
Abstract
Of all the Proteus spp., Proteus mirabilis is the most common species identified in clinical specimens and is a leading agent of complicated urinary tract infection. This study was undertaken to understand the antimicrobial susceptibility, prevalence of antibiotic resistance genes, and molecular typing of P. mirabilis isolates collected from three hospitals in northern Taiwan. The results showed that the collected isolates of P. mirabilis were susceptible to most antibiotics except cefazolin and tigecycline. Many resistance genes were detected in the collected isolates, of which TEM genes were the most common. Resistance to third- or fourth-generation cephalosporins was related to the presence of at least one of the tested extended-spectrum β-lactamase (ESBL) or AmpC genes. The presence of the VEB-1 gene seemed to be a good predictor for both cefepime and ceftazidime resistance, which was further supported by quantitative polymerase chain reaction results. Of the four imipenem-resistant P. mirabilis isolates, three isolates could hydrolyze imipenem by mass spectrometry analysis. Molecular typing by pulsed-field gel electrophoresis showed that the pulsotyping of the selected P. mirabilis isolates was heterogeneous. By analyzing the relationship of antimicrobial resistance and the presence of resistance genes, revision of the Clinical and Laboratory Standards Institute cefepime and ceftazidime MIC breakpoints for Enterobacteriaceae to predict ESBL producers might possibly be needed.
Introduction
Members of the genus Proteus are widely distributed in nature and can be isolated from soil, stagnant water, sewage, and the intestinal tract. Of all the Proteus spp., P. mirabilis is by far the most common species identified in clinical specimens and is a leading agent of pyelonephritis, urolithiasis, prostatitis, and catheter-associated urinary tract infections.1,2
Although P. mirabilis is naturally resistant to penicillin G, oxacillin, macrolides, lincosamides, streptogramins, glycopeptides, rifampicin, and fusidic acid, 3 this microorganism is still susceptible to many categories of antibiotics used in clinical practice. The data of SENTRY antimicrobial susceptibility program from the United States of America and European Union for isolates collected in 2009–2011 reported that <10% of isolates were resistant to amikacin, aztreonam, cefepime, ceftazidime, ceftriaxone, meropenem, and piperacillin/tazobactam. 4 However, resistance to ceftazidime, cefepime, or aztreonam has been reported previously. 5 Clinical isolates of P. mirabilis can produce a diversity of beta-lactamases, including CTX-M- and CMY-type as well as VEB-4 and inhibitor-resistant TEM-type beta-lactamases, which could be responsible for phenotypic resistance. 6 However, the Surveillance of Multicenter Antimicrobial Resistance in Taiwan (SMART) study suggested confirming phenotypic extended-spectrum β-lactamase (ESBL)-positive P. mirabilis with molecular analysis because 78.7% of ceftazidime-susceptible P. mirabilis isolates in the study were ESBL positive. 7
Although there have been several previous studies on P. mirabilis in Taiwan, the correlation between the presence of antibiotic resistance genes, including ESBL and carbapenemase genes, and antimicrobial susceptibility as well as molecular typing are still unclear. Therefore, this study was undertaken by collecting clinical isolates from three hospitals to understand the antimicrobial susceptibility and molecular epidemiology of P. mirabilis in Taiwan.
Materials and Methods
Bacterial strains
All clinical isolates of P. mirabilis were collected nonrepetitively from three hospitals, hospital A with 362 ward beds, hospital B with 693 ward beds, and hospital C with 504 ward beds, in northern Taiwan from November to December 2017. The collected isolates of P. mirabilis were stored at −80°C in trypticase soy broth (Difco Laboratories, Detroit, MI) supplemented with 20% glycerol before antimicrobial susceptibility tests (ASTs) and molecular biology experiments.
Antimicrobial susceptibility test
The identification and AST for all collected clinical isolates of P. mirabilis were performed in the microbiology laboratory of the National Taiwan University Hospital Chu-Tung branch using the BD Phoenix™ automated identification and susceptibility testing system and the method as previously described. 8 However, the susceptibility test for imipenem was performed by the disk diffusion method because the imipenem susceptibility report for P. mirabilis in the Phoenix system has not been approved by the Clinical and Laboratory Standards Institute (CLSI).
Detection of resistance genes by polymerase chain reaction
Detection of important β-lactamase genes in the collected isolates was performed by using a multiplex polymerase chain reaction (PCR) assay as previously described. 9 Group-specific primers used in this study are shown in Supplementary Table S1. Six multiplex PCRs were designed in this study: a blaTEM/blaSHV/blaOXA-1-like multiplex PCR; a blaCTX-M multiplex PCR, including phylogenetic groups 1, 2, and 9; a plasmid-mediated AmpC β-lactamase gene multiplex PCR, including six groups based on percentage of similarity; a blaVEB/blaGES/blaPER multiplex PCR; and two carbapenemase gene multiplex PCRs, one including the blaVIM, blaIMP, and blaKPC genes and the other including the blaGES and blaOXA-48-like genes. One simplex PCR was designed for blaCTX-M-8/-25. To completely identify the VEB amplicons in VEB-positive isolates, the amplicons were amplified by using the primers VEB-F 5′-GTTAGCGGTAATTTAACCAGATAG-3′ (position 3517–3540, accession no. AF205943) and VEB-B 5′-CGGTTTGGGCTATGGGCAG-3′ (position 4587–4569, accession no. AF205943) and then were subjected to DNA sequencing by an ABI 3730 XL DNA Analyzer (Applied Biosystems, Foster City, CA). 6 DNA sequences were compared to those registered in the National Center for Biotechnology Information database. After collecting the data of the AST and the presence of important β-lactamase genes, the correlation between the phenotype and genotype of antibiotic resistance was assessed.
Gene expression of blaVEB by quantitative PCR
Gene expression was analyzed by quantitative PCR (qPCR) as previously described with some modifications. 10 In brief, total RNA was isolated from 1 × 109 P. mirabilis cells, which were then treated with DNase and submitted for cDNA synthesis. Each 20-μL reaction mixture contained 25 ng of cDNA, 10 μL of Power SYBR Green PCR Master Mix (Life Technologies), and 300 nM each forward and reverse primer. The internal forward and reverse primers were designed for the VEB gene (CAAATGCACAAGGATTGGAA and AGTTTGTTCATCGCTGTTGG) by using the DesignStudio web-based tool (Illumina). The reactions were performed with 1 cycle at 95°C for 10 min followed by 40 cycles of 95°C for 15 sec and 60°C for 1 min. The 16S rRNA transcript was used as an endogenous control for the qPCR. The data were analyzed using the Echo Real-Time PCR System (Illumina). The experiments were repeated in triplicate independent experiments.
Detection of carbapenemase hydrolysis by using liquid chromatography-mass spectrometry/mass spectrometry
Each strain was analyzed using liquid chromatography-mass spectrometry/mass spectrometry (LC-MS/MS) to detect imipenem hydrolysis as previously described. 10 In brief, the strains were cultured overnight on Mueller-Hinton agar. The bacteria were suspended in a saline solution and adjusted to OD600 = 2.0. A 1-mL volume of this suspension was incubated with 5 μg/mL imipenem for 1 hr at 37°C with smooth agitation. The suspensions were subsequently centrifuged at 12,000 g for 5 min, and 300 μL of the supernatant was mixed with 700 μL of methanol. After centrifugation at 12,000 g for 5 min, 200 μL of the supernatant was mixed with 800 μL of water. The abundance of imipenem was measured through LC-MS/MS using the Thermo Accela LC system (Waltham, MA) coupled to a TSQ Quantum tandem triple-quadrupole mass spectrometer. Acinetobacter baumannii ATCC 17978 was used as a negative control, and A. baumannii ATCC 17978 IMP, an induced imipenem-resistant strain, was used as a positive control for imipenem hydrolysis.
Bacterial typing by pulsed-field gel electrophoresis
To decide which related isolates of P. mirabilis were pulsotyped by pulsed-field gel electrophoresis (PFGE), rep-PCR was performed first. Rep-PCR was performed according to the method as previously described. 11 PFGE was then performed on the genomic DNA of the selected related P. mirabilis isolates as previously reported. 12 The Rep-PCR and PFGE profiles of the P. mirabilis strains were analyzed using Numerical Taxonomy and Multivariate Analysis System version 2.0 (Applied Biostatistics, Inc., Setauket, NY). Cluster analysis for DNA fingerprints was performed by constructing a dendrogram using the bands-based dice coefficient method. A similarity matrix was generated and subsequently clustered using the unweighted pair group method with arithmetic means.
Results
Antimicrobial susceptibility test
A total of 101 P. mirabilis isolates, including 24, 38, and 39 isolates from hospitals A, B, and C in Taiwan, respectively, were collected from November to December 2017 (Table 1). More than 90% of the collected P. mirabilis isolates in this study were susceptible to amikacin, third generation cephalosporins, cefepime, piperacillin/tazobactam, and carbapenem antibiotics, whereas only 58% and 47% were susceptible to ciprofloxacin (levofloxacin) and trimethoprim/sulfamethoxazole (Table 2). Ninety-six percent of the collected isolates were not susceptible to cefazolin; in contrast, 100% were susceptible to cefmetazole. Only four isolates were resistant to imipenem by the disk diffusion method. However, the difference in imipenem susceptibility between the disk diffusion method and the Phoenix system was marked. In addition, all collected isolates were resistant to tigecycline.
Sources of the Proteus mirabilis Isolates Collected from Three Hospitals
Source: Dallenne et al., 2010. 9
One from a cervical swab and the other from a biopsy.
Antimicrobial Susceptibility of the Collected Proteus mirabilis Isolates
AMK, amikacin; CFZ, cefazolin; FEP, cefepime; CMZ, cefmetazole; CAZ, ceftazidime; CTX, cefotaxime; CIP, ciprofloxacin; ETP, ertapenem; GEN, gentamicin; LVX, levofloxacin; MEM, meropenem; TGC, tigecycline; SXT, trimethoprim/sulfamethoxazole; SAM, ampicillin/sulbactam; TZP, piperacillin/tazobactam; IPM, imipenem.
Detection of resistance genes by PCR
The results of resistance gene detection with multiplex PCR by seven group-specific primers are shown in Fig. 1. 9 Resistance genes detected by blaTEM/blaSHV/blaOXA-1-like multiplex PCR primers were the most prevalent. Forty-two P. mirabilis isolates possessed TEM genes, and seven had OXA genes. Resistance genes of blaCTX-M phylogenetic groups 1, 2, or 9 were detected in five isolates, with group 2 and 9 genes present in the same two isolates. Resistance genes detected by MultiCaseCIT primers for plasmid-mediated AmpC β-lactamase were present in 12 isolates. Five clinical isolates had VEB genes. However, no resistance genes were detected by the two carbapenemase gene multiplex PCRs.

Prevalence of resistance genes in the collected Proteus mirabilis isolates.
Correlation between antimicrobial susceptibility and the presence of resistance genes
Since resistant genes detected by MultiTSO-T and MulticaseCIT primers were the most prevalent in the collected P. mirabilis isolates, the relationship of antimicrobial susceptibility and the presence of these two groups of resistance genes was further examined.Figures 2 and 3 show that the presence of genes detected by MultiTSO-T (TEM genes) and MultiCaseCIT primers (CIT group genes) was related to higher antimicrobial resistance rates for most antibiotics. All of the amikacin-resistant P. mirabilis isolates harbored resistance genes in both the TEM and CIT groups. All of the cefepime-resistant and ceftazidime-resistant P. mirabilis isolates harbored resistance genes in the TEM group. Of the four imipenem-resistant P. mirabilis isolates, three were intermediate-resistant to meropenem (all harboring CIT group genes) and one was susceptible to meropenem (harboring a VEB gene).

Relationship of antimicrobial susceptibility and presence of TEM genes in the collected P. mirabilis isolates. AMK, amikacin; CFZ, cefazolin; FEP, cefepime; CMZ, cefmetazole; CAZ, ceftazidime; CTX, cefotaxime; CIP, ciprofloxacin; ETP, ertapenem; GEN, gentamicin; LVX, levofloxacin; MEM, meropenem; TGC, tigecycline; SXT, trimethoprim/sulfamethoxazole; SAM, ampicillin/sulbactam; TZP, piperacillin/tazobactam; IPM, imipenem.

Relationship of antimicrobial susceptibility and the presence of resistance genes detected by MulticaseCIT primers (CIT) in the collected P. mirabilis isolates.
In this study, only a few isolates were not susceptible to cefepime, ceftazidime, or cefotaxime. Table 3 shows the relationship of cefepime, ceftazidime, and cefotaxime resistance with the presence of ESBL genes (CTX-M or VEB) or AmpC genes (genes detected by MulticaseCIT primers, possibly CMY). Resistance to cefepime, ceftazidime, or cefotaxime was related to the presence of at least one of the listed ESBL or AmpC genes. All the five isolates carrying VEB genes were resistant to cefepime and ceftazidime with two of them still susceptible to cefotaxime.
Correlation of Cefepime, Ceftazidime, and Cefotaxime Resistance with the Presence of Extended-Spectrum β-Lactamase or AmpC Genes in Eleven Proteus mirabilis Isolates
AST, antimicrobial susceptibility test; ESBL, extended-spectrum β-lactamase; FEP, cefepime; CAZ, ceftazidime; CTX, cefotaxime.
Gene expression of blaVEB by qPCR
To clarify the VEB-like genes in the P. mirabilis isolates, the amplicons were sent for sequencing and demonstrated as VEB-1 (data not shown). The gene expression of VEB-1 in the five collected clinical isolates carrying VEB-1 genes was measured by qPCR, and no significant difference was found among these isolates. This provided further evidence that the VEB-1 gene was responsible for cefepime and ceftazidime resistance in these five VEB-positive P. mirabilis isolates.
Detection of carbapenemase hydrolysis by using LC-MS/MS
Table 4 and Supplementary Fig. S1 show the result of imipenem hydrolysis by the collected imipenem-resistant P. mirabilis isolates. The intensity increased and peaked at 3 and 3.5 min, implying that two isomers of imipenem were present in the mass spectrum. The peak value was the area summation under these two peaks. Imipenem could still be detected in the presence of the No. 41 P. mirabilis isolate, suggesting that this strain could not hydrolyze imipenem, whereas no imipenem could be detected in the presence of the No. 43, 59, and 92 P. mirabilis isolates, suggesting that these three strains could hydrolyze imipenem. The results suggested that either carbapenemase or other mechanisms, for example, outer membrane permeability change, could be responsible for imipenem resistance in P. mirabilis.
Detection of Imipenem Hydrolysis for Four Proteus mirabilis Isolates by Using Liquid Chromatography-Mass Spectrometry/Mass Spectrometry
Acinetobacter baumannii ATCC 17978, negative control; A. baumannii ATCC 17978 IMP, positive control.
N/A, not applicable.
Bacterial typing by PFGE
Molecular typing for the collected 101 P. mirabilis isolates was first performed with rep-PCR. The results are shown in Supplementary Fig. S2. The results revealed that all the isolates can be divided into 29 groups with similarity >80% being classified as the same type. The largest group contained 25 isolates, which was further analyzed by PFGE with five VEB-positive P. mirabilis isolates. However, two isolates had a smearing phenomenon on PFGE and were thus excluded for further analysis. Figure 4 shows that the pulsotyping of the 28 selected P. mirabilis isolates was heterogeneous and no predominant pulsotype could be identified. Based on the results of PFGE, no epidemic P. mirabilis strains could be found in this study, which implied that no horizontal transmission or outbreak caused by P. mirabilis occurred in the three hospitals during the study period.
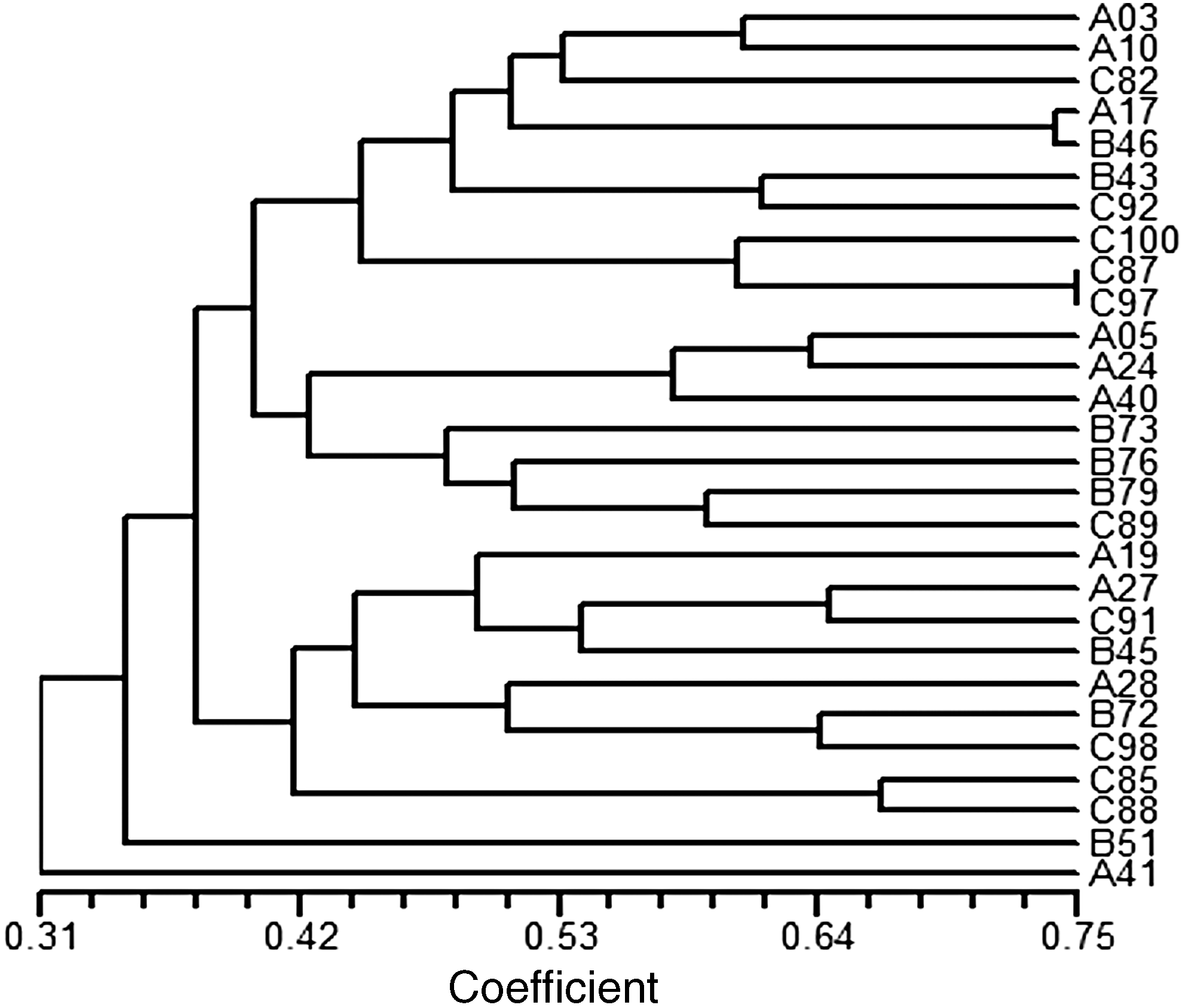
Phylogenetic tree of the 28 selected P. mirabilis isolates by pulsed-field gel electrophoresis.
Discussion
Proteus, which belongs to the family Enterobacteriaceae, is a genus of gram-negative, motile facultative anaerobic rods. The genus Proteus currently consists of five named species: P. mirabilis, P. vulgaris, P. penneri, P. myxofaciens, and P. hauseri and three unnamed genomospecies: Proteus genomospecies 4, 5, and 6. 13 Of all the Proteus spp., P. mirabilis is the most common species identified in clinical practice. The clinical manifestations of infections with Proteus spp. are nonspecific. However, urinary tract infections involving struvite stones are characteristic, which has been ascribed to urease activity. 2
The Taiwan surveillance of antimicrobial resistance (TSAR) data from period III (2002) to VIII (2012) showed that susceptibility to cefotaxime, ceftazidime, and ciprofloxacin decreased significantly during the past decade, from 92.6% to 81.7%, 100% to 95.2%, and 80.1% to 53.8%, respectively (p < 0.01). 14 In this study, the rates of susceptibility to cefotaxime, ceftazidime, and ciprofloxacin were 91%, 93%, and 58%, respectively. It seemed that the antimicrobial susceptibility of P. mirabilis to third generation cephalosporins and quinolones did not change substantially 5 years after the TSAR program. However, the susceptibility rate of cefazolin decreased markedly during the last 5 years from 81.2% of the urine isolates in the TSAR program to only 4% of the collected isolates in this study. These results implied that cefazolin can no longer be used as an empirical therapy for infections caused by P. mirabilis in Taiwan.
Synthesis of β-lactamases is the predominant mechanism for resistance to β-lactam antibiotics. 15 In the last decade, there have also been numerous reports of the production of ESBLs by P. mirabilis.5,7,14,16,17 The prevalence of ESBLs varies in different studies, for example, 3.6% in the 2014 United States SMART study and 8.2% in the 2014 TSAR program.14,17 Different types of ESBLs in P. mirabilis have been detected previously. CTX-M-type ESBLs have been detected in P. mirabilis isolates from Taiwan, Korea, Japan, and Israel.14,18–20 CTX-M2 is the most common ESBL in Japan and appears to be spreading rapidly. 21 In addition, CTX-M has been found on the P. mirabilis chromosome as part of an integrative and conjugative element in addition to being plasmid-encoded. 22 Integron-located VEB-1 ESBL has been detected in a P. mirabilis clinical isolate from Vietnam. 23 Plasmid encoding VEB-1 in P. mirabilis has been reported in Greece. 24 The prevalence and kind of ESBLs in this study were similar to the previous ones, with 10.9% (11 isolates in Table 3) of 101 P. mirabilis isolates harboring ESBLs, CTX-M, and VEB-1.
TEM genes among the ESBL-producing P. mirabilis isolates, including TEM-52 as well as TEM 92 and TEM 177, have been detected in Croatia and Israel, respectively.16,20,25 Although TEM was the most common resistance gene in this study, only TEM-1 and TEM-2 were detected by the primers we used. To detect ESBL TEM genes, the primers should be redesigned in future studies. The first P. mirabilis isolate with PER-1 was from Italy, whereas VIM-1 and SHV were detected in ESBL-positive P. mirabilis isolates from Bulgaria and South Africa, respectively.26–28 However, PER-1, VIM-1, and SHV were not detected in the collected isolates of P. mirabilis in this study.
Another important mechanism of resistance to extended-spectrum cephalosporins in P. mirabilis isolates is the presence of plasmid-mediated AmpC β-lactamases. 28 AmpC type β-lactamases can either be chromosomally encoded or plasmid encoded in P. mirabilis.29,30 Strains with AmpC β-lactamases are resistant to aminopenicillins, carboxypenicillins, ureidopenicillins, and third generation cephalosporins. AmpC β-lactamases generally do not effectively hydrolyze cefepime or carbapenems. CMY has been regarded as the most common type of AmpC β-lactamase in P. mirabilis isolates from Taiwan, Korea, and Spain.14,18,29 The clonal spread of CMY-producing P. mirabilis has been reported in Europe. 31 Genes detected by MulticaseCIT primers in this study included LAT-1 to LAT-3, BIL-1, CMY-2 to CMY-7, CMY-12 to CMY-18, and CMY-21 to CMY-23. Therefore, we speculated that CMY was also possibly the most common type of AmpC β-lactamase in the collected isolates of P. mirabilis.
A previous study performed in Huashan Hospital, China showed that all the collected P. mirabilis were resistant to cefazolin, cefotaxime, and ceftriaxone, while 85.0–96.7% of these strains were susceptible to ceftazidime, cefepime, and aztreonam. 5 Because the cefepime minimal inhibitory concentration (MIC) breakpoints defined by the CLSI and the European Committee on Antimicrobial Susceptibility Testing were poor at predicting ESBL producers, revision of the cefepime MIC breakpoint for Enterobacteriaceae was suggested. 32 In this study, although resistance to cefepime, ceftazidime, or cefotaxime was related to the presence of at least one of the listed ESBL (CTX-M or VEB) or AmpC genes (possibly CMY), some cefepime- or ceftazidime-susceptible P. mirabilis isolates were still ESBL positive (Table 3). Therefore, revision of the CLSI cefepime and ceftazidime MIC breakpoints for Enterobacteriaceae to predict ESBL producers might possibly be needed. However, more P. mirabilis strains must be examined in the future to confirm this suggestion.
The MYSTIC study in 2010 indicated that despite continued use of meropenem, carbapenem resistance is not increasing among Enterobacteriaceae tested. It concluded that clinicians can still administer carbapenems as a reliable and effective choice in managing serious nosocomial infections. 33 The conclusion was consistent with the findings of our study, which revealed that no isolates were highly resistant to meropenem and only four isolates were resistant to imipenem. However, a study from Taiwan found that only 11.4% of P. mirabilis isolates were susceptible to imipenem with the broth microdilution test, although all the tested isolates were still susceptible to meropenem. 34 We do not know whether the high imipenem resistance rate in the Tsai et al. 34 study was determined by an automatic system or due to high exposure of imipenem. In June 2010, the CLSI published new MIC and disk diffusion interpretive breakpoints for Enterobacteriaceae for three carbapenems (imipenem, meropenem, and ertapenem). 35 After implementation of the newer interpretive criteria (M100-S20-U, June 2010), the Modified Hodge test does not need to be performed other than for epidemiological or infection control purposes. Another change in CLSI recommendations occurred in 2012, and SENTRY data from North America indicated that this change decreased the level of imipenem susceptibility compared to the 2010 criteria (64.5% of 1,244 isolates were susceptible by 2012 criteria vs. 99.8% by 2010 criteria). 36 Furthermore, the 2018 CLSI guidelines suggested that imipenem MICs for Proteus spp., Providencia spp., and Morganella morganii tend to be higher than meropenem or doripenem MICs; thus, all automated systems have not been able to report imipenem MIC results. 37 The other evaluated method should be performed due to clinical requirements.
Carbapenemases have been found in P. mirabilis. 38 One report has documented the presence of the class D carbapenemase, OXA-23, in P. mirabilis. 39 Metallo-beta-lactamases were also reported in P. mirabilis isolates. 40 Although no carbapenemase genes could be detected in this study, the result of mass spectrometry showed hydrolysis of imipenem was the main mechanism for imipenem resistance of the collected P. mirabilis isolates. To determine which carbapenemase is responsible for imipenem hydrolysis, other primers should be designed in future studies. Further studies are needed to explore the possible mechanism contributing to imipenem resistance in P. mirabilis besides carbapenemases.
In conclusion, this study not only demonstrated the correlation between the presence of resistant genes and antimicrobial resistance but also implied other possible mechanisms besides carbapenemase for imipenem resistance in P. mirabilis. By analyzing antimicrobial susceptibility data of the clinical P. mirabilis isolates, this study may help clinicians in Taiwan choose empirical antibiotic treatment more appropriately.
Footnotes
Acknowledgments
This study was supported by National Taiwan University Hospital Chu-Tung Branch (grant number 107002). We are grateful to Professor Kai-Chih Chang and Professor Anren Hu for technical support in PFGE and mass spectrometry. We also thank Ms. Chu-Hwa Ten, Ms. Shu-Ling Huang, Ms. Fang-Yu Hsu, Ms. Cheng-Yu Yen, and Ms. Tai-Lin Lin for technical assistance.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.