
Abstract
Drug resistance is of great importance to human and animal health, but wild environments are still poorly understood in terms of their function as reservoirs of dangerous microbes and resistance determinants. The aim of the study was to determine the antibiotic susceptibility and virulence factors of Enterococcus bacteria from wildlife in Tuscany, Italy. Of the 36 mammalian fecal samples, 52 isolates were derived and classified as Enterococcus faecium (46% of isolates), Enterococcus hirae (19%), Enterococcus faecalis (13%), Enterococcus gallinarum (8%), Enterococcus casseliflavus (6%), Enterococcus durans (4%), Enterococcus mundtii (2%), and Enterococcus canintestini (2%) using both matrix-assisted laser desorption/ionization-time of flight mass spectrometry and methods based on analysis of genetic material. The isolates tested showed the most frequent resistance to tetracycline (36.5% isolates), ciprofloxacin (36.5%), and erythromycin (25%). Three isolates showed high level of resistance (minimal inhibitory concentration ≥1,024 μg/mL) to vancomycin and teicoplanin, and 15% of the isolates demonstrated multidrug resistance. No isolate resistant to ampicillin, linezolid, or streptomycin was found. Among resistance genes, aac(6)-Ii (50% isolates), msrA/B (48%), msrC (42%), and tetM (31%) were identified most frequently. All E. faecium and E. faecalis isolates were positive for the efaAfm and efaAfs genes, respectively. Other virulence-associated genes, that is, gelE, cylA, asa1, esp, ace, orf1481, ptsD, and sgrA, were found in the majority of E. faecalis and several E. faecium isolates. Multilocus sequence typing analysis performed for selected isolates revealed three new sequence types. The results obtained indicate that wild mammals might act as reservoirs of resistance and virulence determinants that could be transferred between different ecosystems.
Introduction
Bacteria of the genus Enterococcus are Gram-positive, catalase-negative, nonspore-forming, facultative anaerobic cocci belonging to a group of lactic acid bacteria (LAB). They are regarded as commensals inhabiting the oral cavity, gastrointestinal, and genitourinary tracts in humans and animals and various environments, including plants, soil, water, and traditional fermented food, and dairy products. The broad range of their tolerance to physical and chemical factors, that is, temperature (ability to grow at 5–50°C), oxygenation (ability to grow in both aerobic and anaerobic atmosphere), pH (4.6–9.9), and salinity (Enterococcus faecalis is able to grow in 6.5% NaCl), contributes to the prevalence of these bacteria in the environment.
1
Currently, 61 species of Enterococcus are known (NCBI database),
Several factors have been characterized as important for the virulence of enterococci. Among them, enterococcal surface protein (Esp), aggregation substance (AS), E. faecalis adhesin (Ace), and adhesion-associated protein EfaA (E. faecalis endocarditis antigen A) can facilitate the colonization process in the host by increasing adhesion to the epithelium, bacterial aggregation, or biofilm formation. 12 Other factors such as gelatinase (Gel) and hyaluronidase (Hly), which are lytic enzymes associated with tissue damage, and cytolysin (Cyl) are active against erythrocytes and Gram-positive bacteria. 13 The E. faecium isolates from clinical samples are strongly associated with such genetic markers as esp, hyl, sgrA (coding for surface serine-glutamate-repeat-containing-protein A), ecbA (coding for collagen binding protein), acm (encoding adhesin of collagen), ptsD (encoding phosphotransferase system IID enzyme subunit), orf1481 (encoding sugar binding protein, marker of genomic island), and IS16 (transposase that enhances genomic plasticity). 14
Enterococci have raised research interests not only due to their pathogenic role as opportunistic bacteria but also due to their strong ability to acquire, express, and transfer antimicrobial resistance genes. Enterococci characterized by acquired resistance, including resistance to standard anti-enterococcal antibiotics, that is, ampicillin, vancomycin (VRE; vancomycin resistance enterococci), and aminoglycosides (high-level resistance), are mainly isolated from clinical cases from humans, 15 but also animals, particularly livestock, may be reservoirs of drug-resistant strains.16–18 Moreover, it has been shown that clones associated with human infections occur in animals. 19
Wildlife are generally not exposed to clinical antibacterial drugs, but they can acquire resistant bacterial strains by contact with the contaminated environment, and water polluted with feces appears to play a key role. Intestinal microorganisms from humans and animals (livestock, aquaculture, and human companion animals) undergoing antibiotic therapy are subject to constant release into the environment through wastewater or manure. Sewage and outflows from fields fertilized with manure get into rivers, and fecal bacteria may thus be transported over long distances. In addition, migrating birds that can acquire antibiotic-resistant bacteria through contact with contaminated water or food waste from garbage act as vectors and play an important role in the spread of resistance. It should also be taken into account that many antimicrobials can be excreted by humans and farm animals or pets in an active form and persist in the environment, thereby promoting the selective advantage of antibiotic-resistant bacteria.19,20
Monitoring the antibiotic resistance and virulence of intestinal bacteria from wildlife is a useful tool to get information on the spread of dangerous strains and resistance determinants in natural ecosystems. Currently, to the best of our knowledge, there are only a few literature reports dealing with the issue of drug susceptibility of enterococci from wild animals (mammals, birds, and reptiles).2,3,11,17,21,22 For this reason, the aim of the present study was to investigate antimicrobial resistance and pathogenicity factors in Enterococcus bacteria from wildlife in the Apuan Alps Regional Park located in northwestern Tuscany, Italy. The studied area extends over 20,000 ha. It is inhabited by various animals, and the direct impact of humans on this environment is limited to a minimum.
Materials and Methods
Materials
A total of 36 fecal samples of wild animals, including 20 wolves (Canis lupus), 6 badgers (Meles meles), 6 mouflons (Ovis aries musimon), 2 foxes (Vulpes vulpes), and 2 wild boars (Sus scrofa), were collected from the Apuan Alps Regional Park (Tuscany, Italy) in 2017. No information on the health status of the animals was available. The samples were stored frozen at −20°C until analysis, that is, up to 3 months. Enterococci were prepropagated on tryptic soy broth (BTL, Poland) at 37°C for 24 hr and then seeded on Slanetz and Bartley agar (BTL). Plates were incubated overnight at 37°C, and then individual colonies with different morphology and color were collected and propagated as pure cultures. Bacteria were stored at −80°C until further analysis.
Identification of Enterococcus isolates
Collected bacteria were identified by matrix-assisted laser desorption/ionization-time of flight mass spectrometry (MS) using a standard extraction method as previously described 23 and an UltrafleXtreme MALDI TOF mass spectrometer (Bruker, Germany). The mass spectra obtained from each isolate were processed with the MALDI Biotyper 3.0 software package (Bruker), and the results were shown as the top 10 identification matches along with confidence scores ranging from 0.000 to 3.000. According to the manufacturer's criteria, a log(score) below 1.70 does not ensure reliable identification; a log(score) between 1.700 and 1.999 allows identification to the genus level; a log(score) between 2.000 and 2.299 means highly probable identification at the genus level and probable identification at the species level, and a log(score) higher than 2.300 (2.300–3.000) indicates highly probable identification at the species level. Each sample was analyzed in triplicate.
In the case of isolates for which the log(score) values obtained by MALDI-TOF MS were lower than 2.000, an identification based on 16S rDNA and rpoA sequence analysis has been performed. The 16S rRNA and rpoA genes were amplified by PCR using primers fD1 5′-AGAGTTTGATCCTGGCTCAG-3′ and R1530 5′-AAGGAGGTGATCCAGCCGCA-3′ and rpoA-21-F 5′-ATGATYGARTTTGAAAAACC-3′ and rpoA-23-R 5′-ACHGTRTTRATDCCDGCRCG-3′, as it was described before.24,25 The sequences of amplicons were determined by a commercial DNA sequencing service provider (Genomed, Warsaw, Poland) using Sanger method and compared to reference sequences available in the GenBank database using the NCBI BLAST algorithm.
Differentiation of E. casseliflavus and E. gallinarum by internal transcribed spacer PCR/restriction fragment length polymorphism
Since MALDI-TOF MS, as well as 16S rDNA and rpoA sequence analysis, was characterized by low resolution in relation to E. gallinarum and E. casseliflavus isolates, the identification of these species was additionally based on analysis of 16S–23S rDNA regions. Internal transcribed spacer (ITS)-PCR was carried out using universal primers L1 and G1. 26 The PCR products were digested using RasI restriction enzyme (Thermo Scientific) for 4 hr at 37°C. 27 The study involved the reference strains E. casseliflavus ATCC 700327 and E. gallinarum ATCC 49573. PCR products and restriction fragments were separated by electrophoresis in a 3% (wt/vol) high-resolution agarose (Blirt, Poland) gel with SimplySafe stain (Eurx, Poland) in 0.5 × tris-borate-EDTA (pH 8.0) buffer. Gels were documented with a Gel Doc apparatus (Bio-Rad), and band sizes were determined using the M100–1,000 bp DNA Ladder (Blirt) and Quantity One software (Bio-Rad).
Antibiotic susceptibility
Antibiotic susceptibility of all bacterial isolates was determined by the broth microdilution assay according to the Clinical and Laboratory Standards Institute guideline. 28 The test was carried out using Mueller-Hinton broth (Oxoid, United Kingdom) and antimicrobial substances such as ampicillin (Roth), tetracycline, erythromycin, streptomycin, gentamycin, chloramphenicol, vancomycin, teicoplanin, ciprofloxacin, linezolid (Sigma-Aldrich, Poland), and tiamulin (BIOWET DRWALEW S.A., Poland). Stock solutions of the drugs were prepared as described previously 29 ; vancomycin, teicoplanin, and ciprofloxacin were dissolved in distilled water and linezolid in dimethyl sulfoxide.
Before antimicrobial susceptibility testing, the enterococci were cultured on cation-adjusted Mueller-Hinton broth (CAMHB) overnight at 37°C. 30 Inocula were prepared by suspending the bacteria in 0.85% NaCl to the turbidity of a 0.5 McFarland standard using a DENSI-LA-METER II (Erba, Czech Republic). Microdilution plates were inoculated with 50 μL of a 1:100-diluted (in CAMHB) inoculum and 50 μL of the appropriate antibiotic solution. Plates were incubated at 37°C in aerobic conditions for 24 hr, and minimum inhibitory concentration (MIC) values were read as the lowest concentration of an antimicrobial agent at which visible growth was inhibited. E. faecalis ATCC 29212 was used as a quality control strain.
Interpretation of the results was based on the breakpoint values suggested by CLSI 31 (for ampicillin, vancomycin, teicoplanin, linezolid, erythromycin, tetracycline, chloramphenicol, and ciprofloxacin) and by EUCAST 32 (for streptomycin and gentamicin). For tiamulin, no cutoffs were proposed due to the lack of a bimodal distribution of MICs and insufficient number of isolates.
Detection of resistance and virulence-associated genes
Genomic DNA from enterococci was isolated using a GeneMATRIX Bacterial & Yeast Genomic DNA Purification Kit (Eurx) following the manufacturer's instruction with some modifications consisting in prolongation of the bacterial incubation in a lysis buffer up to 60 min. PCRs based on gene-specific primers (Supplementary Table S1) were used to detect the presence of resistance genes, that is, tetL, tetM, tetK, tetO, ermA, ermB, mefA/E, msrA/B, msrC, aac(6)-Ii, aph(3′)-IIIa, aac(6′)-Ie-aph(2′′)-Ia, ant(4′)-Ia, ant(6)-Ia, aph(2′)-Ib, aph(2′′)-Ic, aph(2′′)-Id, cat, vanA, vanB, vanC1/C2, vanM, cfr, lsaE, lsaC, vgaA, vgaAv, the Tn916/Tn1545 transposon integrase gene int-Tn, as well as genes associated with virulence in Enterococcus spp., that is, efaAfm, efaAfs, gelE, cylA, asa1, esp, ace, orf1482, ptsD, sgrA, and IS16. PCRs were performed in an Eppendorf Mastercycler using DreamTaq polymerase (Thermo Scientific, final concentration 0.024 U/μL). The temperature program of all PCRs was as follows: initial denaturation at 95°C for 5 min, 30 cycles of 95°C for 45 sec, 48–60°C (depending on the primers used, Supplementary Table S1) for 45 sec, 72°C for 75 sec, and a final extension step at 72°C for 8 min. PCR products were separated by electrophoresis (100 V) on 2% agarose gels and visualized by SimplySafe (Eurx) staining. Wild LAB strains containing resistance genes and an integrase gene, as well as three reference Enterococcus strains (E. faecium ATCC 51559, E. faecalis ATCC 51299, and E. gallinarum ATCC 49573) containing van genes, were used as a positive control. The sequence of the PCR products (counterparts of resistance genes) obtained for the representative wild-type strains was analyzed using the NCBI BLAST algorithm
Phenotypic tests for detection of gelatinase and cytolysin
For screening the production of gelatinase, the enterococci were cultured onto Trypticase Soya Agar (Oxoid) containing 3% gelatin (Avantor Performance Materials, Poland). After overnight incubation at 37°C followed by refrigeration at 4°C for 30 min, colonies surrounded by opaque zones were considered to be gelatinase activity positive. E. faecalis ATCC 29212 was used as a positive control. 33
The production of hemolysin was determined by streaking bacterial cultures on Columbia Agar Base (Oxoid) supplemented with 5% of sheep blood (Pro Animali Company, Poland). Plates were incubated at 37°C for 24 hr either aerobically or in atmosphere containing 7% CO2. A positive result was indicated by the formation of hemolytic (clear) zones around the colonies. E. faecalis ATCC 29212 was used as a positive control. 33
Multilocus sequence typing
The multilocus sequence typing (MLST) of representative isolates of E. faecalis and E. faecium was performed according to the previously developed schemes.34,35 The sequences of primers used for MLST are shown in Supplementary Table S3. The DNA sequences of seven housekeeping genes were edited and aligned using DNAMAN ver. 4.13 software (Lynnon BioSoft, Quebec, Canada) and compared with a reference set of alleles of E. faecalis and E. faecium strains deposited in MLST web-based database
Results
Isolation and identification of Enterococcus spp.
Enterococci were isolated from 30 out of the 36 fecal samples. A total of 52 isolates were classified as bacteria of the genus Enterococcus using the MALDI Biotyper software package (version 3.0) with the reference database version 3.1.2.0 (Bruker Daltonik GmbH). One to four Enterococcus isolates were isolated from one sample. The log(score) was 2.300–3.000 for 23 (44%) isolates, 2.000–2.299 for 24 (46%), and 1.700–1.999 for 5 (10%). Lower log(score) values (1.700–1.999), regarded as reliable identification to the genus level, were obtained for five isolates, three of which were preclassified as E. casseliflavus, one as E. gallinarum, and one as E. faecium (Supplementary Fig. S1).
Based on log(score) values in the range of 1.700–3.000 among the 52 Enterococcus isolates, the following species were identified: E. faecium (24 isolates), E. hirae (10), E. faecalis (7), E. gallinarum (4), E. casseliflavus (3), E. durans (2), E. mundtii (1), and E. canintestini (1).
Species identification by sequencing of the 16S rRNA and rpoA genes
The identification based on the analysis of 16S rRNA and rpoA gene sequences was performed for five isolates, which were characterized by low log(score) values (<2.000) in the MALDI-TOF MS method. The results of the comparative analysis of the obtained 16S rDNA sequences with the reference sequences deposited in GenBank have shown that the 16S rDNA sequencing method is effective in identifying E. faecium, but does not distinguish E. gallinarum from E. casseliflavus (Table 1). Phylogenetic analysis showed that the 16S rDNA sequences of the 49WP, 50MF, and 51BG isolates (identified initially by MALDI-TOF MS as E. casseliflavus) are identical and differ from the 16S rDNA sequence of 46W isolate (initially identified as E. gallinarum) in only three positions (variation in two nucleotides and one deletion).
Results of Genetic Identification of Isolates for Which Log(Score) Values Obtained by MALDI-TOF MS Were <2.000
c. genome, complete genome; ITS-PCR/RFLP, internal transcribed spacer PCR/restriction fragment length polymorphism; MALDI-TOF MS, matrix-assisted laser desorption/ionization-time of flight mass spectrometry; NT, not tested.
The analysis of the rpoA sequences confirmed the initial identification of the 23BG isolate as E. faecium, but it was not slow to establish unambiguous species affiliation for 46W, 49WP, 50MF, and 51BG isolates (E. gallinarum, E. casseliflavus). Obtained sequences showed >99% homology to the sequence of both E. gallinarum and E. casseliflavus deposited in GenBank (Table 1). However, phylogenetic analysis showed that the sequences of the 49WP, 50MF, and 51BG isolates (E. casseliflavus) differ from the rpoA sequence of the 46W isolate (E. gallinarum) in as many as 42 positions. The rpoA sequences of the 49WP and 50MF isolates were identical and differed from the rpoA sequence of the 51BG isolate at two positions (Fig. 1).
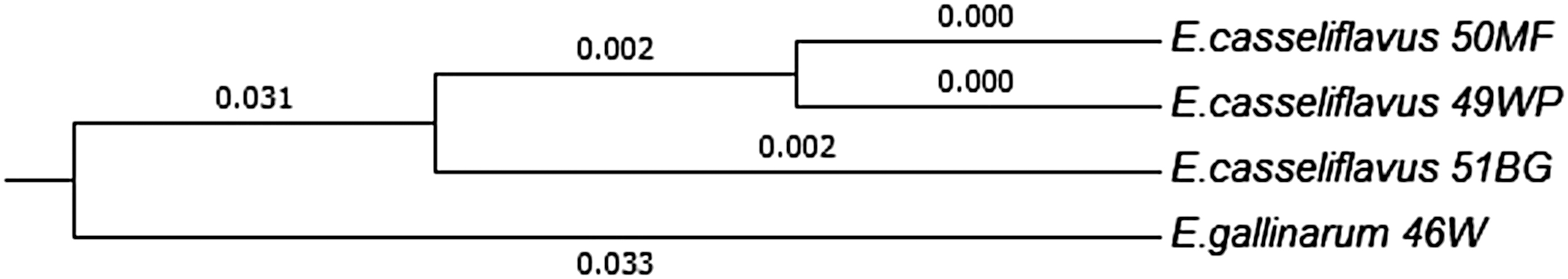
The phylogenetic relationship of Enterococcus casseliflavus and Enterococcus gallinarum isolates determined by comparative sequence analysis of rpoA gene.
Differentiation of E. gallinarum and E. casseliflavus by ITS-PCR/RFLP
Electrophoretic profiles of 16S–23S rDNA amplicons of wild isolates 46W, 49WP, 50MF, and 51BG (initially identified as E. gallinarum or E. casseliflavus) and reference E. gallinarum ATCC 49573 and E. casseliflavus ATCC 700327 strains were identical (Fig. 2A). However, the use of restrictive analysis allowed for species diversity within the tested isolates. Electrophoretic profiles contained two or three restriction fragments. They were 350 and 243 bp for E. casseliflavus and 285, 243, and 180 bp (or only 285 and 180 bp) for E. gallinarum (Fig. 2B).
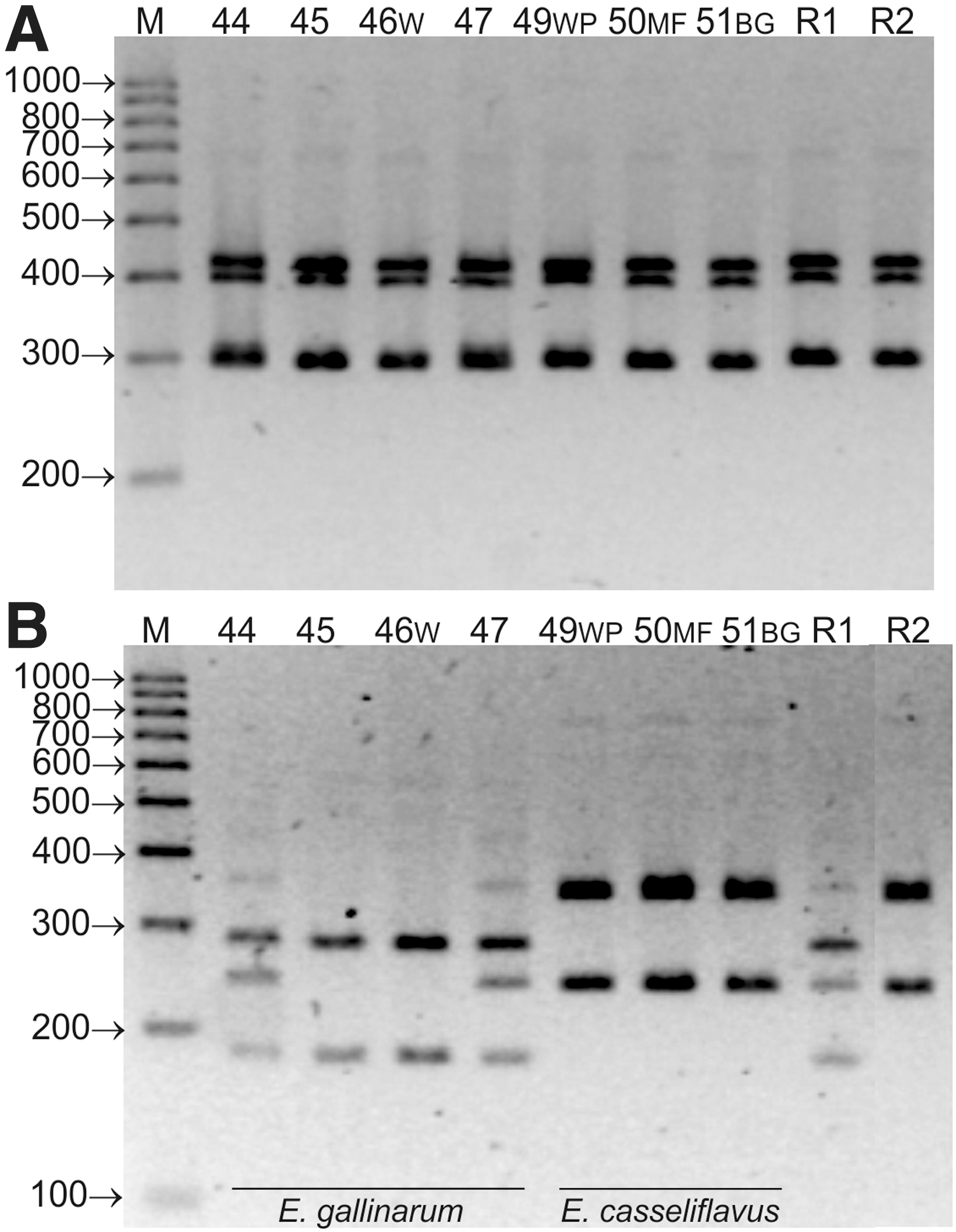
Electrophoretic profiles of 16S–23S rDNA PCR amplicons of E. gallinarum and E. casseliflavus isolates
Antimicrobial susceptibility testing
The MIC of 11 antibiotic agents was analyzed for 52 Enterococcus isolates from wild living animals (Table 2). According to the adopted criteria, no strain showed resistance to ampicillin, streptomycin, and linezolid, and the resistance to other drugs ranged from 4% to 36.5% (Table 3). The highest frequency was noted for resistance to tetracycline (36.5% resistant isolates), ciprofloxacin (36.5%), and erythromycin (25%). Resistance to vancomycin and teicoplanin (6%), chloramphenicol (6%), and gentamicin (4%, HLAR [high-level aminoglycoside resistance]) was observed much less frequently. The occurrence of intermediately susceptible isolates was found in relation to linezolid (77%), erythromycin (40%), ciprofloxacin (23%), and chloramphenicol (10%). Eighteen (35%) out of the 52 Enterococcus strains did not show resistance to any antibiotic, and multidrug resistance (MDR) was detected in 8 (15%) isolates.
Distribution of Minimum Inhibitory Concentrations of Antibiotics Among Enterococci Tested


Number of Enterococcus Isolates Displayed Resistance or Intermediate Susceptibility
Bold font in the last line highlights aggregate results.
Resistant for min 3 groups of antimicrobial agents (the analysis excluded tiamulin for which the break points have not been established).
HLAR, high level aminoglycoside resistance; I, intermediate; R, resistant.
AMP, ampicillin; CHL, chloramphenicol; CIP, ciprofloxacin; ERY, erythromycin; GEN, gentamicin; LIZ, linezolid; STR, streptomycin; TEIC, teicoplanin; TET, tetracycline; VAN, vancomycin.
Only E. faecium and E. faecalis isolates showed resistance to erythromycin, gentamicin, and chloramphenicol. Resistance to vancomycin and teicoplanin (MIC ≥1,024 μg/mL) was detected in one isolate of E. faecium (26W), one isolate of E. hirae (34MF), and one E. mundtii (52WP) and, interestingly, these isolates demonstrated simultaneous resistance to tetracycline and ciprofloxacin (Supplementary Fig. S1).
The cutoffs for tiamulin were not established due to the lack of a bimodal distribution of MICs for most of the Enterococcus species tested and the insufficient number of isolates, but the bimodal MIC distribution observed for the E. faecium and E. hirae isolates indicated a value of 32 μg/mL as a break point. High MIC values, that is, ≥32 μg/mL, were characteristic for a majority (81%) of the enterococci, and for the remaining 19% of isolates, the tiamulin MICs ranged from 0.5 to 8 μg/mL (Table 2).
A clear bimodal distribution of MICs indicative of acquired resistance was observed for vancomycin teicoplanin and tetracycline. For the other drugs tested, MICs showed rather an extended than bimodal distribution in the case of most of the Enterococcus species (Table 2).
Detection of antibiotic resistance genes
All Enterococcus isolates (both phenotypically resistant and susceptible) were screened by PCR for the presence of genetic determinants of resistance.
The tet genes conferring resistance to tetracyclines were found in 17 (33%) isolates, including 15 phenotypically tetracycline-resistant and 2 susceptible isolates. The tetM gene was significantly more frequent (in 16 isolates) than the tetL (in 5 isolates) and was present in the isolates of various Enterococcus species, while tetL was found only in E. faecalis and E. faecium (Tables 2 and 4; Supplementary Fig. S1).
Number of Enterococcus Strains Carrying Resistance Genes, Integrase Gene and Virulence-Associated Genes

The ermB (coding methylase), msrA/B, and msrC (coding for ATP-dependent efflux pumps, ABC transporters) genes, conferring resistance to macrolides, were present in 6%, 48%, and 42% of isolates, respectively. The ermB gene was detected in 3 (6%) out of 13 erythromycin-resistant isolates, and msrA/B and msrC were found in both resistant and susceptible isolates. The msrA/B and msrC genes were present simultaneously in most cases and were mainly found in E. faecium isolates. The ermB methylase gene was detected only in three isolates (2/3 resistant E. faecalis and 1/10 E. faecium) with high MIC values, that is, ≥128 μg/mL (Tables 2 and 4). The ermB gene was always associated with the presence of the tetM gene, and the msrA/B and msrC genes generally coexisted with the aac(6′)-Ii gene (encoding 6′-N-aminoglycoside acetyltransferase, a member of a subfamily of AAC(6′)-I enzymes) (Supplementary Fig. S1).
Among genes determining resistance to aminoglycosides, we noted the widespread presence of the aac(6′)-Ii gene (50% of isolates) and sporadic occurrence of the ant(4′)-Ia (8%), aph(3′)-IIIa (2%), aac(6′)-Ie-aph(2′′)-Ia (2%), and ant(6)-Ia (2%) genes. The aph(3′)-IIIa gene encoding aminoglycoside-3′-phosphotransferase and the aac(6′)-Ie-aph(2′′)-Ia gene, which encodes the bifunctional aminoglycoside-modifying enzyme AAC(6′)-APH(2′), were present only in one E. faecalis isolate with a high level of resistance to gentamicin (MIC ≥1,024 μg/mL). The presence of other aminoglycoside resistance genes was not correlated with phenotypic resistance to streptomycin or gentamicin (Table 2).
The cat gene encoding chloramphenicol acetyltransferase, which converts chloramphenicol to inactive diacetyl chloramphenicol, 28 was present in three (6%) isolates, including one phenotypically chloramphenicol-resistant E. faecalis isolate (MIC = 64 μg/mL) and two susceptible isolates (Tables 2 and 4).
The int-Tn gene coding for the integrase of the Tn916/Tn1545 family of conjugative transposons was detected in 29% of isolates, often in coexistence with tetM gene.
None of the Enterococcus isolates contained genes conferring resistance to pleuromutilins, that is, lsaC, lsaE, vgaA, and vgaAv genes, which code for ABC transporters in Gram-positive bacteria, or the cfr gene coding for RNA methyltransferase. 28 We did not detect either vancomycin resistance genes, that is, vanA, vanB, vanC1/C2, vanM or tetK, tetO, ermA, mefA/E, aph(2′)-Ib, aph(2′′)-Ic, and aph(2′′)-Id genes.
Detection of virulence genes and production of gelatinase and cytolysin
Among the virulence-associated genes, genes coding for cell wall adhesion antigens were most widespread in enterococci tested. The presence of the efaAfm and efaAfs genes was recorded in all isolates of E. faecium and E. faecalis, respectively, and in one isolate of E. canintestini. The gelE, cylA, asa1, esp, and ace genes were present in several E. faecalis isolates and in two E. faecium isolates. Five out of the seven tested E. faecalis isolates contained two virulence genes (three isolates contained gelE+ace, one isolate gelE+asa1, and one isolate esp+ace), and one isolate (E. faecalis 3W) harbored three virulence genes (asa1+cylA+esp). The latter one seems to be particularly dangerous because apart from three virulence genes it also contained six resistance genes (tetM, ermB, aph3-IIIa, ant(4′)-Ia, aac(6′)-Ie-aph(2′′)-Ia, msrA/B) and the integrase gene int-Tn. None of the Enterococcus isolates contained the hyaluronidase (hyl) gene. In 8 out of 24 E. faecium isolates, genetic virulence markers other than efaAfm have been reported. These were: gelE (1 isolate), esp (1), ace (1), sgrA (5), ptsD (1), and orf1481 (2). Three E. faecium isolates contained more than one of these virulence genes (isolate No. 11 contained orf1481+sgrA, isolate No. 15 gelE+ace, and isolate No. 29 orf1481+ptsD) (Supplementary Fig. S1; Table 4).
The presence of the gelE gene was found for all isolates (five isolates of E. faecalis and one isolate of E. faecium), which correlated with the results of the phenotypic test confirming the breakdown of gelatin by a bacterial enzyme. A similar correlation between the genotype and the phenotype was observed in the case of the cylA gene, which was identified in one isolate of E. faecalis. However, it is worth emphasizing that beta hemolysis was seen on the Columbia medium supplemented with blood after incubation of the plates in the atmosphere containing 7% CO2, but not in aerobic conditions.
Multilocus sequence typing
In three of the six isolates analyzed (two isolates of E. faecalis and four isolates of E. faecium) by MLST, new STs were determined, that is, ST1574, ST1575, and ST1576. They referred to E. faecium isolates, including VRE isolate 26W, isolate 9W showing high degree of resistance to gentamycin, and 14W—characterized by high degree of resistance to ciprofloxacin. In the E. faecium 23BG, we identified the previously described ST530, and the isolates E. faecalis 2W and 3W represented ST21 and ST16, respectively. E. faecium isolates were assigned to CC17, CC94, and CC117 clonal complexes and E. faecalis to the CC16 and CC21 (Table 5).
Multilocus Sequence Typing Results for Representative E. faecalis and E. faecium Isolates
Subscript means the MIC values.
CC, clonal complex; MIC, minimal inhibitory concentration; ST, sequence type.
Discussion
To date, only a few studies have investigated the distribution and antibiotic susceptibility of Enterococcus species in the fecal material of wild mammals.16,21,28 In the examined samples, most of which came from wolves (20/36), we identified eight species of enterococci, among which the highest percentage (46%) were E. faecium isolates. Similar results were obtained by Gonçalves et al., 37 who showed that E. faecium (51%) and, with lower frequency, E. hirae (28%) and E. faecalis (12%) dominated among the enterococci from feces of Iberian wolves.
Our studies have shown that wild mammals inhabiting the Apuan Alps Regional Park in Tuscany carry antibiotic-resistant Enterococcus strains. These results correlate with the data presented by Busani et al., 16 who showed very high frequency of resistance to tetracycline (85.5–87%), erythromycin (75–90%), and ciprofloxacin (28–47%) among isolates of E. faecium and E. faecalis collected in 1996–2000 in Italy from raw meat products, farm animals (poultry, swine), and human clinical infections. The resistance to all these antibiotics was much more frequent in E. faecium than in E. faecalis. Antibiotic-resistant Enterococcus strains have also been identified in wild animals. As in our study, a significant percentage of isolates resistant to tetracycline (55%) and erythromycin (22%) were found in enterococci from Iberian wolves living in Portugal, while the frequency of resistance to ciprofloxacin was more than twice lower (15%); resistance to ampicillin, chloramphenicol, streptomycin, and gentamicin was reported sporadically (≤4% of resistant isolates). 37 Drug-resistant Enterococcus strains were also found among wild animals in Poland, but the incidence of resistance was lower compared to that observed for the pool of isolates tested in this work. 17
In the isolates analyzed, the resistance to ciprofloxacin seems to be intrinsic, since the MIC values (≤16 μg/mL) for most isolates recognized as resistant were slightly higher than the established cutoffs. Only one isolate (E. faecium 14W) exhibited a high MIC, that is, 256 μg/mL, indicating acquired resistance, which may be due to a mutation in the gyrA or parC genes or occurrence of efflux pumps. 38
Almost all tetracycline-resistant isolates harbored the tetM gene coding for ribosomal protection protein, and four isolates additionally contained tetL, which codes for energy dependent efflux protein. 29 The occurrence of these genes, most often in coexistence, is common in enterococci, including isolates from wild living mammals and wild birds.6,11,37 Our research is also consistent with the reports of Grassotti et al., 2 who observed occasional occurrence of the tetM and tetL genes in phenotypically susceptible Enterococcus isolates. The common presence of the msrA/B and msrC genes (encoding ATP-dependent efflux pump, which determines resistance to 14-membered and 15-membered macrolides and streptogramin type B) 39 in E. faecium and sporadic in E. faecalis, observed in our research, have been also noted by several other authors.2,40,41 In addition, Grassotti et al. 2 detected the msrC gene in both erythromycin-resistant and erythromycin-sensitive enterococci derived from monkeys and noted the absence of the ermB gene in these isolates. Different results were presented by Gonçalves et al. 37 in relation to enterococci from wild Iberian wolves, among which almost half of the resistant isolates contained the ermB gene.
The low percentage of aminoglycoside-resistant strains reported in these studies is in line with the results obtained with enterococci from wild mammals inhabiting other countries.2,17,37 The aph(3′)-III and aac(6′)-Ie-aph(2′′)-Ia genes, which we detected in one isolate showing high-level resistance to gentamicin and the ant(6)-Ia gene present in one streptomycin susceptible isolate, are most frequently detected genes in the aminoglycoside resistant enterococci.6,42 The presence of the ant(6)-Ia gene in phenotypically sensitive isolates has been previously noted in other LAB. 29 In the enterococci tested, no susceptibility to kanamycin was determined, but the aac(6)-Ii gene, which catalyzes the acetylation of the 6′amino group of kanamycins A and B, neomycin, and tobramycin, was present in all E. faecium isolates and in single isolates of other species. Our results are consistent with the data described in the literature, according to which the production of chromosomally encoded 6′-N-aminoglycoside acetyltransferase type Ii (Aac(6)-Ii) is characteristic for E. faecium. 42
The presence of the Tn916/Tn1545 transposon integrase gene (int-Tn) observed in 29% of the isolates tested has been previously reported for E. faecium and E. faecalis isolates from humans, livestock, and wildlife, and similar to our studies, this gene was found to coexist with the tetM, ermB, or aph(3′)-IIIa genes.37,43
The glycopeptide vancomycin is the first choice alternative to the penicillin–aminoglycoside combination for treatment of enterococcal infections, and therefore, the rapid spread of VRE strains, especially E. faecalis and E. faecium, has been of particular concern. 4 The vancomycin-resistant enterococci detected in this study showed high level of resistance (MIC ≥1,024 μg/mL) to both vancomycin and teicoplanin, but they did not contain the van genes (vanA, vanB, vanC1/C2, vanM) conferring resistance. This indicates the presence of some other mechanism of acquired resistance to glycopeptide antibiotics in these VRE strains. Many authors have demonstrated the presence of VRE isolates in animal samples. They were often found to colonize the gastrointestinal tract of broilers and pigs from Italian farms 44 and occur in meat (35% isolates) and in environmental samples (26.5% isolates) collected in Italy. 45 They have also been found among enterococci isolated from wild mammals in Poland (10% isolates) and Portugal (VRE were recovered in 8/365 analyzed fecal samples from Iberian wolves and Iberian lynx).17,46
The elevated MIC values for tiamulin ≥32 μg/mL observed in the majority (81%) of the tested isolates seem to be characteristic of Enterococcus bacteria. Jones et al. 47 reported tiamulin MIC90 >32 μg/mL for all (71) tested enterococci of human and animal origin. In the isolates tested, we did not detect the cfr, lsaE, lsaC, vgaA, and vgaAv genes that confer resistance to pleuromutilins, lincosamides, and streptogramin A and have been previously described for enterococci 48 or other Gram-positive bacteria. 29
The ST16 ST identified in the MDR isolate 3W is one of the most prevalent STs detected in E. faecalis in the world. 34 They are isolated mainly from hospitalized patients (MLST database),*** and the strong ability of ST16 strains to create biofilm may contribute to their pathogenicity. 49 The E. faecalis strains of the ST21, determined in the 2W isolate, are also frequently found and have been detected in hospitalized patients, environmental samples, food of animal origin, and livestock (MLST database). Clonal complex 17, to which two of the analyzed E. faecium strains (9W and 23BG) were qualified, is high-risk genetic lineage associated with the majority of nosocomial E. faecium outbreaks and clinical infections worldwide. 50 The results of our research are partly consistent with literature information, according to which CC17 strains usually show resistance to ampicillin, quinolones, glycopeptides, gentamycin, and erythromycin and have the Esp. 50
Our studies have shown that enterococci from wild mammals in Italy contained virulence genes whose products allow strains to colonize and invade host tissue and translocate through epithelial cells and avoid host immune response. 51 The results are consistent with the observations of other authors who noted the widespread occurrence of virulence genes in E. faecalis and the much lower frequency in other species of Enterococcus from both clinical6,52 and nonclinical samples. 53 Poeta et al. 22 reported a high frequency of the gelE gene in E. faecalis from wild animals in Portugal, but the ace and esp genes have been only detected in ≤9.6% of isolates of this species. Completely different results were shown by Gonçalves et al., 37 who found no virulence-associated genes (gelE, agg, ace, cpd, fsr, esp, hyl, cyl) in enterococci (n = 227), including E. faecalis, from wild Iberian wolves. The presence of the surface adhesin genes efaAfs and efaAfm detected in all E. faecalis and E. faecium isolates in this study, respectively, is in agreement with the previous studies on enterococci of various origin.54,55 It is also noteworthy that some of the tested strains of E. faecium contained putative virulence markers, that is, esp, sgrA, ptsD, and orf1481, that are strongly associated with hospital strains, mainly clade A1. 14
Concluding, our studies have shown that wild mammals living in the Apuan Alps Regional Park in Tuscany are carriers of antibiotic-resistant enterococci, including strains of genetic lines characteristic for clinical pathogens. In our recent article, we have demonstrated that the wildlife from Tuscany are also a source of drug-resistant and pathogenic strains of Escherichia coli. 56 The intestinal microflora of animals inhabiting the park was not subjected to the natural selection pressure associated with administration of antimicrobial agents, but they may have acquired-resistance bacteria by contact with contaminants that can be transmitted to the wild from urbanized areas with water or migratory birds. The frequency of resistance in the enterococci studied was generally lower than that observed in the case of isolates from hospitalized patients or livestock, 16 but the occurrence of MDR strains, VRE, and virulence genes in the isolates tested may pose a threat to public health. Through their predatory and travelling habits, wild mammals (including wolves) may contribute to transmission of resistant bacteria and their genes between different ecosystems and, hence, they can be an important link in the development of antibiotic resistance.
Footnotes
Acknowledgments
The authors thank Fabio Viviani (Apuan Alps Regional Park, Italy) and Flavio Bresciani (Department of Veterinary Science, University of Pisa, Italy) for their assistance in collecting the fecal samples and Prof. Grzegorz Tomczyk (National Veterinary Research Institute in Puławy, Poland) for providing the DNA of the vanA-positive Enterococcus faecium ATCC 51559 reference strain.
Disclosure Statement
No competing financial interests exist.
Funding Information
All stages of research were financed entirely from the budget of the University of Life Sciences in Lublin, Poland.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.