
Abstract
Aims:
The emergence and clonal occurrence of multidrug-resistant (MDR) Escherichia coli isolates are increasing worldwide. In this study, phenotypic and genotypic characteristics of MDR E. coli isolates overexpressing efflux pump were investigated in medical centers of Azerbaijan. Two hundred nineteen consecutive and nonduplicated isolates of E. coli were collected and screened, and confirmed for extended-spectrum β-lactamase, AmpC, and carbapenemase activities and respective genes. MDR isolates were selected and subjected to efflux pump overexpression assay. Cefoxitin-nonsusceptible isolates were subjected to mutational analysis of promoter region of chromosomal ampC gene. MDR isolates with overexpressed efflux were analyzed for acrR and marR mutations and assigned to multilocus sequence typing.
Results:
Eighty (36.5%) isolates had MDR pattern, among which 16 (20%) isolates were positive for overexpressed efflux. Ninety-eight of 99 suspected isolates were positive for any β-lactamase genes, particularly CTX-M groups 1 and 9. Ten out of 33 cefoxitin-nonsusceptible isolates had mutations in promoter region of chromosomal AmpC gene, including −32T→A (n = 5), −42C→T, and −18G→A (n = 3) and −13TT and GT insertion (n = 2). Detected mutations in efflux regulatory genes include G103S and Y137H (n = 15), K62R (n = 8), S3N (n = 3), and A53E (n = 1) in marR and L109 (n = 2) and L190 (n = 1) frameshift mutations and T12M, T213I, N214T, I113V, and H115Y point mutations (n = 5) in acrR.
Conclusions:
Overexpressing efflux pump isolates belonging to sequence type (ST)131 and ST73 clones are emerging in Azerbaijan hospitals. Clonal occurrence of MDR E. coli is an alarming situation in Azerbaijan hospitals.
Introduction
In recent years, occurrence of multidrug-resistant (MDR) Escherichia coli, as a major public health concern, has been increasingly reported.1,2 Infections caused by these organisms are often associated with treatment failure and high rates of morbidity, mortality, and health costs. 3 MDR has been defined as nonsusceptibility to three or more different classes of antibiotics. Several acquired and/or intrinsic mechanisms are responsible for MDR phenotype among E. coli strains. It has been known that high-level MDR involves acquired mechanisms along with mutations at several structural and regulatory genes.4,5
Acquired β-lactamases, including extended-spectrum β-lactamases (ESBLs), plasmid-mediated Ambler class C cephalosporinases (pAmpCs), and carbapenemases, play an important role in the resistance to a wide range of β-lactam (through direct hydrolysis) and non-β-lactam (through simultaneous dissemination of resistance determinants on mobile genetic elements by which they are carried) drugs.6–9 As an intrinsic mechanism, it has been shown that overexpression of AcrAB-TolC, a chromosomally encoded efflux pump, plays an important role in the emergence of MDR isolates 4 and is associated with cross-resistance to β-lactams, quinolones, chloramphenicol, cyclines, 10 and even carbapenems and cephamycins (the most effective drugs for ESBL producers), if coincided with membrane permeability defects resulted from porin loss or porin structural changes.11,12
AcrAB-tolC operon encodes a multidrug efflux pump under negative regulation of AcrR repressor and also positive regulation of MarA transcriptional activator (product of MarRAB operon). 13 It has been shown that mutations at the acrR and marR (the repressor of marRAB operon) increase the expression of the acrAB operon by inactivation of AcrR repressor and overexpression of MarA transcriptional activator, respectively.13–15 This study was conducted to achieve MDR profile among clinical isolates of E. coli with respect to major intrinsic and acquired mechanisms.
Materials and Methods
Clinical isolates
Between January and July 2016, 219 consecutive and nonduplicated isolates of E. coli were collected from major medical centers in East and West Azerbaijan, Iran. All patients with at least 48 hours of hospitalization were included in this study from various clinical settings. Admitted patients with previous community-acquired infection were excluded. Only one isolate per patient was included in this study. Standard biochemical tests were used to identify bacterial isolates. Institutional Review Board approval was obtained from ethical review board of West Azerbaijan University of Medical Sciences.
Initial screening and confirmatory tests for β-lactamase activity
All isolates were screened for ESBL, AmpC, and carbapenemase production using the disc diffusion method against third-generation cephalosporins (ceftazidime, cefotaxime, cefuroxime, and ceftriaxone), carbapenems (imipenem, meropenem, and ertapenem), and cefoxitin. To identify MDR isolates, the susceptibility of isolates also was tested against ciprofloxacin, gentamicin, amikacin, cefepime, ampicillin, amoxicillin/clavulanic acid, piperacillin/tazobactam, and trimethoprim-sulfamethoxazole. All discs were purchased from Rosco diagnostica, Denmark. Chromosomal AmpC hyperproducer isolates and positive isolates for overexpressed efflux activity were also tested to determine the minimum inhibitory concentration (MIC) of ceftazidime, cefotaxime, and ciprofloxacin using E test Strips on Mueller Hinton agar (AB Biodisk, Solna, Sweden). Categorical interpretations for susceptibility patterns recommended in Clinical and Laboratory Standards Institute (CLSI) document M100 (29th Edition) were considered for selected antimicrobial agents. 16 β-Lactamase producers were selected based on the CLSI screening criteria for ESBL, carbapenemase, and AmpC activities. Suspected β-lactamase producers were further phenotypically confirmed using total ESBL/AmpC confirm kit, AmpC/ESBL confirm kit and KPC/MBL, and OXA-48 confirm kit (Rosco Diagnostica, Denmark). The results were interpreted according to manufacturer's instructions. Escherichia coli ATCC 25922 and Klebsiella pneumoniae ATCC 700603 were used as quality control strains in each set of susceptibility tests.
Detection of overexpressed efflux pump activity
MDR isolates were selected according to the MDR definition as nonsusceptibility to three or more different classes of antibiotics 17 and were subjected to efflux pump overexpression assay using previously described ethidium bromide (EtBr)-agar Cartwheel method. 18 The overexpressed efflux activity was further confirmed by determining the MIC of ciprofloxacin in the presence of efflux inhibitor, Phenylalanine-Arginine β-Naphthylamide (PAβN; Sigma Aldrich). MIC was determined using Etest stripes (AB Biodisk) in Mueller Hinton agar plates containing 20 mg/L PAβN for each isolate according to CLSI recommendations. Efflux activity was confirmed by at least 1/4 decrease of the MIC value in the presence of PAβN compared to original values.
Detection of acquired β-lactamase genes
All MDR isolates and suspected isolates for β-lactamase activity were further examined for the presence of ESBL, pAmpC, and carbapenemase genes using the following set of multiplex PCR assays: detection of blaTEM, blaSHV, blaCTX-M, blaGES, blaVEB, blaPER, blaIMP, blaVIM, and blaKPC genes was performed using previously described multiplex PCR assays by Dallenne et al., 19 including I, II, IV, and VI. Also, multiplex PCR groups A, B, C1, C2, and D1 described by Voets et al. 20 were used to detect blaSME-1–3, blaIMI-1–3, blaNMC-A, blaGIM-1, blaSIM-1, blaNDM-1, blaSPM-1, blaCMY-1,8–11,19, blaMOX-1–4,5–7, blaDHA-1–3,7, blaACC-1–4, blaCMY-2, blaLAT-1,4, blaACT-1–3, blaMIR-1–4,8, blaFOX-1–8, and blaOXA-48 genes. 20 Total DNA was extracted from one heated colony (95°C for 10 minutes), from overnight culture grown on solid media in 20 μL of tissue buffer (0.25% SDS +0.05 M NaOH) followed by a centrifugation step at 13,000g for 1 minute. Each set of amplification was done according to respective primer concentrations and conditions described by Dallenne et al. 19 PCR products were visualized after running them on a 1% agarose gel containing EtBr at 100V for 1 hour. A 100 bp DNA ladder (CinnaGen, Iran) was used as a size marker. In this study, quality control strains for each set of amplification were provided from microbiology department of Tabriz Institute for Pharmaceutical Research. Resulting PCR amplicons were purified using the QIAquick PCR purification Kit (Qiagen Company, Germany) and subjected to bidirectional sequencing using the same primers as for PCR and ABI 3730XL DNA Analyzer. Each sequence was then compared with already known β-lactamase gene sequences using the NCBI Antimicrobial Resistance Gene Finder (AMRFinder) according to the latest version of AMR gene database.
Mutational analysis of chromosomal AmpC promoter/attenuator region
All isolates with a cefoxitin inhibition zone <20 mm and/or cefuroxime MIC value >8 μg/mL were considered possible AmpC hyperproducers and were subjected to mutational analysis of promoter/attenuator region by PCR and sequencing as previously described. 21 Briefly, a 271 bp fragment of E. coli ampC gene containing the promoter, attenuator, and part of the coding region was amplified and sequenced using a set of primers. Resulted sequence of each product was aligned against the GenBank entry for Escherichia coli K12 (GenBank accession no. NC000913) and E. coli control strain ATCC 8739 (GenBank accession no. CP000946) using Clustal W online software. Jaurin et al. 22 numbering system was used to record nature and position of each mutation.
Mutational analysis of acrR and marR transcriptional repressors
To determine the possible contribution of mutations in overexpression of AcrAB efflux pump, all MDR isolates with phenotypically confirmed overexpressed efflux activity were subjected to sequence analysis of acrR and marR genes. Amplification of the acrR gene was performed using AcrR-Forward (5′-GCTGCGTTTATATTATCGTCGTGC-3′) and AcrR-Reverse (5′-GTCAAACCGCAAGAATATCACGACG-3′) primers, and previously described PCR protocol. 23 marR gene, including its operator region, marO, was amplified with MarR-Forward (5′-AGCTAGCCTTGCATCGCA-3′) and MarR-Reverse (5′-TACGGCAGGACTTTCTTAAGCA-3′) primers, and PCR protocol described by Park et al. 24 After sequencing, resulted nucleotide sequences were compared with the GenBank sequences found under accession number M96235 for mar regulon and AE000152 U00096 for Escherichia coli K12 wild-type acrR gene.
Multilocus sequence typing
All MDR isolates that were characterized for overexpressed efflux and chromosomal AmpC activities were assigned to multilocus sequence types (MLST) using the Institut Pasteur tools. PCR amplification and sequencing of seven housekeeping genes (dinB, icdA, pabB, polB, putP, trpA, and uidA) were performed using the primers and protocol specified at the website. Sequence type (ST) of each isolate was determined by comparing of sequences with corresponding alleles available in MLST E. coli database and combining seven allelic profiles.
Data analysis
The descriptive statistics, including the frequencies, case summaries, and cross tabulation of clinical, microbiological, and epidemiological characteristics, was performed using the computer software program SPSS, version 21.
Results
During January to July 2016, a total of 219 clinical isolates of E. coli (122 and 97 isolates from East and West Azerbaijan, respectively) were isolated from admitted patients in various hospital wards, including internal (n = 144, 65.7%), surgery (n = 37, 16.9%), intensive care units (n = 24, 11%), pediatric (n = 11, 5%), and burn wards (n = 3, 1.4%). The mean age of patients was 54.22 (standard deviation = 21.4) years and 64.8% were females. The majority of isolates were predominantly obtained from urine samples (n = 177, 80.8%), while other samples were recovered from blood cultures (n = 29, 13.2%), burn wound exudates (n = 3, 1.4%), wound purulent discharges (n = 3, 1.4%), tracheal and bronchial secretions (n = 3, 1.4%), abdominal fluid (n = 2, 0.9%), and sputum and cerebrospinal fluid (one isolate of each). Thirty-five patients (24.2%) were hospitalized for more than 21 days and 36 patients (16.4%) had a history of hospitalization in the last 6 months. One hundred twelve patients (51.1%) had a history of antibiotic use in the last 4 weeks. Additional patient's clinical characteristics are described in Table 1.
Patient's Clinical Characteristics
CNS, central nervous system; ICU, intensive care unit.
Initial screening and phenotypic confirmatory tests for β-lactamase activity
According to the results of initial screening, 136 of 219 isolates (62.1%) were nonsusceptible to at least one tested antimicrobial. Ninety-nine isolates were suspected for β-lactamases activities, among which 99, 33, and 4 isolates met selection criteria for ESBL, AmpC hyperproducer, and carbapenemase activities, respectively. Ninety-eight of 99 initially suspected isolates were confirmed for respective β-lactamase activities, including 75 (34.2%, overall rate) isolates for ESBL, 14 (6.4%) isolates for simultaneous ESBL and AmpC hyperproducer, and 9 (4.1%) isolates for only AmpC hyperproducer. The overall rates of ESBL and AmpC hyperproducer isolates were 27.8% (61 out of 219) and 10.5% (23 out of 219), respectively. No carbapenemase activity was detected for four carbapenem-nonsusceptible isolates. Eighty isolates (36.5%) were recognized to have MDR pattern. The results of initial screening and antimicrobial susceptibility tests are presented in Table 2.
Initial Screening for β-Lactamase Production and Antimicrobial Susceptibility Patterns of Escherichia coli Isolates
Detection of overexpressed efflux pump activity
According to the results (Fig. 1), 16 of 80 (20%) MDR isolates were recognized to have overexpressed efflux pump activity, all of which were confirmed by at least 1/4 decrease in ciprofloxacin MICs in the presence of 20 mg/L PAβN. In five isolates, which had low level MICs against ciprofloxacin, the rates of MICs were fully reverted to susceptible range (≤1 μg/mL) in the presence of efflux inhibitor, suggesting that MDR pattern observed in these isolates was solely related to overexpressed efflux pump activity. Remaining 11 isolates showed high-level ciprofloxacin MICs and displayed more than 1/4 MIC reduction toward intermediate range (2 μg/mL), indicating a partial contribution of efflux pump activity toward respective MDR pattern.
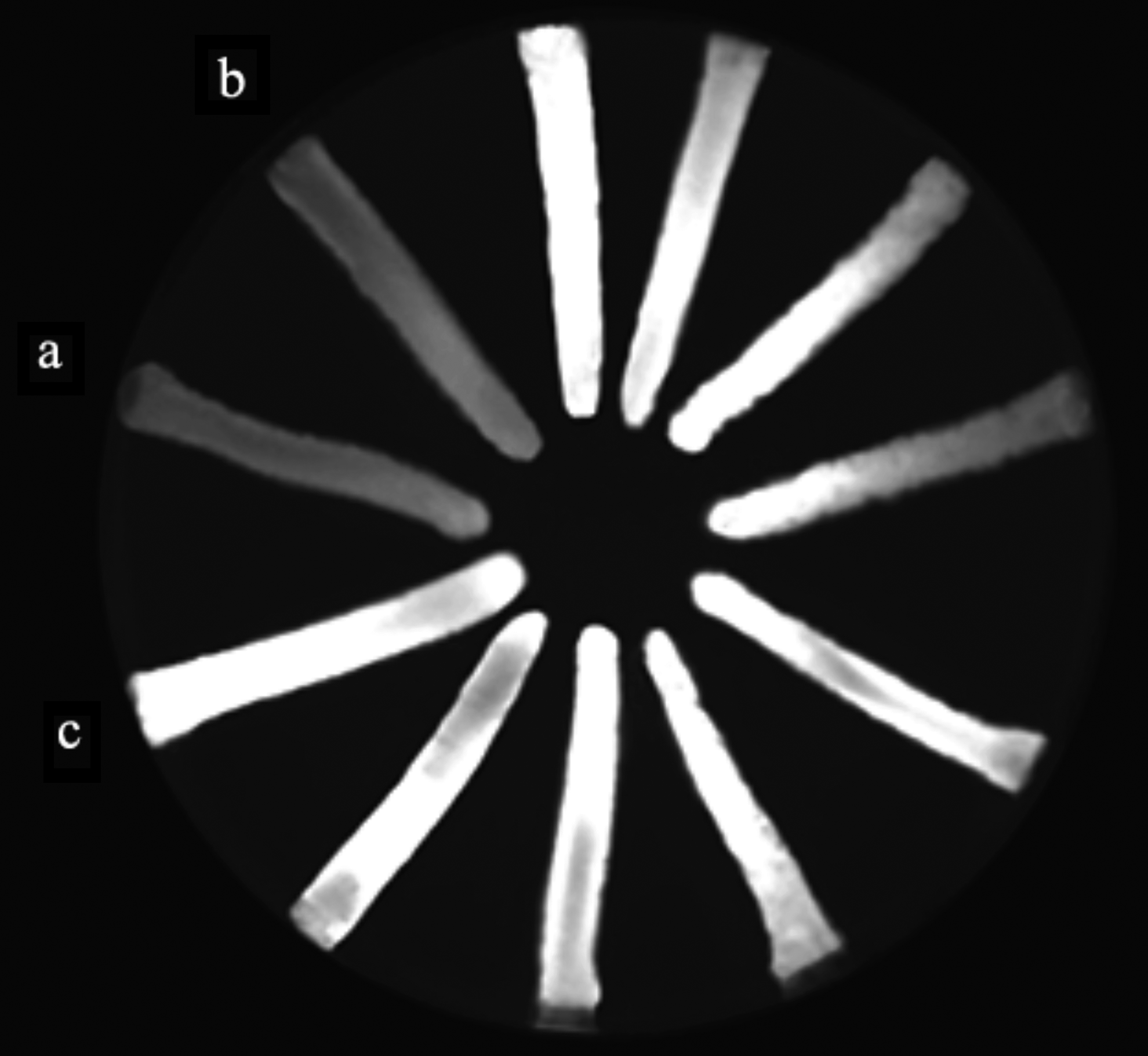
Detection of overexpressed efflux pump activity using the EtBr-agar cartwheel method. Overnight cultures of MDR isolates were swabbed in TSA plates containing 2.5 mg/L EtBr. Following overnight incubation at 37°C for 16 hours, fluorescence was detected under UV light. Isolates displaying no or weak fluorescence in contrast to Escherichia coli control strain with functional efflux pump were considered positive for overexpressed efflux pump activity.
Detected acquired β-lactamase genes
In this section of study, all phenotypically confirmed isolates and nonsuspected isolates for β-lactamase activity, and isolates with negative phenotypic confirmatory tests were screened for corresponding genes using multiplex PCR and respective alleles were further confirmed by sequencing (Table 3). Ninety-eight out of 99 suspected isolates for β-lactamase activities were positive for one or more β-lactamase genes. No acquired β-lactamase gene was detected for 37 nonsuspected isolates. Also, one nonsusceptible isolate with negative confirmatory test was negative for any β-lactamase gene. CTX-M group 1 (69 isolates), blaVEB (11 isolates), and CTX-M group 9 (9 isolates) were the most prevalent ESBL genes. BlaCTX-M-type genes from other groups were not detected. BlaCTX-M group 1 genes were detected alone or in combination with other genes in 54 out of 75 phenotypically confirmed isolates for ESBL, 8 out of 14 phenotypically confirmed isolates for simultaneous ESBL/AmpC hyperproducer, and 3 out of 9 phenotypically confirmed isolates for only AmpC hyperproducer activities. Plasmid-mediated AmpC cephalosporinases were detected in 26 isolates (11.8%) alone or in combination with other genes. These isolates were recovered from internal (20 isolates, 9.1%), surgery (4 isolates, 1.8%), intensive care unit, and burn (1 isolate each) wards. The most prevalent genes among this class were blaCMY (17 isolates, 7.7%), followed by blaFOX (4 isolates, 1.8%) and blaDHA (3 isolates, 1.3%), and blaLAT (2 isolates, 0.9%). BlaCMY gene was detected in 1 out of 75 phenotypically confirmed isolates for ESBL, 6 out of 14 phenotypically confirmed isolates for simultaneous ESBL/AmpC hyperproducer, and 7 out of 9 phenotypically confirmed isolates for only AmpC hyperproducer activities. No carbapenemase gene was detected among carbapenem-nonsusceptible isolates.
Detected β-Lactamase Genes and Related Antimicrobial Susceptibility Patterns of Escherichia coli Isolates
All isolates carrying combinations of at least two genes are presented.
CTX-M group 1.
CTX-M group 9.
Am, amikacin; AMC, amoxicillin/clavulanate; Amp, ampicillin; CAZ, ceftazidime; CFO, cefoxitin; CIP, Ciprofloxacin; CTR, ceftriaxone; CTX, cefotaxime; ETP, Ertapenem; FEP, cefepime; Gen, gentamicin; IMP, imipenem; MRP, meropenem; PIZ, piperacillin/tazobactam; SXT, trimethoprim/sulfamethoxazole.
Mutational analysis of chromosomal AmpC promoter/attenuator region
According to the results, 33 isolates were nonsusceptible to cefoxitin and suspected for AmpC hyperproducer activity, from which 14 isolates were confirmed for simultaneous ESBL/AmpC and 9 isolates for only AmpC activities. Accordingly, all 33 isolates were subjected to mutational analysis of chromosomal AmpC promoter/attenuator region. Based on sequence analysis of the chromosomal ampC genes, 10 isolates were recognized to have mutations. Seven of 10 mutants were previously confirmed for ESBL/AmpC activity. According to the results of multiplex PCR, blaCMY-2 gene was responsible for AmpC activity detected in remaining 7 out of 14 isolates that were confirmed for ESBL/AmpC activity. Two of 10 mutant isolates was previously confirmed for only AmpC activity. One cefoxitin-nonsusceptible isolate with promoter/attenuator mutation had no cAmpC activity. Based on sequence analysis, five isolates had −32 T → A mutation that created altered consensus −35 promoter box. Three isolates were mutant at both −42 and −18 positions, which displaced promoter region. These isolates also were recognized to have overexpressed efflux pump activity based on Cartwheel method and were nonsusceptible to ertapenem. Two isolates also had spacer mutations that increased the spacer to 18 base pair. Other mutations in the ampC promoter/attenuator regions of 10 E. coli isolates are presented in Fig. 2.

Sequences of chromosomal AmpC promoter/attenuator region showing mutations detected in 10 Escherichia coli isolates overproducing AmpC. Five isolates had −32 T → A mutation that altered TTGTCA (−35 box in wild-type sequence) to strong consensus −35 promoter box. Three isolates had C → T and G → A mutation at −42 and −18 positions, which displaced promoter region. Two isolates also had spacer mutations as −13 TT and GT insertion, which increased the spacer length to 18 base pair. Three isolates were recognized to have attenuator mutations at +17 and +23 positions. EA, East Azerbaijan; WA, West Azerbaijan.
Mutational analysis of acrR and marR efflux regulatory genes
According to the results of EtBr-agar Cartwheel assay, we detected 16 MDR isolates displaying overexpressed efflux pump activity, from which five isolates had low-level ciprofloxacin MICs in the range of 8–16 μg/mL, which were fully reverted to susceptible range (≤1 μg/mL) in the presence of efflux inhibitor. MIC rates for remaining 11 high-level ciprofloxacin-resistant isolates (MICs 16–256 μg/mL) were reverted to intermediate range (2 μg/mL). All 16 isolates demonstrating overexpression of efflux pump activity were selected for evaluation of acrR and marR genes. Based on the results of DNA sequencing, 15 isolates had alterations in marR, eight isolates had alterations in acrR, and 11 isolates had mutations in marO, using the GenBank sequences found under accession number M96235 for mar regulon and AE000152 U00096 for Escherichia coli K12 wild-type acrR gene for comparison (Table 4). Mutations detected in this study include G103S and Y137H (n = 15 isolates), K62R (n = 8), S3N (n = 3), and A53E (n = 1) in marR sequence and L109 (n = 2) and L190 (n = 1) frameshift mutations and T12M, T213I, N214T, I113V, and H115Y point mutations (n = 5) in acrR sequence. The latter mutations, except T12M, were only detected in high-level ciprofloxacin-resistant isolates. In addition, we detected A1332C (n = 11), C1375T, A1331G, and C1379T (one isolate each) alterations in the operator-promoter region of marR (marO).
Phenotypic and Genotypic Characteristics of 16 Multidrug-Resistant Escherichia coli Isolates Displaying Overexpressed Efflux Pump Activity
blaCTX.M group 1.
blaCTX.M group 9.
Intermediate.
C-ampC, chromosomal ampC cephalosporinase gene; CXM, cefuroxime; fs, frameshift; ST, sequence type.
Discussion
This study revealed that the clonal spread of MDR E. coli is emerging in Azerbaijan and resistance determinants responsible for MDR phenotypes are stably circulating through specific clones. Considering intrinsic mechanisms, AmpC cephalosporinases confer resistance to penicillins, cephalosporins, and cephamycins.6,8 In this study, we detected 10 (4.5%) isolates hyperproducing chromosomal AmpC as a result of various mutations in promoter/attenuator region of ampC genes. According to the report of Jørgensen et al., 25 these mutations are responsible for moderate (−32 T → A and −13 TT/GT insertion) to pronounced (−42 C → T and −18 G→A) resistance to cephalosporins. Our MICs for these isolates confirm moderate and intense resistance to cefuroxime for −32/−13 (MICs 16–32 μg/mL) and −42/−18 (MICs ≥32 μg/mL) mutations, respectively, but in comparison to Jørgensen's report, higher levels of resistance were observed against ceftazidime (MIC levels ≥8 μg/mL) and cefotaxime (MIC levels ≥128 μg/mL) in our cAmpC hyperproducer isolates, which may be related to higher rates of acquired ESBLs in these isolates. Overexpression of AcrAB-TolC may also lead to MDR phenotype by inactivation of AcrR repressor and overexpression of MarA transcriptional activator due to mutations at the acrR and marR repressors. 26 Detected mutations in marR and acrR loci of 5 low-level and 11 high-level ciprofloxacin-resistant isolates indicated that despite acrR mutations, marR mutations had no effect on repressor activity, which is consistent with the results of Vinué et al. 27 The existence of different efflux activities despite the same marR mutations between low- and high-level ciprofloxacin-resistant isolates may be related to increased expression of other efflux systems such as MdtEF and YdhE, in addition to AcrAB. 27
Among acquired mechanisms, acquisition of resistance determinants for β-lactam (ESBLs, p-AmpCs, and carbapenemases) and non-β-lactam drugs is the most important mechanism for emergence of MDR pattern.7,9,28 In our isolates, CTX-M group 1, VEB, and CTX-M group 9 were the most prevalent ESBLs. The high rates of ESBLs, particularly in the case of CTX-M group 1, were in accordance with our recent report from West Azerbaijan and the other area of Iran, including northeast region and Tehran province.29–31 On the other hand, comparing our values with published data from the SENTRY Program in 2007 (16.0%) and 2010 (33.8%) indicates the high rates of CTX-M-type ESBLs in comparison with other parts of the world. These rates may represent the fact that mobile genetic elements carrying blaCTX-M and other MDR determinants are stably circulating in our clinical settings under high selection pressure.32,33 Recent data from Iran represent an ever-increase rise of antibiotic resistance among hospital- and community-acquired E. coli isolates. Based on the reports, the resistance rates vary from region to region of Iran, including 6–73% for aminoglycosides, 10–87% for third-generation cephalosporins, 8.3–64% for fluoroquinolones, and 5.7–93.4% for co-trimoxazole. 34 Even though our results (Table 2) are in the range of reported values, successful surveillance and treatment of MDR infections will not be feasible without identification of indigenous mechanisms involved in the antimicrobial resistance and evaluation of epidemiological relatedness of MDR strains in each region. In recent decade, Escherichia coli ST131 has been known as a major clone linked to the spread of blaCTX-M-15 and other MDR genes,35,36 and has now been identified to be the cause of variety of hospital- and community-acquired extraintestinal infections, particularly urinary tract infection and bacteremia.37–41 In this study, 6 out of 16 MDR E. coli isolates with overexpressed efflux pump activity belonged to ST131 clone and mostly had blaCTX-M-type (group 1) genes (Table 4). These results indicate that clonal occurrence of overexpressing efflux pump isolates is emerging in Azerbaijan hospitals.
Our study faced some limitations. First, the numbers of examined E. coli isolates were not equal from each province. Second, we used the E test strips rather than broth microdilution method for MIC evaluation. Third, a phenotypic confirmatory method performed in this study using Rosco confirmatory kits were not approved by CLSI, but we used the same β-lactamase inhibitors recommended by CLSI.
Based on our findings, unfortunately, the rate of MDR E. coli isolates is very high in clinical settings of Azerbaijan. Accordingly, we are in an alarming situation that requires serious attention on an international scale, particularly due to clonal occurrence of MDR, overexpressing efflux pump isolates. The lack of continuous monitoring and surveillance programs, nonstandardization of indigenous antimicrobial susceptibility tests, physician coercion from the patients to prescribe medication, and overuse, self-medication, and empirical use of third-generation cephalosporins and carbapenems without careful monitoring of resident MDR isolates in hospital settings are important factors exacerbating selective pressure in emergence of MDR strains in Azerbaijan.
Footnotes
Acknowledgment
This article was based on data obtained from a research project registered and financially supported by Islamic Azad University, Maku branch, Maku, Iran.
Disclosure Statement
No competing financial interests exist.