
Abstract
KPC-producing Klebsiella pneumoniae (KPC-Kp) has become an important public health issue. The previous intestinal colonization by KPC-Kp has been an important risk factor associated with the progression to infections. The objective of this study was to assess the genetic characterization of KPC-Kp isolates recovered from human rectal swabs in Brazil. We selected 102 KPC-Kp isolates collected during 2009–2013 in 11 states. Antimicrobial susceptibility was determined by disk diffusion, E-test, and broth microdilution. The resistance and virulence genes were investigated by PCR. Molecular typing was performed by pulsed-field gel electrophoresis (PFGE) and multilocus sequence typing (MLST). The isolates were mostly resistant to β-lactams, sulfonamides, chloramphenicol, quinolones, and aminoglycosides but susceptible to fosfomycin/trometamol, polymyxin B, and tigecycline. The blaKPC-2 was mostly associated with Tn4401b. Besides that, the isolates carried blaCTX-M, blaSHV, blaTEM, and aac(6′)-Ib in high frequency and aac(3′)IIa and qnr genes in moderate frequency. The PFGE revealed 26 pulsotypes and MLST performed in representative strains revealed 23 sequence types, 45% belonging to clonal complex 258 (CC258). Isolates of CC258 were found in all states. Seventy percent of the 102 KPC-Kp isolates belonged to CC258-associated pulsotypes. We describe the dissemination of KPC-2-Kp associated with Tn4401b belonging to CC258 colonizing patients in Brazil, which is also prevalent in infected patients, suggesting a clear colonization–infection correlation.
Introduction
The globally widespread Klebsiella pneumoniae carbapenemase (KPC)-producing K. pneumoniae (KPC-Kp) has become one of the most important issues in public health. 1 KPC-Kp is often resistant to most β-lactams and other antimicrobial agents 2 and high rates of morbidity and mortality usually resulting in infections caused by these pathogens. 1 The KPC coding gene (blaKPC-2) has a high capacity to spread because it is mostly associated with transposons such as Tn4401, usually carried on transferable plasmids. 3
The expansion of KPC-Kp clonal lineages belonging to clonal complex 258 (CC258) plays a major role in the spread of carbapenem resistance worldwide. 1 The knowledge of the epidemiologic, phenotypic, and molecular features of KPC-producing strains became fundamental in understanding the resistance panorama allowing measures to control dissemination. 2 In Brazil, KPC-Kp has an endemicity status 1 and molecular studies describe the prevalence of KPC-Kp isolates belonging to CC258.4,5
It is important to note that most studies focus on isolates collected from clinical samples of infected patients with limitation of isolates from colonization surveillance culture. It is also relevant that previous intestinal colonization by KPC-Kp is one of the most important risk factors associated with the progression to extraintestinal nosocomial KPC-Kp infections. 6 Furthermore, colonized patients may serve as a reservoir for the spread of KPC-Kp in health care facilities. 1
Therefore, the characterization KPC-Kp from colonization may provide insights to understand the progression of colonization to infection. 7 In this regard, this study aims to assess the microbiological and genetic features in a KPC-Kp population collected from the gut environment of colonized patients to continue monitoring the evolution of KPC-Kp in Brazil.
Materials and Methods
Bacterial isolation and identification
As part of the Bacterial Nosocomial Infection Resistance Surveillance network, the Laboratório de Pesquisa em Infecção Hospitalar (LAPIH), located at Oswaldo Cruz Institute (Rio de Janeiro, Brazil), routinely receives clinical bacterial isolates from Brazilian health care services and state public health laboratories to perform multiplex PCR for investigation of genes currently associated with carbapenem resistance in Enterobacteriaceae isolates (blaKPC, blaNDM, and blaOXA-48-like). The isolates were previously identified by automated systems in the laboratories where they were first collected and then reconfirmed by conventional biochemical techniques in LAPIH. All isolates belong to the Culture Collection of Hospital-Acquired Bacteria (CCBH) being stored at −70°C in BHI broth (20% glycerol).
This study included 102 nonduplicated KPC-Kp isolates collected from rectal swabs of hospitalized patients from 2009 to 2013 in 11 Brazilian states, comprising four different geographical regions of this country.
Antimicrobial susceptibility testing
Antimicrobial susceptibility tests of all isolates were evaluated using the disk diffusion method to amikacin 30 μg (AMK), aztreonam 30 μg (ATM), cefepime 30 μg (FEP), cefotaxime 30 μg (CTX), cefoxitin 30 μg (FOX), ceftazidime 30 μg (CAZ), cephalothin 30 μg (CEF) chloramphenicol 30 μg (CHL), ciprofloxacin 5 μg (CIP), ertapenem 10 μg (ETP), fosfomycin/trometamol 200 μg (FOT), gentamicin 10 μg (GEN), levofloxacin 5 μg (LVX), piperacillin/tazobactam 110 μg (PTZ), and sulfamethoxazole/trimethoprim 25 μg (SXT) (Oxoid, Basingstoke, United Kingdom). The minimum inhibitory concentrations (MIC) were determined using the E-test (AB Biodisk, Sweden) for imipenem (IPM), meropenem (MEM), and tigecycline (TGC). For polymyxin B (PMB), we first screened with E-test and then confirmed the PMB-resistant strains with cation-adjusted Mueller–Hinton broth microdilution. The results were interpreted according to the Clinical and Laboratory Standards Institute (CLSI) breakpoints 8 except for PMB and TGC, for which the European Committee for Antimicrobial Susceptibility Testing (EUCAST) 9 criteria were used. Phenotypic detection of carbapenemases was based on the use of phenylboronic acid and EDTA as enzymatic blockers. 10 Escherichia coli ATCC®25922 was used as a control for antimicrobial susceptibility testing.
The characterization of the resistance patterns for Enterobacteriaceae used to define multidrug resistance (MDR); extensively drug resistance (XDR), and pandrug resistance (PDR) was performed according to Magiorakos et al. 11
Genetic relatedness and genotyping
Genetic similarities among all the KPC-Kp isolates were investigated by pulsed-field gel electrophoresis (PFGE). The DNA was digested with XbaI, and the DNA fragments were separated by 1% agarose gel electrophoresis for 17 hrs using a CHEF-DR III apparatus (Bio-Rad), with pulses varying from 0.5 to 35 sec at a voltage of 6 V/cm. After staining with ethidium bromide (0.5 μg/mL), the gels were examined using BioNumerics v.6.6 (Applied Maths, Belgium). Similarity among the isolates was estimated using the Dice coefficient with 1.5% optimization and a 1.0% tolerance setting. The PFGE pulsotypes were defined as DNA patterns sharing ≥80% similarity and named by a capital letter.
Representative isolates were randomly selected among distinct pulsotypes for each studied state to perform multilocus sequence typing (MLST) according to the protocol of the Institut Pasteur. New alleles and sequence types (STs) were submitted to the MLST website for approval.
Molecular investigations of antimicrobial resistance
All KPC-Kp isolates were screened by PCR for the presence of other carbapenemase genes (blaNDM, blaOXA-48-like, blaIMP, and blaVIM) 12 and mcr-1 gene. 13 For the representative isolates we also performed PCR assays to detect the β-lactamase blaCTX-M, blaSHV, and blaTEM 14 encoding genes; plasmid-mediated quinolone and aminoglycoside resistance determinants qnrA, qnrB, qnrS, aac(3′)IIa, aac(6′)-Ib, 14 armA, rmtA, rmtB, rmtD, rmtG, and npmA. 15 The PCR amplicons of blaKPC 16 and blaCTX-M from the representative isolates were sequenced using Sanger methodology and analyzed using Geneious v.6.1.8 (Biomatters Ltd., New Zealand) and the National Center for Biotechnology Information (NCBI) website with BLASTn tool.
Detection of flanking regions of blaKPC
The genetic environment of the blaKPC gene was analyzed for the representative isolates by multiplex PCR according to Naas et al. 17 except for the amplification of the ISKpn7 region, in which the primers described by Kitchel et al. 3 were adopted. The Tn4401 isoform was determined by the sequencing of ISKpn7 amplicon.
Detection of virulence-associated features
For the representative isolates, the virulence factor-encoding genes cf29a, ycfM, mrkD, fimH, (adhesion), entB, iroN, kfu, ybtS (siderophores), magA (hypermucoviscous phenotype), and allS (allantoin metabolism) were searched by PCR 14 and the hypermucoviscosity (HMV) phenotype was searched by the string test. 14
Results
The 102 isolates were recovered from hospitals located in the states of Rio de Janeiro (n = 12), Espírito Santo (n = 11), and Minas Gerais (n = 14) in southeastern Brazil; Goiás (n = 5), Federal District (n = 14) and Mato Grosso (n = 1) in midwestern Brazil; Maranhão (n = 5), Paraíba (n = 2), and Pernambuco (n = 14) in northeastern Brazil; Santa Catarina (n = 14) and Rio Grande do Sul (n = 10) in southern Brazil. No isolates from the northern region were received by LAPIH during the study period. We included isolates from the years 2009 (n = 1), 2010 (n = 12), 2011 (n = 51), 2012 (n = 9), and 2013 (n = 29).
All KPC-Kp isolates were positive for phenotypic carbapenemase production and no other carbapenemase was detected among the investigated genes. Most of isolates were nonsusceptible to all β-lactams tested (95%), CIP (95%), SXT (90%), CHL (78%), and GEN (62%), but exhibited susceptibility to PMB (92%) and FOT (88%), being half of isolates susceptible to AMK (52%) and TGC (50%) as given in Fig. 1. About 78% (n = 80) isolates was considered MDR, 21% (n = 21) were XDR, and 1% (n = 1) PDR.

Antimicrobial susceptibility profile of KPC-Kp isolates. CEF, cephalothin; FOX, cefoxitin; TZP, piperacillin–tazobactam; CTX, cefotaxime; CAZ, ceftazidime; FEP, cefepime; ATM, aztreonam; ETP, ertapenem; MEM, meropenem; IPM, imipenem; GEN, gentamicin; AMK, amikacin; CIP, ciprofloxacin; LVX, levofloxacin; SXT, trimethoprim/sulfamethoxazole; CHL, chloramphenicol; FOT, fosfomycin/trometamol; PMB, polymyxin B; TGC, tigecycline.
A total of 26 PFGE pulsotypes (A to Z) was observed among the 102 KPC-Kp (Fig. 2). The MLST was performed in 40 randomly selected isolates representing each distinct PFGE pulsotypes by each state, which revealed 23 STs as given in Table 1 (ST11, ST13, ST16, ST17, ST36, ST37, ST101, ST147, ST198, ST273, ST283, ST327, ST336, ST340, ST437, ST442, ST514, ST736, ST855, ST864, ST874, ST1422, and ST2277). The ST2277 was first described in this study. A total of 18 of 40 (45%) of the representative isolates belonged to CC258, which was detected in all studied states during all studied years (Fig. 3). The CC258 comprised 70% of all the 102 isolates (ST437, n = 28; ST11, n = 26; ST340, n = 10; ST855, n = 7; and ST864, n = 1). Despite the clonal diversity observed, the most prevalent profile was the A/ST437 clone (28 isolates). Among the isolates considered XDR and PDR, 68% (n = 15/102) belonged to CC258.

Dendrogram and genetic features of representative KPC-Kp of the study. DF, distrito federal; ES, Espírito Santo; GO, Goiás; MA, Maranhão; MG, Minas Gerais; MT, Mato Grosso; PB, Paraíba; PE, Pernambuco; RJ, Rio de Janeiro; RS, Rio Grande do Sul; SC, Santa Catarina; HMV, hypermucoviscosity; ST, sequence type.
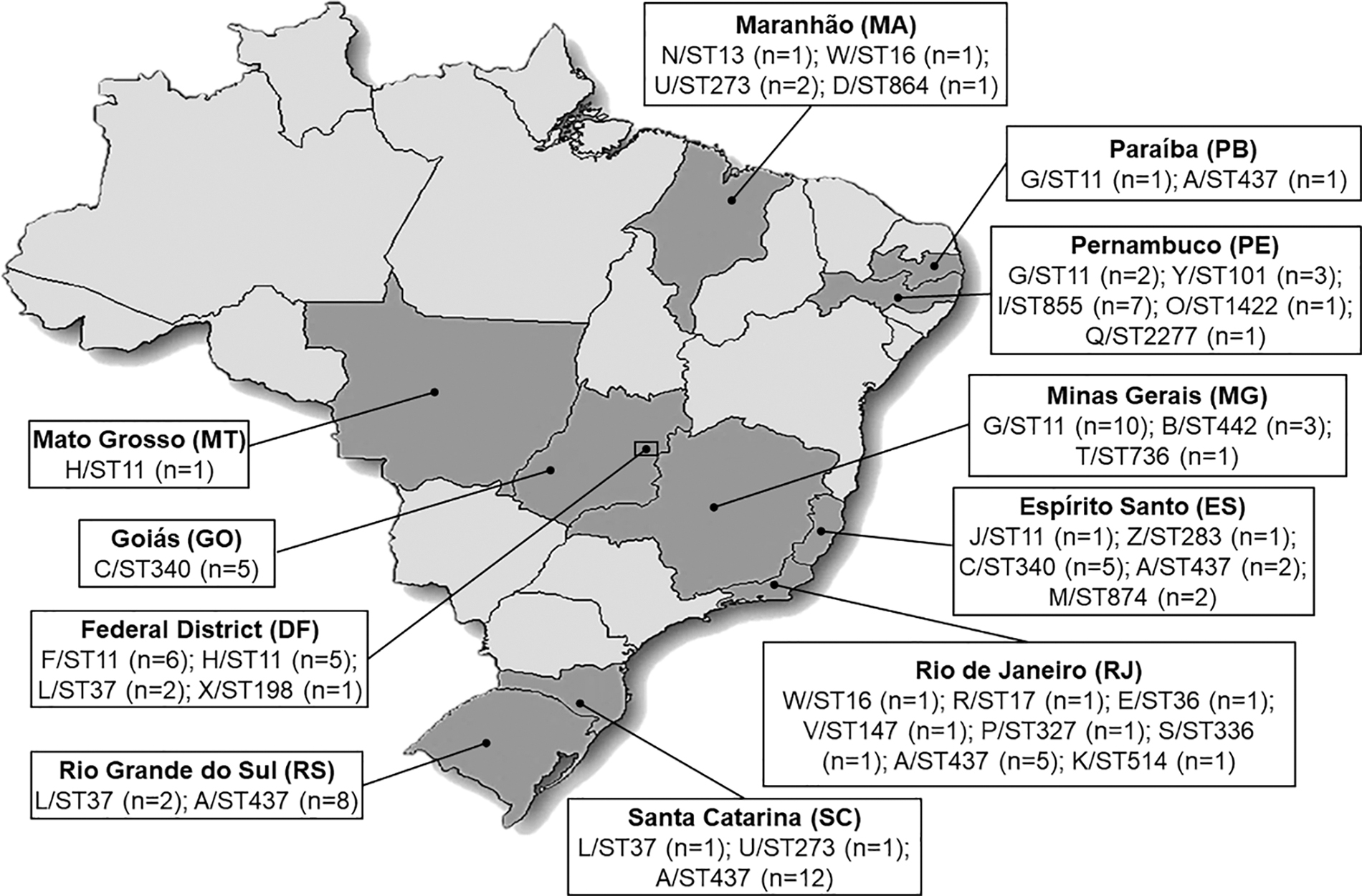
Distribution of the sequence types according to the isolation state of 102 KPC-Kp from Brazil.
Proportion of All Studied Isolates for Each Sequence Type
DF, distrito federal; ES, Espírito Santo; GO, Goiás; MA, Maranhão; MG, Minas Gerais; MT, Mato Grosso; PB, Paraíba; PE, Pernambuco; RJ, Rio de Janeiro; RS, Rio Grande do Sul; SC, Santa Catarina; ST, sequence type.
In the 40 representative isolates, the blaKPC-2 allele was the only variant found of the blaKPC gene and other genes associated with β-lactamases, resistance to fluoroquinolones and aminoglycosides were detected (Fig. 2). Most of the isolates harbored blaSHV (100%), blaTEM (92.5%), blaCTX-M (75%), and aac(6′)-Ib (67.5%) genes. The variants blaCTX-M-15 (53%) and blaCTX-M-2 (33%) were the most prevalent, whereas blaCTX-M-8 (5%), blaCTX-M-9 (2.5%), and blaCTX-M-14 (2.5%) were the least detected. The qnrB variant was the most prevalent (30%) of the investigated qnr genes, whereas only one isolate was positive for rmtG among the studied 16S rRNA methyltransferases. The average of additional resistance genes in isolates belonging to CC258 was X̅ = 4.5 genes/isolate, whereas in the non-CC258 clones it was X̅ = 3.8 genes/isolate.
The research for virulence determinants identified mrkD (97.5%), fimH (100%), ycfM (100%), kfu (15%), ybtS (87.5%), and entB (100%) genes and positive HMV phenotype (7.5%). The other investigated genes were not detected (Fig. 2). The average of virulence determinants was similar between CC258 and non-CC258 isolates (approximately five genes/isolate).
The blaKPC-2 gene was associated with Tn4401b in 76% of representative strains. No isolates amplified the inverted repeat sequences (IRS) of the flanking region of Tn4401. Amplification of only TnpA+ISKpn6, ISKpn6+ISKpn7, and ISKpn6 was detected in 15% (n = 6), 7.5% (n = 3), and 5% (n = 2) of representatives, respectively. Besides that, one of the isolates presented a deletion in ISKpn7 region compatible with Tn4401 isoform “d” (Fig. 2).
Discussion
Most of the Brazilian studies that assessed the genotype of KPC-Kp by MLST have focused on bacteria collected from infection sites.5,18–22 Data assessing genotyping of KPC-Kp from colonized patients is very limited in Brazil with restricted isolate numbers.4,23–26 Therefore, this study intended to analyze the population structure of the KPC-Kp collected by rectal swabs from different states in Brazil to point characteristics that will allow understanding the dynamics of dissemination and a possible progression from colonization to infection. We found 23 STs with a prevalence of those from CC258, mainly ST437, ST11, ST340, and ST855, respectively. Other studies with KPC-Kp isolated from gut colonization also detected CC258 isolates as ST11 and ST437, but other STs as ST25, ST29, ST48, ST258, ST392, ST1046, and ST33734,23–26 showing a high clonal diversity in this population.
In Brazil, the ST11, ST340, and ST437 are the predominant clones.4,5,23 These STs belong to CC258 being considered single-locus variants of ST258, a successful high-risk clone disseminated worldwide. 1 In our study, the ST258 was not detected, but the surveillance of this clone deserves attention. There have been few reports of the ST258 in specific locations in Brazil such as Londrina in Paraná (PR), 19 Porto Alegre in RS state, 21 and two cities of São Paulo (SP) state. 23 However, the dissemination of this clone still seems to be limited to some hospitals.
Comparing our study with other studies with KPC-Kp collected from infections,5,18–22 a clear correlation is observed regarding the prevalence of CC258 among the isolates of these two populations. In addition, we found other important non-CC258-related STs such as ST13, ST16, ST17, ST37, ST101, ST147, ST273, and ST442 that are frequently found in KPC-Kp from infected patients in Brazil.4,5,19–22 Our results are consistent with the findings of Baraniak et al. that showed that the endemic clones in European countries and Israel that predominated in clinical infection samples were the same clones that colonized patients. 27 Thus, this analysis provided a sample overview of the KPC-Kp clones distributed in some regions of Brazil, which reflects the endemic situation in this country, suggesting an important role of the high-risk clones colonizing patients and probably causing subsequent infections.
The clustering of PFGE analysis was consistent with the clones indicated by MLST, except for ST11, which were grouped in four pulsotypes (F, G, H, and J). This phenomenon of intraclonal variability is commonly described in ST11,20,22 showing the diversity of this clone.
Most of the isolates in this study were MDR (78%). However, a considerable number of XDR isolates were found (21%). A high level of resistance to different antimicrobial classes has frequently been observed in KPC-producing isolates worldwide. 2 Several Brazilian studies indicate that most of KPC-Kp, including clinical and colonization origins, are MDR with high resistance to all β-lactams classes and fluoroquinolones. For GEN and AMK, the percentage is variable, tending to be more resistant to GEN and more susceptible to AMK. High resistance is also observed for CHL and SXT when tested, whereas polymyxins, TGC, and FOT were shown to be the most effective agents.5,18,22,23,26,28 This pattern was also detected in this study.
We observed increased resistance rates for TGC compared with other studies.5,18,28 Resistance to TGC and polymyxins is very worrisome because these antimicrobial agents have been the last options for the treatment of severe infections caused by KPC-producing organisms. 2 A recent study with KPC-Kp from ICU in Tocantins (northern Brazil) also detected high resistance to TGC in KPC-Kp collected from rectal swabs with a significant statistical difference compared with other sources, suggesting an association between a pathogen from a specific source and an antimicrobial resistance pattern. 26 In a Greek study, high rates of colistin-resistant KPC-Kp (24.4%) and TGC-resistant KPC-Kp (17.9%) were found colonizing ICU patients during their stay. Multivariate analysis revealed that the administration of these antimicrobial agents and the number of colonized patients in nearby beds were significant risk factors associated with colonization by these pathogens. 29
In this study, 8% of the isolates were resistant to PMB because of alterations in the chromosomal mgrB gene reported in a previous publication. 13 This low percentage is consistent with studies that collected KPC-Kp in the same period.5,28 Despite this, some studies with KPC-Kp collected in periods after this study, have demonstrated an increase in polymyxin resistance,18,26,30 probably associated with overuse and misuse.26,30 No isolate from this study showed the plasmid-mediated colistin resistance mcr-1 gene. However, we recently detected a clinical KPC-Kp in Brazil carrying this gene, 31 further aggravating the scenario of antimicrobial resistance because this gene has a high dissemination potential.
The urgent expansion of XDR and PDR KPC-Kp strains reported in Brazil 32 has proved to be a major challenge for the medical community because of the limited treatment options. The PDR isolate present in this study was previously analyzed regarding the genomic characteristics 33 showing several acquired genes and mutations associated with antimicrobial resistance, highlighting the need for identification of such high-risk strains.
The blaKPC is predominantly associated with the transposon Tn4401, which is a Tn3-type active transposon of ∼10 kb in size bracketed by IRS. This structure consists of a transposase gene, a resolvase gene, the blaKPC gene, and two insertion sequences, ISKpn6 and ISKpn7.2,3 At present, nine isoforms of Tn4401 (Tn4401a to Tn4401i) were identified, differing by the upstream sequences of the blaKPC gene. 34
In this study, Tn4401b was prevalent among the isolates without the original IRS of the flanking region. This structure is commonly detected in several countries, including Brazil.2,3,5 However, in other isoforms, the loss of regions of this genetic element and other structures commonly described as non-Tn4401, were found in 13 of 40 isolates suggesting the participation of other elements in blaKPC spread, as shown by other studies in Brazil.23,34,35
The presence of other β-lactamase encoding genes, plasmid-mediated quinolone genes, and aminoglycoside resistance determinants detected in this study are described as commonly associated with KPC-Kp.20,23,24,32 We detected mainly blaSHV, blaTEM, blaCTX-M, being blaCTX-M-15 and blaCTX-M-2 the most prevalent variants, as already documented.20,22,24 In addition, we found that the aac(3′)IIa, aac(6′)-Ib, and qnrB genes seem to be very closely associated with KPC-Kp from CC258.22,36,37 These same genes are also present in non-CC258 isolates, reflecting a higher nonsusceptibility to aminoglycosides and fluoroquinolones throughout the studied KPC-Kp population.
Regarding the 16S rRNA methyltransferases, its coding genes seem to be detected at high frequency in KPC-Kp from China, mainly rmtB.36,37 We found only one ST340 isolate carrying the rmtG gene. The RmtG is an enzyme described for the first time in Brazil, that confers high resistance to aminoglycosides and was detected in Kp-producing-KPC and/or CTX-M from SP, including an ST340 isolate that also produced CTX-M-15, 24 similar to our study.
Some authors have reinforced the importance of active surveillance studies in addressing hospital-associated MDR K. pneumoniae clones. These studies should not only focus on the clonal origin and antimicrobial resistance but also on the virulence context to provide advantages concerning this global threat.25,38 In this study, we found a high-frequency fimH, entB, mrkD, entB, and ybtS, and a low frequency of the kfu gene and the HMV phenotype. This pattern of virulence factors has been reported in several studies including those of populations with different antimicrobial resistance profiles, clones, and sources19,20,25,39,40 showing no correlation among virulence factors, carbapenem resistance, and infections types.39,40
It is worth mentioning that the ybtS gene presented a much higher frequency in the KPC-Kp population from the colonization of our study compared with other Brazilian studies with KPC-Kp populations from infection sources.19,20,25 This fact is supported by Ferreira et al. that detected the presence of ybtS in 60% of a population composed of 56% of isolates collected from rectal swabs. 26 This finding corroborates the results of Cerdeira et al., which indicated that the presence of yersinbactin (ybtS) may be contributing to the emergence and persistence of highly virulent lineages of CC258 KPC-Kp in South America. 38 However, this finding requires further investigation.
Conclusions
In summary, this study characterized KPC-Kp isolates from colonized patients as multidrug resistant and belonging to CC258. The highlighted population profile is also the prevalent isolate profile commonly collected from infected patients, suggesting a clear colonization–infection correlation. Thus, we highlight the importance of surveillance primarily for the high-risk clones that serve as reservoirs of a wide variety of resistance determinants with the potential for progression from colonization to infection.
Ethical Approval
This study has ethical approval given by CEP/Fiocruz with reference number 167/02.
Footnotes
Acknowledgment
The authors thank CAPES (Brasil Sem Miséria/Brazilian governmental program) and the Instituto Oswaldo Cruz/FIOCRUZ–Brazilian Ministry of Health for the support, and the PDTIS-IOC DNA Sequencing Platform for DNA sequencing.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by CAPES (Brasil Sem Miséria/Brazilian governmental program fellowship to (C.A.M.A.), the Instituto Oswaldo Cruz/FIOCRUZ–Brazilian Ministry of Health (PAPES), CNPq, and FAPERJ.