
Abstract
Acinetobacter baumannii and Pseudomonas aeruginosa are the most relevant Gram-negative bacteria associated with hospital and opportunistic infections. This study aimed to evaluate the dynamics of drug-resistant A. baumannii and P. aeruginosa and biofilm formers from two public hospitals in northeastern Brazil. One hundred isolates (35 from A. baumannii and 65 from P. aeruginosa) were identified using the automated Vitek®2 Compact method (bioMérieux) and confirmed using the MALDI-TOF (MS) mass spectrometry technique. Molecular experiments were performed by polymerase chain reaction (PCR) to detect the frequency of blaKPC, blaIMP, blaVIM, and blaSHV genes. The biofilm formation potential was evaluated using crystal violet in Luria Bertani Miller and trypticase soy broth culture media under the following conditions: at standard concentration, one quarter (25%) of the standard concentration and supplemented with 1% glucose. In addition, the genetic diversity of the isolates was verified by the ERIC-PCR technique. Isolates presented distinct resistance profiles with a high level of beta-lactam resistance. The highest index of genes detected was blaKPC (60%), followed by blaSHV (39%), blaVIM (8%), and blaIMP (1%). All the isolates were sensitive to the polymyxins tested and formed biofilms at different intensities. Twelve clones of A. baumannii and eight of P. aeruginosa were identified, of which few were indicative of intra- and interhospital dissemination. This study reveals the dispersion dynamics of these isolates in the hospital environment. The results demonstrate the importance of monitoring programs to combat the spread of these pathogens.
Introduction
Infections caused by Gram-negative bacteria are among the leading causes of death worldwide. 1 The development of antibiotic therapies represented a major advancement in the treatment of these diseases. However, the extensive and indiscriminate use of antimicrobial drugs has led to the emergence of multidrug-resistant (MDR) bacterial strains and has rendered several antibiotics ineffective 2 ; thereby, becoming a clinical challenge that involves high cost to the health systems.3,4 Presently, an exponential increase in the number of MDR and extensive drug-resistant (XDR) strains is reported for several microbial species. 5
Microbial resistance is observed against almost all antibiotics that are frequently used in hospitals; however, an abrupt increase in resistance has been observed mainly against the β-lactam antibiotics. 6 This phenomenon can be explained by the widespread occurrence of the extended-spectrum β-lactamases (ESBLs), a diverse group of enzymes encoded by genes that provide resistance to the third- and fourth-generation cephalosporins and monobactams. Among these enzymes, the SHV-1 type is considered the most prevalent, comprising >100 varieties found in a wide range of species worldwide.7,8 In addition, enzymes such as carbapenemases have been described, which are capable of hydrolyzing all β-lactams. The increase in bacterial resistance is mainly due to the production of these enzymes that are derived from punctual mutations.9,10
The resistance to β-lactams, in particular, is evident in Gram-negative bacilli such as A. baumannii and P. aeruginosa, which have a relatively low transmembrane diffusion coefficient for β-lactamases. 11 Their resistance to carbapenems is related to the decreased permeability of the bacterial outer membrane and hyperexpression of efflux pumps. 12 Most of the carbapenemase genotypes, such as VIM, IMP, SPM, GIM, AIM, DIM, SIM, KPC, BIC, OXA-48, and NDM, were described in these two bacteria, of which IMP and NDM were most frequently reported.13–15
The hospital environment represents a hotspot for the dissemination of resistant bacterial strains, and among the pathogens that cause nosocomial infections, the Gram-negative bacteria A. baumannii and P. aeruginosa are predominant. Infections caused by these microorganisms result in serious issues, specifically in the patients of the intensive care unit (ICU) due to multiresistance against antibiotics.2,16
Another factor contributing to antimicrobial resistance may be the ability of these pathogens to form biofilms. Biofilms are three-dimensional, multicellular complex structures consisting of cells in close contact with each other, presenting as communities, formed by the most diverse types of microorganisms, such as fungi, algae, protozoa, or by one or more bacterial species.17,18 In A. baumannii and P. aeruginosa, this ability contributes significantly to the persistence and versatility of these species causing nosocomial infections. Biofilm formation allows these bacteria to grow in unfavorable environments, thereby posing concerns by adhering to medical equipments such as catheters and endotracheal tubes, representing a significant risk for patients.14,19,20
The ability to form biofilm is one of the main virulence attributes in A. baumannii, allowing survival in adverse environmental conditions. Although biofilm formation is variable in this species, it involves chemical factors and signs that are not fully elucidated; this pathogen has the chaperon/usher system and the proteins OmpA and Bap, allows adhesion, development, and stabilization, as well as extracellular polysaccharide poly-β-(1,6)-N-acetylglucosamine, related to adhesion between biofilm cells.20,21
Fatal P. aeruginosa infections have been related to biofilm formation through chronic colonization, being able to secrete a large amount of matrix, leading to the formation of mucoid biofilm, the main characteristic of these infections. It mainly composed of extracellular polymeric substances, which are mainly polysaccharides, proteins, extracellular DNA (eDNA), and lipids. The exopolysaccharides Psl, Pel, and alginate are the main constituents of P. aeruginosa biofilm matrix involved in surface adhesion and, together with eDNA, determine the biofilm architecture, being responsible for cell migration, cell–cell interaction, protecting against phagocytosis and oxidative stress, providing resistance to immune and antimicrobial responses.18,22
In this context, this study reports the occurrence and dissemination of A. baumannii and P. aeruginosa resistance and biofilm formation in two public hospitals located at Recife, Pernambuco, Brazil, and discusses the importance of monitoring programs to combat the occurrence and dispersion of these pathogens in the hospital environment.
Methods
Ethics statement
This study was approved by the Ethics Committee on Research with Humans of the Universidade Federal de Pernambuco, Brazil (CEP: 1.480.085).
Clinical isolates
In total, 100 isolates were obtained; 35 from A. baumannii and 65 from P. aeruginosa from April to October 2016. Samples were collected from several infection sites of patients hospitalized in different sectors of the two public hospitals in Recife, Pernambuco, Brazil. Samples were seeded in brain heart infusion (BHI) broth and incubated at 37°C for 24 hr. The isolates were maintained in 25% (v/v) glycerol at −80°C.
Bacterial identification
The isolates were identified using the automated Vitek®2 Compact bioMérieux method. They were later confirmed by the MALDI-TOF mass spectrometry (MS) method, where the bacterial colonies were resuspended in 300 μL of Milli-Q water, after addition of 900 μL of absolute ethanol. The suspensions were centrifuged at 15,600 g for 2 min, the supernatant was removed, and the pellet was obtained after drying on vacuum concentrator for 5 min. Next, 50 μL of 70% (v/v) formic acid and 50 μL of acetonitrile were added. The mixture was vortexed and centrifuged at 15,600 g (2 min), and the supernatant was transferred to a new microtube. The matrix was prepared with α-cyano-4-hydroxycinnamic acid (10 mg/mL) in 50% (v/v) acetonitrile and 0.3% (v/v) trifluoroacetic acid and applied to the MALDI plate together with the sample at room temperature (25°C), for crystallization. The acquisition of the MS spectra in linear positive mode (acceleration voltage 20 kV and m/z detection range 2,000–20,000) was performed using the Flex Control version 3.0 software in a MALDI-TOF Autoflex III mass spectrometer (Bruker Daltonics, Billerica, MA). The spectra obtained were compared with the MALDIBiotyper version 3.1 database.
Antimicrobial resistance profile and determination of minimal inhibitory concentrations
The susceptibility profile and the minimal inhibitory concentrations (MICs) for the antibiotics were determined using the Vitek 2 Compact (bioMérieux) automated method. The following 21 antimicrobials were used to characterize the isolates: amikacin, aztreonam, cefazolin, cefepime, ceftazidime, ceftriaxone, cefuroxine, cefuroxine/axethyl, ciprofloxacin, gentamicin, imipenem, levofloxacin, meropenem, norfloxacin, ampicillin, ampicillin/sulbactam, cefalotin, cefoxitin, ertapenem, ticarcillin/clavulanic acid, and polymyxins B and E. Of these, eight (polymyxin B, ciprofloxacin, amikacin, gentamicin, meropenem, imipenen, cefepime, ceftriaxone, and ceftazidime) were common to both species. The resistance profile was evaluated and compared between the two hospitals investigated based on the frequency of MDR, XDR, and nonmultidrug-resistant (N-MDR) isolates.
Total DNA extraction
After incubation in BHI broth over a period of 18–24 hr, the total DNA was extracted using the PureLink® kit (Invitrogen) according to the manufacturer's instructions. The quality of the extracted DNA was evaluated through electrophoresis on 1% (w/v) agarose gel. After running the gel, it was observed under a UV transilluminator and photodocumented. The DNA concentration was determined by measuring the optical density (OD) at 260 and 280 nm in a NanoDrop 2000 (Thermo Fisher Scientific) spectrophotometer, and an estimate of the DNA amount present in the sample and the degree of purity was obtained.
Conditions for detection of blaKPC, blaIMP, blaVIM, and blaSHV genes through PCR
The isolates were subjected to polymerase chain reaction (PCR) to identify the blaKPC, blaIMP, blaVIM, and blaSHV genes. The reaction was prepared to a final volume of 25 μL per tube, comprising 50 ng genomic DNA, 10 pmol of the 1 × primer, 200 μM dNTPs, 1.5 mM MgCl2, and Taq DNA polymerase enzyme (2U) (Invitrogen). The reactions were carried out on a thermocycler (Biosystems) and, after preheating (95°C for 5 min), 35 cycles of 1 min at 95°C, 1 min at the specific temperature for each primer (Table 1), and 1 min at 72°C were performed. The final extension was 10 min at 72°C for the blaSHV and blaKPC genes and 5 min at 68°C for the blaIMP and blaVIM genes. The PCR products were visualized through electrophoresis under ultraviolet light and were then photodocumented.
Primers Used for Detection of β-Lactamases Genes
Biofilm formation
To evaluate the biofilm formation potential of the isolates, the crystal violet stain 26 was added to 96-well polystyrene flat-bottom microtiter plates using different culture media: Luria Bertani Miller (LB Miller), LB1/4, trypticase soy broth (TSB), TSB1/4, and TSB plus LB supplemented with 1% (w/v) glucose.
The strains were seeded in BHI agar and were incubated at 37°C for 24 hr. Colonies were inoculated in 0.85% NaCl and homogenized to avoid lump formation. Subsequently, the OD at 600 nm was measured and adjusted to be in the range of 0.145 to 0.155. Next, 20 μL of the bacterial suspension and 180 μL of one of the afore-mentioned culture media were added to the microwell plate. Eight replicates were prepared for each sample. The plate was incubated for 24 hr at 37°C. Later, the unadhered cells were removed by washing with 200 μL of 0.85% NaCl, and they were stained with 0.4% (w/v) crystal violet for 15 min. The adhered cells were fixed (30 min) with 200 μL of 100% ethanol, and OD was determined at 570 nm. The mean values of each sample (ODs) were determined in comparison with the OD in the sterility control (ODc). The samples were classified as strong (4 × ODc < ODs), moderate (2 × ODc< ODs ≤4 × ODc), or weak (ODc<ODs ≤2 × ODc) biofilm formers.
Conditions for enterobacterial repetitive intergenic consensus-PCR
For ERIC-PCR (enterobacterial repetitive intergenic consensus-PCR), 27 isolates of A. baumannii and 41 of P. aeruginosa were used. The ERIC-PCRs were prepared in a total volume of 25 μL per tube, comprising the following reagents: 100 μg genomic DNA, 10 pmol of primers (ERIC-1 [5′-ATG TAAGCTCCTGGGGATTCAC-3′]; ERIC-2 [5′- AAGTAAGTGACTGGGGTGAGCG-3′]), 1 × buffer, 200 μM deoxyribonucleotide triphosphate, 1.5 mM MgCl2, and 1.0 U of the DNA Taq polymerase enzyme. The following amplification parameters were used in the ERIC-PCR: the initial denaturation cycle involved 95°C for 3 min, followed by 30 cycles of denaturation at 92°C for 1 min, annealing at 36°C for 1 min, and extension at 72°C for 8 min. After 30 cycles, a final elongation step of 16 min at 72°C ensued. The PCR products were stained with greenish blue stain and were subjected to 1.5% agarose gel electrophoresis, visualized under UV light, and photodocumented for later analysis of clonal profiles. 27
Results
Bacterial identification
The taxonomic identification of the isolates was obtained using the automated Vitek 2 Compact Biomérieux method and was confirmed by MALDI-TOF MS analysis, with score values between 2,099 and 2,900, indicating high compatibility and reliability indices (Table 2).
Identification of Bacterial Isolates by Acquisition of Mass Spectra, Hospital (Sector), Infection Site, Resistance Profile, Biofilm Formation, Genotype and Group of Clones
In bold, the clonal groups.
Isolates used in ERIC-PCR.
H1A, A. baumannii-Hospital 1; H2A, A. baumannii-Hospital 2; H1P, P. aeruginosa-Hospital 1; H2P, P. aeruginosa-Hospital 2; ANSU, Advanced Neurosurgery Support Unit; UCO, Coronary Unit; BTU, Burn Treatment Unit; ICU, Intensive Care Unit.
Antibiotic susceptibility profiles
The resistance profile of A. baumannii and P. aeruginosa isolates to the nine antibiotics tested was evaluated (Fig. 1), and the percentages between the sectors and the different hospitals investigated (Table 2) were compared. Ciprofloxacin (class of fluoroquinolones) exhibited less efficacy against both A. baumannii (91.4%) and P. aeruginosa (84.6%) species, thereby indicating drug resistance. For the other antibiotics, the two species exhibited different resistance profiles, being mainly resistant to the β-lactam group (imipenem, meropenem, ceftazidime, and ceftriaxone). All isolates were sensitive to polymyxins B and E.
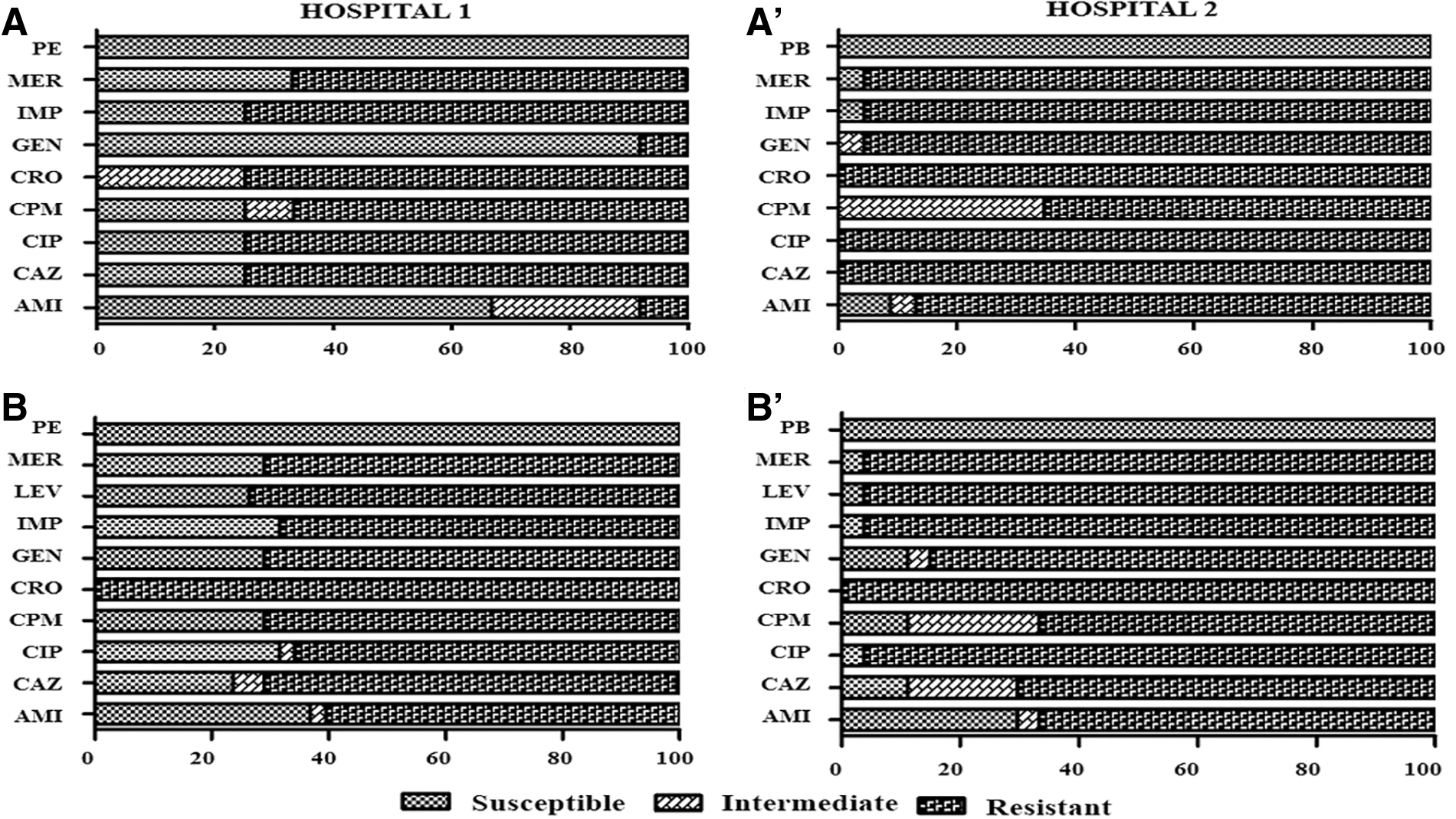
Antimicrobial susceptibility profiles of Acinetobacter baumannii and Pseudomonas aeruginosa isolates in both hospitals:
The Hospital 2 (H2) had the highest number of isolates classified as XDR, presenting 95.7% of A. baumannii and 81% of P. aeruginosa species. In Hospital 1 (H1), 75% and 47.4% of A. baumannii and P. aeruginosa were XDR, respectively. The A. baumannii species had the highest number of XDR isolates distributed in both hospitals. In contrast, only P. aeruginosa had MDR isolates, found in both hospitals (H1: 23.7% and H2: 14.8%, Table 2). The most frequent origin of the isolates for both species was tracheal secretion (36%), followed by urine (17%) and cerebrospinal fluid (11%). The ICU of both hospitals presented the highest percentage of patients infected with both species, totaling 39% of the isolates.
Detection and genotypic frequency of the genes blaSHV, blaKPC, blaIMP, and blaVIM
Among the tested isolates, 60 (54.3% of A. baumannii and 63% of P. aeruginosa) presented the genotype KPC for blaKPC, whereas 39 (22.85% of A. baumannii and 38.46% of P. aeruginosa) revealed the genotype SHV of blaSHV gene. The genotype VIM of blaVIM was detected in eight isolates (2.8% of A. baumannii and 7.7% of P. aeruginosa), and the genotype IMP of blaIMP was observed only in one isolate of A. baumannii (2.85%). The gene combination more frequently observed was blaSHV+blaKPC (26%), followed by blaKPC+blaVIM (3%), blaSHV+blaVIM (2%), and blaSHV+blaKPC+blaVIM (1%). The combination between these four genes was not identified in any sample (Table 2). H1 revealed a higher percentage of KPC (75% for A. baumannii and 76.32% for P. aeruginosa), whereas H2 had 43.48% and 44.44% for the two species, respectively. The sequencing of the amplicons confirmed the molecular sequence of the amplified fragments revealing high similarity level and homology when compared with the sequences deposited in the GenBank at NCBI.
Biofilm formation in different growth media
All the analyzed isolates formed biofilms in the tested media (TSB, TSBG, TSB1/4, LB, LBG, LB1/4) but with different intensities. In A. baumannii, the TSB medium proportioned the highest biofilm formation, whereas for P. aeruginosa, it was the LBG medium (Fig. 2).
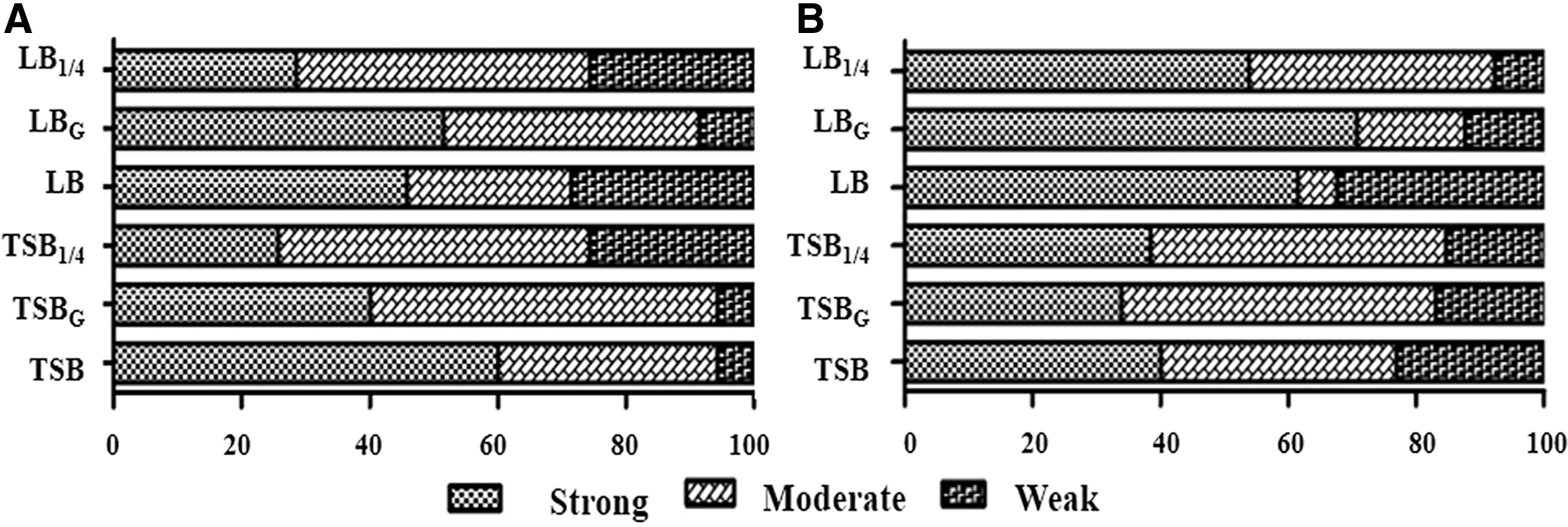
Biofilm formation in different growth media by the isolates of A. baumannii
Among the 35 isolates of A. baumannii, 24 (68.6%) were revealed as strong biofilm formers (23 XDR and one N-MRD), 10 (28.6%) were moderate formers (9 XDR and 1 N-MDR), and only 1 (2.9%) was a weak former, indicating an XDR profile (Fig. 3A). In regard to the 65 isolates of P. aeruginosa, 59 (91%) exhibited strong biofilm formation (47 XDR, 11 MDR, and 1N-MDR), 3 (4.6%) indicated moderate formation (2 XDR and 1 MDR), and 3 (4.6%) revealed weak formation (2 XDR and 1 MDR).
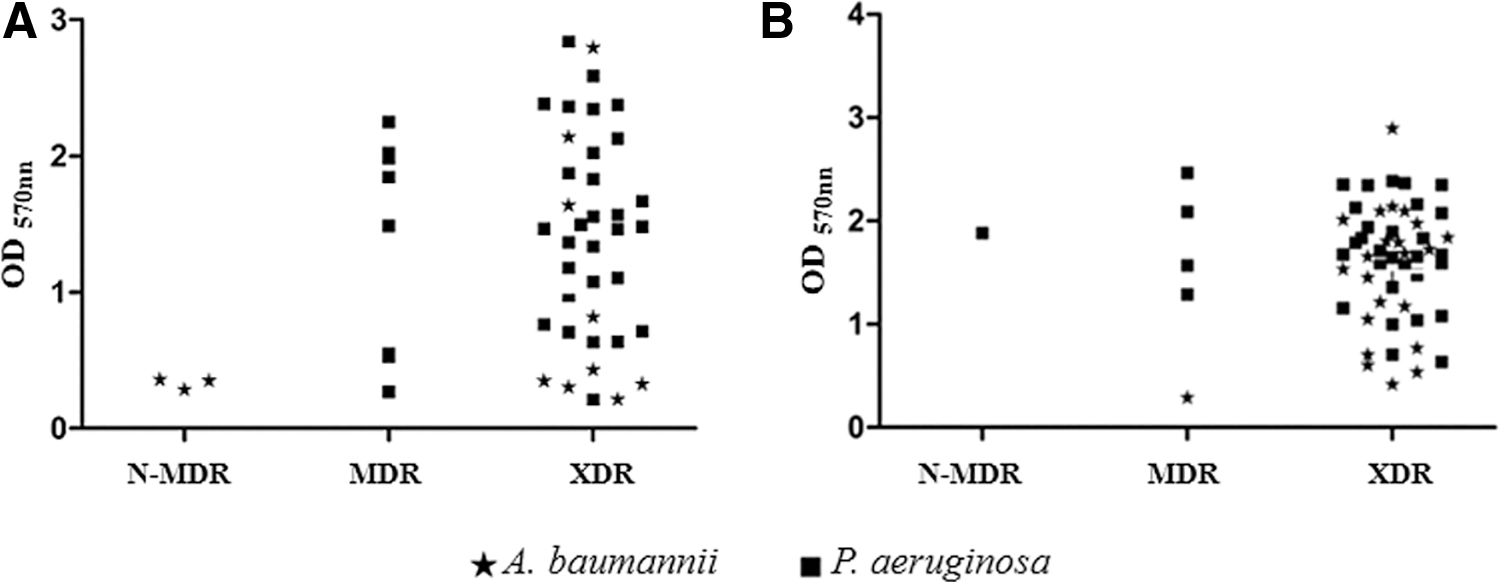
Formation of biofilm and resistance profile correlation for A. baumannii and P. aeruginosa isolates from hospital 1
It was not possible to correlate the resistance profile of the organism with its capacity of biofilm formation. In the two species, XDR isolates presented strong, moderate, and weak biofilm formation. Figure 3 presents the distribution of isolates according to the resistance profile and the capacity of biofilm formation in both hospitals.
ERIC-PCR
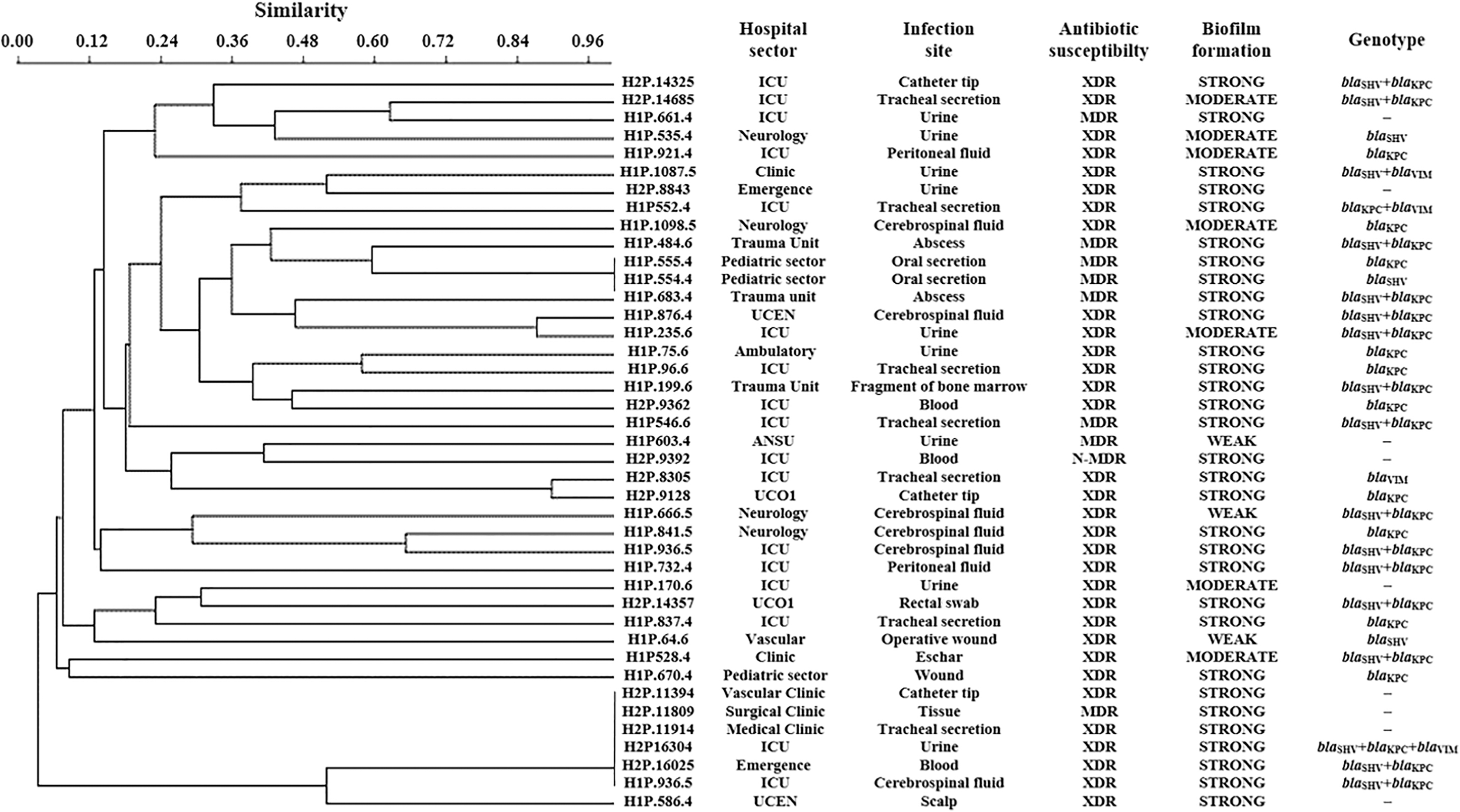
In this study, we identified 12 clones of A. baumannii further divided into five groups (1A, 2A, 3A, 4A, and 5A) and 8 of P. aeruginosa (1B and 2B), as illustrated in Figs. 4 and 5. The data are indicative of a possible intrahospital clonal dissemination occurred in six H1 and three H2 sectors; and the interhospital spread was observed in Group 3A (A. baumannii) and in Group 2B (P. aeruginosa). The data indicated here revealed a greater dispersion of these isolates between the different sectors of the same hospital and to a lesser extent among the different hospitals studied.

Dendrogram of the ERIC-PCR method on discrimination of genetically related and unrelated isolates of A. baumannii. ERIC-PCR, enterobacterial repetitive intergenic consensus-polymerase chain reaction.
Dendrogram of the ERIC-PCR method on discrimination of genetically related and unrelated isolates of P. aeruginosa.
Discussion
In this study, the identification of the isolates was initially performed by an automated method described in the methodology. Subsequently, the identity was confirmed by MALDI-TOF MS based on protein analysis. This technique generates mass spectra of the proteins from the unknown microorganism providing a proteomic fingerprint, which is then compared with a reference database, leading to precise identification at the genus and species level. 28 The identity of all bacterial isolates was confirmed with 100% agreement between the results obtained and the database records.
Infections caused by XDR and MDR bacteria are extremely difficult to treat. The antimicrobial susceptibility test revealed that the isolates of A. baumannii and P. aeruginosa studied here presented high levels of resistance to most of the antibiotics tested in both hospitals. A. baumannii isolates presented resistance >90% to cephalosporins of third generation and 65% to those of second generation. In addition, it presented >90% resistance to carbapenems, which are usually the drugs of choice for the treatment of these infections.29–31 Approximately 60% of the isolates were resistant to aminoglycosides and 90% to fluoroquinolone tested. Similar results were reported by other authors who demonstrated the high resistance of isolates of this species to antibiotics of different action classes.31,32
Most of the isolates of P. aeruginosa exhibited resistance to fluoroquinolone (86%), cephalosporins (70%), and aminoglycosides (>60%). The high index of resistance to these antibiotics has been reported by other authors, contrasting only in the percentage found. 33 A previous study by Ribeiro et al. 34 reported amicacin as a treatment option for these microorganisms; however, this study demonstrated an increase in resistance to this drug in both species, restricting the treatment options to polymyxins. These results are worrying and reinforce the need to address new therapeutic alternatives.
Compared with previous studies,32,35,36 this study reinforces the occurrence of XDR and MDR isolates of the species investigated in the hospital environment. These data demonstrate the relevance of this study and the need to monitor dissemination of these pathogens in the hospital environment.
These data present the frequent isolation of ESBL-producing strains, with different variants, indicating therapeutic failure by administering the cephalosporins of first and second generation. This consequently contributed to the choice of treatment with carbapenems; however, this led to the onset and spread of carbapenemases (KPC-2, SPM-1, PXA-23, IMP-1, and OXA-143) throughout Brazil, related to health care-associated infections. 8 The rate of nosocomial infections caused by ESBL in Brazil is among the highest in the world.36,37 In this study, the incidence of beta-lactamase enzymes KPC (60%), SHV (36%), VIM (10%), and IMP (1%) indicates the need to implement effective measures to control infections through microbiological surveillance and monitoring programs of antimicrobial resistance standards to reduce the morbidity and mortality rates caused by these microorganisms and associated hospital costs.
It was possible to compare and confirm the resistance profiles by biochemical methods in the presence of the resistance genes investigated, except for a few strains (two of A. baumannii and six of P. aeruginosa) that were positive for the KPC gene, but were not resistant to carbapenems. This is because the acquisition of carbapenemase genes is not always enough to guarantee the resistance. A genetic suppression that leads to gene silencing or even the presence of other resistance mechanisms can evade phenotypic evidence.38,39 Moreover, some strains were not positive for the studied genes; however, they presented the XDR and MDR phenotypic profiles, which can be related to the presence or association of other mechanisms of resistance and virulence that both the species possess, such as hyperexpression of efflux pumps, biofilm formation, and iron acquisition systems.18,20,40–43
Among all the genes investigated, blaKPC was the most abundant, and present in 60% of the isolates (in H1, 75% of A. baumannii and 71% of P. aeruginosa, and in H2, 43% of A. baumannii and 44.4% of P. aeruginosa). The difference between the two hospitals may be related to the species, as well as to the therapeutic practice used in different hospitals. This difference shows the importance of microbiota study and the profile of each hospital even when they are located in the same city.36,44 The first preferred drug class for treating infections by these microorganisms is the carbapenems, but the increasing occurrence of resistance toward this class of drugs has limited therapeutic options, increasing mortality rates, and often restricting treatment options to polymyxins or tigelin.31,39
In this study, the KPC gene frequency was higher in the strains of A. baumannii. These data reveal an increase in the incidence of A. baumannii KPC positive in Brazil when compared with the previous studies. Ribeiro et al. 34 found an incidence of 16.4% in 21 isolates of A. baumannii they investigated. For P. aeruginosa, the first report of blaKPC in Brazil appeared in Recife. 45 In 2016, Jacome et al. 33 detected the presence of this gene in 25.8% of the 58 isolates they investigated, whereas in this study a percentage of 63.0% (65 isolates) was observed, demonstrating the constant increase of gene incidence in Brazil. The increased prevalence of the MDR-KPC clinical phenotype has been associated with high mortality rates.34,46
All the investigated isolates of both species formed biofilms in the different media tested, but with diverse intensities. A. baumannii presented higher biofilm formation in the TSBG medium (64.6%), whereas P. aeruginosa exhibited higher ability in LB medium (90.8%). Noteworthily, only the last species responded to the stress treatments when subjected to variation in TSB medium for biofilm formation. Several biofilm-forming isolates were observed when subjected to 25% TSB medium by comparing the results obtained in TSB medium at normal concentration and in TSB medium plus glucose. This result occurs when exposed to stress conditions, since the microorganism presumably increases its pathogenicity and synthesis of virulence factors. In this case, a low concentration of supplements may have led to a higher biofilm formation. 18
The profiles of strong biofilm formers in both species were predominantly independent of the antimicrobial susceptibility. This result demonstrates the increasing virulence degree of these species and the severity of the infection caused by them. Presently, the authors differ on their views regarding the biofilm formation ability and the antimicrobial resistance phenotype. Certain studies with A. baumannii and P. aeruginosa, in addition to other species, affirm that there is a positive relationship between these two events, considering that the MDR profile is directly linked to high biofilm formation.47–50 However, other authors concluded that there is a possible inverse relationship between resistance and biofilm production; that is, biofilm formation is directly related to the N-MDR profile.32,51,52 In this study, no significant relationship was observed between the biofilm production and the resistance phenotypes, indicating the presence of multifactorial events or that biofilm formation is an intrinsic characteristic of the pathogens investigated.31,33,53
One previous study has reported that the presence of plasmids containing genes for classes A and D β-lactamases has the potential to cause disturbances in the bacterial adhesion structures essential for biofilm formation and motility. This property was observed in strains of Escherichia coli and P. aeruginosa, and is specific only for these classes. However, class C did not present this property. 54 In this study, the class A β-lactamases (blaSHV and blaKPC) did not interfere in the biofilm formation, thus demonstrating a greater virulence in these species.
The ERIC-PCR technique was used to confirm whether the isolates present correspond to the clones that are being disseminated between the sectors and hospitals. This technique has been widely used for the molecular typing of different bacteria in epidemiological studies, and its advantages are well known55,56 and have been used for Gram-positive 57 as well as Gram-negative bacteria. 58 Li et al., 59 Jacome et al., 36 and Lima et al. 53 were able to identify the clones in isolates of A. baumannii and P. aeruginosa using this technique.
The ERIC-PCR revealed that 12 of the analyzed isolates of A. baumannii presented the same pattern being indicative of clonality (Fig. 4 and Table 2) grouped into five profiles (1A–5A), each containing 2–3 isolates, indicating clonal dispersion of bacteria from the same sector, between different sectors (neurology, ICU, recovery room, pediatric sector, UCO, and emergency), and to a lesser extent among the two hospitals analyzed (H2A8747, H2A14967, and H1A620.5; Fig. 4A). Li et al. 31 obtained similar results for isolates of A. baumannii of hospital origin by the ERIC-PCR technique (of the 106 isolates tested, 78 were found as clones). In relation to P. aeruginosa, eight isolates presented the profile segregated into two (1B and 2B) distinct groups (Fig. 5); that is, one with two isolates from the same hospital sector (pediatric sector) and the same phenotypic profile, and a second group comprising six isolates, one of which had a different hospital origin, whereas the others differed only in the sectors of origin, with or without phenotypic similarities.
Considering the isolates that showed clonal relationships, few presented different profiles of susceptibility or positivity for some of the resistance genes analyzed. This is in accordance with other studies that did not observe this relationship between the ERIC-PCR profiles, and other techniques (PFGE and RAPD), with profiles of microbial susceptibility, considering relative evidence of clonality.39,60–62
Figure 6 provides an overview of the different bacterial profiles found in the hospital sectors, and their intra- and interhospital dissemination. The results described in this study demonstrate the critical situation in the transmission of hospital infections, reinforcing the need for intervention with strategies for both treatment and monitoring of these infections as well as prevention of the cross-infection. It is also worrying that several authors have suggested health professionals as one of the dissemination vehicles of these infections based on molecular techniques.63,64

Intra- and interhospital dissemination of A. baumannii and P. aeruginosa in public hospitals in Recife, Pernambuco, Brazil.
Conclusion
This study presents the occurrence and dissemination of drug-resistant and biofilm-forming A. baumannii and P. aeruginosa isolates in hospital environments. The high occurrence and dissemination of microbial profiles observed in A. baumannii and P. aeruginosa indicate the difficulty in the effective choice of treatment of these microorganisms. These data are worrying and reveal the present critical scenario of nosocomial infections, emphasizing the need for continuous monitoring, with epidemiological surveillance of these infections, thereby helping in minimizing their spread in the hospital environment.
Footnotes
Acknowledgment
The authors are grateful to the Analytical Central of Centro de Tecnologias Estratégicas do Nordeste (CETENE) by space given.
Disclosure Statement
No competing financial interests exist.
Funding Information
The authors are grateful to the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for financial support and fellowship (THN and PMGP); the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES; Financial Code: 001) for financial support; and the Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco (FACEPE; APQ-0661-2.08/15) for financial support.