcp based. 9
These isolates lacked the resistance gene.
Discussion
This study provides for the first time the antibiotic susceptibility profile of netF+ C. perfringens strains derived from foals and dogs with necrotizing enteritis. The results of this study show a high degree of susceptibility to antimicrobial agents commonly used against anaerobic pathogens, with the exception of tetracycline. Beta-lactams, macrolides, metronidazole, and tetracycline are the most commonly recommended antibiotics for the treatment of C. perfringens infections in animals, 7 including severe diarrheal illness in dogs. 10
The uniform susceptibility to beta-lactam antibiotics is in agreement with previously published studies of C. perfringens isolated from numerous different animal species,11–18 although reduced susceptibility or resistance has been occasionally reported.19,20
Metronidazole has been described, like penicillin, as a first choice option for treatment of clostridial diarrhea in dogs and horses.10,21 In this study, no resistance to this drug was observed (MIC90 ≤ 8 μg/mL). Decreased susceptibility of C. perfringens to metronidazole has been described,14,22 including in canine isolates, 15 although full resistance is rare in clinical isolates from humans and animals. Álvarez-Pérez et al. found only 4 of 80 isolates (5%) from various animal species resistant (MIC ≥32 μg/mL) to metronidazole. 23 Similarly, only 1 of 131 canine C. perfringens was deemed resistant to metronidazole. 24 All isolates were susceptible to chloramphenicol and clindamycin.
An interesting finding of this study was the consistent presence of reduced susceptibility or resistance to tetracycline (Table 1) associated with the presence of tetracycline resistance genes (Table 2). The identity of these genes was directly related to the clonal complex to which the isolates belonged (Table 2). High MICs to this antibiotic have been frequently reported in C. perfringens strains from different animal origins.12,13,24,25 It has been shown that C. perfringens isolates resistant to tetracycline carry one of the following three combinations of tet genes: (1) tetA(P) and tetB(P), (2) tetA408(P) and tet(M), and (3) tetA408(P). 6 These genes are commonly found on plasmids or transposons. 9 Earlier genome sequencing of 32 of netF+ isolates showed that the tetA(P)-tetB(P) and tetA408(P) genes were located on plasmid and chromosome, respectively, and no tet(M) gene was identified.4,5 Similar results were described by others.25,26 For instance, Kather et al. found that 96% (119/124) and 41% (51/124) of canine C. perfringens isolates were positive for tetA(P) and tetA(P)/tetB(P) genes, respectively, and that none possessed the tet(M) gene. In addition, only half of the tet-positive isolates in that study were phenotypically resistant to this drug. 26
The striking association of different tetracycline resistance genes with the two clonal complexes of netF+ strains isolated from different continents is further support for the divergence of these strains, which possess closely related pNetF, pCPE-CPB2, and pBacteriocin plasmids in common, during their evolution. The striking absence of resistance to clindamycin unlike isolates from farm animals,14,16 and unlike the 21% resistance described in other C. perfringens from dogs in Korea, 27 suggests that these strains have not been exposed to lincosamides or macrolides. Speculatively, the selection pressure for maintaining tetracycline resistance genes encoding reduced susceptibility or resistance to tetracycline might be to defend against environmental microbes producing this antibiotic rather than anthropogenic sources.
Footnotes
Acknowledgments
We thank Drs. Tom Besser, Robert Carman, Vincent Perreten, Scott Weese, Diego Gomez, and Ashley Whitehead for providing some of the strains used in this study. We also thank Dr. Durda Slavic for providing access to a Sensititre autoinoculator and Sensititre manual viewer. We thank the Natural Sciences and Engineering Research Council of Canada for funding of research relating to netF-C. perfringens.
Disclosure Statement
No competing financial interests exist.