
Abstract
The multiresistance plasmid, pZM3, from a 1970 Salmonella enterica serovar Wien isolate from Algeria represents the multiresistance FIme-type plasmids conferring resistance to ampicillin, chloramphenicol, kanamycin, neomycin, sulfonamides, streptomycin, spectinomycin, tetracycline, and mercuric ions circulating in the Middle East in the 1970s. pZM3 was sequenced to determine the relationship between IS1936, the IS26-like insertion sequence it carries, and IS26. IS1936 is identical to IS26. pZM3 is a 166.8-kb plasmid with three replicons typed as FIA-1, FIB-1, and FII-1, consistent with other FIme plasmids. However, Tn3, containing the blaTEM-1a ampicillin resistance gene, disrupts the FII repA gene. pZM3 also contains an IS1-flanked virulence region, including the sit and aerobactin operons, shared with many other FIB-1 virulence plasmids. The remaining resistance genes are located in a 44.7-kb complex resistance island that includes the Tn21-like transposon, Tn1935, identified previously. Relative to Tn21, Tn1935 includes an additional gene cassette, oxa1, and Tn4352 in tniA. Tn1935 is in the same Tn2670 context as Tn21 in NR1, and identity to NR1 extends beyond the IS1 flanking the catA1 gene. On the other side, IS1-mediated events have brought in a Tn10 remnant and inverted part of it, highlighting the role of IS1 in resistance region evolution. The backbone of pZM3 was found to be almost identical to that of pRSB225, recovered in Germany in 2013, and their resistance islands are in the same position. The pRSB225 resistance island has evolved in situ from the pZM3 configuration through an insertion, a replacement, and an inversion.
Introduction
Plasmids are important vehicles for dissemination of both antibiotic resistance genes and virulence determinants in bacterial populations. Many contemporary plasmids carry complex, mosaic, antibiotic resistance islands that are many evolutionary steps removed from their progenitor. They often contain a cluster of mobile genetic elements, including insertion sequences and transposons, together with many different antibiotic resistance genes, making the relationships between different plasmids difficult to unravel. On the other hand, older plasmids recovered in the presequencing era can provide valuable insights into the origin and evolution of the multiple antibiotic resistance and virulence plasmids circulating in bacterial populations today.
Between 1969 and 1980, Salmonella strains carrying plasmids containing genes conferring resistance to multiple antibiotics and virulence determinants were responsible for a number of epidemic outbreaks in North Africa and Europe.1,2 The plasmid pZM3 was first identified in ZM3, a Salmonella enterica serovar Wien (previously known as Salmonella wien) isolate recovered in 1970 in Algeria,1,3 and ZM3 was the earliest of the epidemic S. enterica Wien strains that was believed to be representative of the North African epidemic. 3
pZM3 was characterized by classical incompatibility testing as an FIme plasmid, a subgroup of FI plasmids recovered in the Middle East, which at the time were known to be incompatible with other IncFI plasmids and the MP10 plasmid isolated from S. enterica Typhimurium phage type 36.1,4 When PCR-based replicon typing was later introduced, the plasmids belonging to the FIme group were shown to carry FIA-1, FIB-1, and FII-1 replicons. 5 A combination of restriction mapping and electron microscopy was originally used to estimate the size of pZM3 at 165 kb. 3 It was shown to carry a 23.5 kb Tn21-like transposon, which was named Tn1935. Tn1935 contains the entirety of Tn21, conferring resistance to spectinomycin, streptomycin, sulfonamides, and mercuric ions, plus two additional fragments of 0.95 and 2.7 kb conferring resistance to ampicillin and kanamycin, corresponding to oxa1 and a kanamycin resistance transposon, respectively.2,3 Southern hybridization with an IS26-specific probe (IS15, made up of two copies of an IS26 variant, one inside the other, was used) demonstrated that the 2.7-kb fragment was a composite element, comprising two IS26-like insertion sequences named IS1936 in direct orientation, flanking the kanamycin resistance determinant.
Further analysis demonstrated that IS1936 was ∼800 bp in size, and restriction mapping showed that IS1936 had single restriction sites for SalI and PstI in an arrangement that had previously been reported for IS15Δ1, IS26, IS46, IS140, IS160, and IS176. 3 Both copies of IS1936 surrounding the kanamycin resistance gene could mediate cointegrate formation using a replicative mechanism at a frequency of ∼2 × 10−5 cointegrates per transconjugant. 3 It is now known that IS46, IS140, IS160, and IS176 are identical to IS26, 6 and IS15Δ1 (also called IS26* or IS26-v1) only differs by three nucleotides from IS26. Hence, the available data suggested that IS1936 was likely identical or nearly identical to IS26. However, like IS6, the IS after which the family that IS26 belongs to was named, IS1936 had never been sequenced.
In this study, the complete sequence of pZM3 was determined to clarify the relationship of IS1936 with IS26. This revealed a complex resistance region and a number of virulence determinants, which had only been partially characterized previously.
Materials and Methods
Bacterial strain and conjugation
The S. enterica serovar Wien strain ZM3 was kindly provided by Prof. Alessandra Carattoli (Istituto Superiore di Sanita, Rome, Italy). ZM3 contains a single conjugative FIme plasmid, pZM3.
3
The antibiotic resistance profile of ZM3 was confirmed using a disc diffusion assay method, as described previously.
7
An annular radius of <6 mm was indicative of resistance. The plasmid was first transferred from ZM3 to the nalidixic acid-resistant Escherichia coli strain UB5201 (pro met recA gyrA, NxR) by mixing equal amounts of stationary-phase cultures on
Genome sequencing
The complete genome of strain ZM3 was determined by sequencing using long-read MinION (Oxford Nanopore) and short-read Illumina MiSeq technologies. Genomic DNA was prepared as described previously. The MinION run (210 × depth) generated a total of 48,640 reads and the Illumina MiSeq run (40 × depth) generated a total of 866,542 150-bp paired-end reads. The MinION reads were filtered using Filtlong (version 0.2.0) to remove any reads <1,000 bp, followed by removal of the lowest 10% of reads by quality. The long and short reads were combined in a hybrid assembly using Unicycler (version 0.4.0) 8 with default assembly parameters.
Protein coding, rRNA, and tRNA genes were annotated using Prokka (version 1.12). 9 Antibiotic resistance genes and plasmid replicons were identified using ABRicate (version 0.8.13) and were annotated manually. ISs were identified using the ISfinder database 10 and annotated manually. The PubMLST database was used to sub-type the plasmid replicons. Plasmids related to pZM3 were identified using a standard BLAST search and filtering manually for complete plasmid sequences.
Nucleotide sequence accession number
The 166,767-bp nucleotide sequence of pZM3 has been deposited in GenBank under accession no. MK797990.
Results
pZM3
Plasmid pZM3 was recovered from isolate ZM3 by conjugation into E. coli UB5201 (NxR) and selection for kanamycin (Km) resistance. All pZM3 transconjugants were also resistant to ampicillin (Ap), chloramphenicol (Cm), streptomycin (Sm), sulfonamides (Su), and tetracycline (Tc), as reported previously. 1 In addition, the transconjugants were also resistant to neomycin (Nm) and spectinomycin (Sp). Although pZM3 had previously been reported to be conjugative, 3 the transfer frequency was not reported. The E. coli–E. coli conjugation efficiency was determined by mating UB5201 containing pZM3 with UB1637-rifR and selecting for transconjugants using rifampicin and kanamycin. pZM3 transferred at an average frequency of 4.3 × 10−4 transconjugants per donor (average of three independent determinations; 5.2 × 10−4, 6.5 × 10−4, and 1.2 × 10−4). This frequency is similar to that reported for other FIA-FIB-FII plasmids. 11 Several presumptive transconjugants were tested by disc diffusion assay and by patching onto solid media containing antibiotics, and all transconjugants had gained the resistance phenotype of pZM3.
Sequence of pZM3
The hybrid assembly of strain ZM3 from short- and long-read sequence data generated two circular molecules of 4,512,656 and 166,767 bp, representing the S. enterica serovar Wien ZM3 chromosome and plasmid pZM3, respectively. The copy number of pZM3, estimated by comparing the coverage of the plasmid molecule with the coverage of the circular chromosome, was 1.49.
The ABRicate output indicated that pZM3 contains FIA, FIB, and FII replicons. Using the PubMLST database, they were further sub-typed as FIA-1, FIB-1, and FII-1. This is consistent with the replicon combination previously reported for pZM3 and other FIme plasmids that were recovered in the 1970s and 1980s and subsequently replicon typed using PCR-based replicon typing (PBRT). 5 ABRicate also indicated that pZM3 contains the aphA1a (kanamycin and neomycin), aadA1 (streptomycin and spectinomycin), blaTEM-1a (ampicillin), catA1 (chloramphenicol), oxa1 (ampicillin), sul1 (sulfonamides), and tet(B) (tetracycline) resistance determinants. The blaTEM-1 gene is within a complete copy of Tn3 (GenBank accession no. V00613), 12 which has interrupted the FII repA gene, and is surrounded by 5-bp target site duplication (TSD). The remaining resistance genes are contained in a single 44,717-bp complex resistance region (see “The pZM3 resistance region”) that is not surrounded by TSD.
IS1936 is IS26
The regions surrounding the aphA1a kanamycin and neomycin resistance gene are 100% identical to IS26. 13 Indeed, the 2,680-bp region containing aphA1a together with the flanking IS is identical to Tn4352 (Fig. 2A), which was first characterized from a small S. enterica Typhimurium multicopy plasmid in 1987, 14 1 year before pZM3 was first characterized. The sequence of IS1936 has been submitted to the ISfinder database to consolidate the entry with the existing IS26 record as a synonym.
pZM3 backbone
The backbone of pZM3 was derived by removing the complete copy of Tn3, and the TSD generated by its insertion, and the large 44.7 kb resistance island that includes Tn1935, from the complete pZM3 sequence. This generated a backbone of 117,097 bp (Fig. 1). Comparison of the backbone of pZM3 with other plasmids in the GenBank nonredundant database showed that like many F-type plasmids, the backbone of pZM3 is a mosaic of backbone regions derived from many different plasmid lineages.

Circular map of pZM3. pZM3 is drawn to scale from GenBank accession no. MK797990. Genes coding for proteins of known function or predicted function are shown as colored arrows, with only some of the important genes labeled. Insertion sequences are shown as colored boxes. The positions of the large resistance island and Tn3 are shown by arrows. The location and extent of the sit and aerobactin operons, and the leading region and transfer region, are marked. Color images are available online.
pZM3 has a complete conjugative transfer region (Fig. 1) that shares ∼95% nucleotide identity with the transfer region of plasmid F (GenBank accession no. AP001918) and contains all of the genes previously shown to be essential for conjugative transfer in plasmid F. 15 A copy of IS1 and a partial copy of ISKpn26 interrupt the otherwise contiguous transfer region, but have not abolished transfer ability. The leading region of pZM3, located between the FIA repA and the conjugative transfer region, contains all the genes previously shown to be essential for survival in the host immediately following conjugative transfer.15,16 This includes genes encoding the single-stranded binding protein Ssb (ssb), SopA (sopA) and SopB (sopB) partitioning proteins, KlcA (klcA) antirestriction protein, and YhdJ (yhdJ) DNA adenine methyltransferase. Two toxin/antitoxin systems (pemIK and ccdA/ccdB)17,18 were also identified in the pZM3 backbone (Fig. 1).
pZM3 shares regions with other virulence plasmids
In pZM3, two sets of virulence-associated genes are located between two inversely oriented copies of IS1 (Fig. 1) in a transposon-like configuration that has been described previously1,19,20 and is also found in ColV virulence plasmids. 21 The iutA/iucABCD operon contains a set of genes coding for the synthesis of bacterial iron-chelating siderophore aerobactin and its uptake. 22 Aerobactin is a virulence factor that enables the bacteria to sequester iron in iron-poor environments such as the gastrointestinal tract or the urinary tract. The second virulence-associated operon, known as the sit operon, contains four genes (sitABCD) that encode a high-affinity iron and manganese transport system. 23 This configuration of the two operons, bounded by the two copies of IS1, is found in over 130 plasmids or chromosomes in the GenBank nonredundant database (last searched May 27, 2019). The location in pZM3, adjacent to the backbone region containing the FIB-1 replicon, is particularly common, accounting for 117 of the 130 sequences in GenBank.
The nucleotide identity throughout the remaining backbone, including the transfer and leading regions, varies from sharing no significant identity to sharing up to 96% identity in some regions. This further supports the conclusion that like many other F-type plasmids, these plasmids are complex mosaics made up of different segments with many different origins that have come together over time.
The pZM3 resistance region
The 44,717-bp pZM3 resistance region (Fig. 2A) is a complex mosaic of complete and partial transposons and insertion sequences, bounded at one end by IS26 and at the other end by IS1. At the core of this island is Tn1935, which had been previously characterized by restriction mapping and Southern hybridization as a Tn21-like transposon. 3 The Tn1935 sequence is identical to that of Tn21, 24 except for the additional oxa1 gene cassette, and the presence of Tn4352, which has interrupted the tniA gene of the integron and has generated an 8-bp TSD (CATCGGCG). The precise junctions between IS26 and tniA are found in hundreds of plasmids and chromosomal resistance islands in the GenBank nonredundant database.
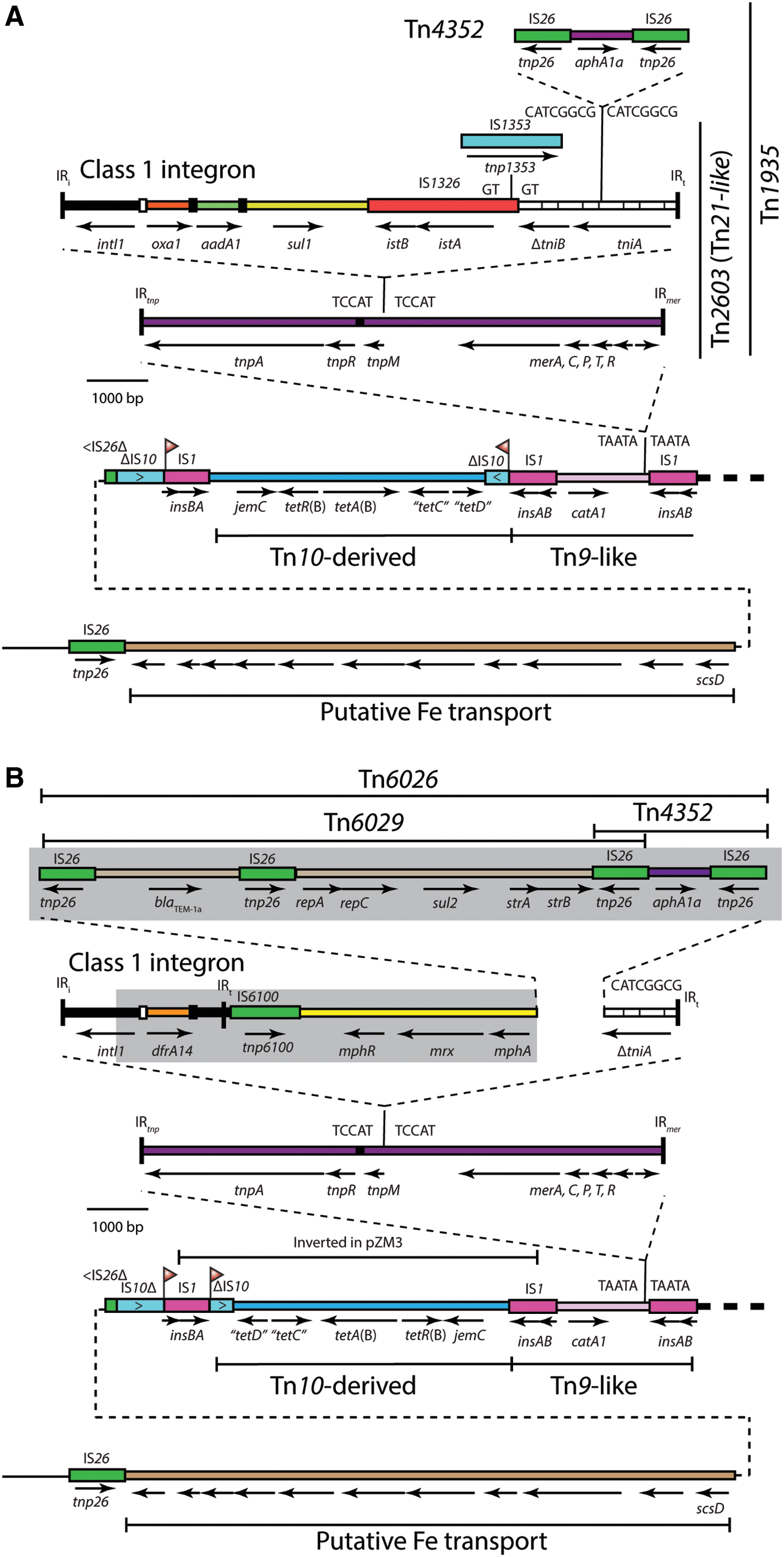
Linear maps of the resistance islands of
Tn1935 is also closely related to Tn2603, 25 which contains the oxa1 gene cassette and is the source of the first nucleotide sequence of oxa1, 26 but does not have Tn4352 in tniA. In pZM3, Tn1935 is inserted in a catA1-containing Tn9-like transposon and surrounded by a 5-bp TSD (TAATA). This position is identical to that of Tn21 in Tn2670 from NR1 (R100),24,27,28 indicating that the evolution from Tn21 occurred in situ. Furthermore, ∼5.3 kb of backbone adjacent to the right of the resistance island (as shown in Fig. 2A), including pemIK and the FII-1 repA, is the same in pZM3 and NR1. As NR1 was isolated in the late 1950s, this suggests acquisition from NR1 or a close relative. However, the sequence adjacent to Tn10 (left side in Fig. 2A) is not from NR1.
To the left of the Tn2670-derived transposon is a 6,526-bp region containing a large portion of Tn10, including the tet(B) tetracycline resistance determinant (Fig. 2A). Interestingly, NR1 contains both Tn2670 and Tn10, separated by a 14-kb backbone region that is absent from pZM3 (Fig. 3), giving rise to the possibility that the configuration in the resistance region in pZM3 arose through an IS1-catalyzed deletion event in NR1 or a closely related plasmid. In pZM3, this region has acquired an additional copy of IS1 through an IS1-mediated adjacent inversion event that has interrupted IS10 and inverted the Tn10-derived fragment between the two inversely oriented copies of IS1 (Fig. 3). This event is evidenced by the presence of inversely oriented 9-bp TSD (denoted by flags in Fig. 2A) in pZM3.

Evolution in situ of resistance islands derived from Tn2670 and Tn10. Regions are drawn to scale from GenBank accession nos. MK797990 (pZM3), JX127248 (pRSB225), and DQ364638 (NR1). The plasmid backbone shared with NR1 is represented on the right by a thick dashed line. The dashed line and double line on the left denote a sequence that differs between NR1 and pZM3/pRSB225. Insertion sequences are shown as boxes. The location and orientation of TSDs are indicated by vertical flags or uppercase letters. The orientation and extent of genes are indicated by horizontal arrows. The location of Tn21 or Tn1935 is marked by a vertical line. Color images are available online.
Beyond the Tn10 fragment is an 11.7-kb region containing a number of uncharacterized genes predicted to encode proteins involved in high-affinity Fe2+ binding (Fig. 2A), another putative virulence factor. The origin of this fragment is unknown. However, the configuration of this fragment adjacent to the remnant of Tn10 was recently found in the ARI-A resistance island of the IncC plasmid pB2-1 (isolated before 1995, see Fig. 2 in Harmer and Hall 29 ). Consistent with the scenario described above, there is no inversion in the Tn10 remnant in pB2-1.
Related plasmids
The closest match in the GenBank nonredundant database to the pZM3 backbone is an FIA-1:FIB-1:FII-1 plasmid, pRSB225 (GenBank accession no. JX127248), recovered from an uncultured bacterium in Germany in 2013. 30 pZM3 and pRSB225 share 99.97% identity across 96% of the backbone. The main difference between the backbones of pZM3 and pRSB225 is the presence of three extra fragments in pZM3, namely a group II intron that has interrupted the copy of IS629 (Fig. 1), an 1,192-bp fragment of unknown origin that does not contain any mobile genetic elements or genes of known function, and a small insertion of 163 bp. A 50-bp backbone region also differs between both pZM3 and pRSB225. pRSB225 does not have a copy of Tn3, but it does include a large resistance island in the same backbone position as the resistance island in pZM3.
The next closest match to pZM3 is plasmid 2 of the E. coli strain MS14385 (GenBank accession no. LR130556), sharing 99.96% identity across only 84% of the backbone, and E. coli strain DA33137 plasmid pDA33137-178 (GenBank accession no. CP029580), sharing 99.94% identity across only 64% of the backbone. Both backbones largely differ from pZM3 due to recombination with related plasmids within the transfer region. For example, the region between trbA and traL has been replaced between pZM3 and MS14385 plasmid 2. These plasmids also have resistance islands in a different location to the one in pZM3 and were not analyzed in further detail.
Evolution of the pRSB225 resistance region
The plasmid pRSB225 harbors a resistance island (Fig. 2B) in the same location as that in pZM3, but the internal resistance gene content is different. The differences can be traced to three distinct events.
The first is homologous recombination between the two copies of IS1 that had inverted the tet(B)-containing fragment of the copy of Tn10 in pZM3 (Fig. 3). This has returned this region to the original orientation seen in Tn10 and pB2-1, and the inversely oriented 9-bp TSDs seen in pZM3 are in direct orientation surrounding the IS1 in pRSB225.
The second event was the replacement of the region between intI1 and tniA, which has changed the internal content of the class 1 integron. pRSB225 contains an unusual In4 family integron31,32 that was first characterized in the IncHI1 plasmid, pHCM1 (GenBank accession no. AM412236), found in a multidrug resistance S. enterica Typhimurium strain. 33 It contains a dfrA14 gene cassette, conferring trimethoprim resistance, and a copy of the IS26-IS6 family insertion sequence IS6100. In comparison with the integron of pHCM1, the outermost IRt (located downstream of IS6100) is missing from pRSB225, presumably due to replacement of the DNA immediately adjacent to IS6100 by the macrolide resistance gene mphA, which is located between IS6100 and an IS26. The IS6100-mphA-tniA-IS26 configuration is seen in many hundreds of plasmids in the GenBank nonredundant database, but the configuration with dfrA14 upstream of the IS6100-mphA-tniA-IS26 fragment is relatively rare, present in only 23 sequences in the GenBank nonredundant nucleotide database (last searched May 27, 2019). The integron in pRSB225 does not contain the three additional G nucleotides in the 5′-CS that give rise to P2 and are characteristic of In2 from Tn21 and relatives such as Tn1935 in pZM3. This indicates that the recombination event that gave rise to pRSB225 has likely occurred within intI1 at one end and in IS26 at the other end.
The third event was the introduction of Tn6029 adjacent to Tn4352 in an overlapping configuration sharing an IS26 (Fig. 2B), possibly through IS26-mediated introduction of a translocatable unit, 34 which is known to occur >1,000-fold more frequently than homologous recombination, 35 to generate the transposon Tn6026. Tn6026 still shares the same IS26-ΔtniA boundary as Tn4352 in pZM3. However, this event could also have occurred previously, with the transposon coming in with the internal integron replacement.
Discussion
Although this study originally sought to clarify the relationship between IS1936 and IS26, the sequence of pZM3 ultimately proved valuable to the study of plasmids containing antibiotic resistance determinants and virulence genes. In the genomic era, the study of older historic plasmids is often neglected in favor of large-scale mass sequencing of contemporary antibiotic-resistant isolates. The resistance regions they contain are often very complex. However, over the past few years, a few older plasmids have been sequenced and analyzed (e.g., pIP40a, 36 R16a, 37 R55, 38 R1215, 39 R805a, 40 and R141), and although their resistance regions are also complex, these have provided significant insights into the evolutionary history of plasmids carrying several antibiotic resistance determinants and virulence determinants circulating today. The association between virulence and resistance determinants on conjugative plasmids is not well understood. In particular, older plasmids were often not examined for virulence determinants at the time of their discovery and thus need to be revisited. The aerobactin and sit determinants carried by pZM3 are found on many plasmids of different types, including the FII-FIB virulence plasmids that produce the ColV colicin. 21 These plasmids often carry a suite of virulence determinants and have later acquired resistance determinants.
The antibiotic resistance regions in plasmids are often complex mosaics, comprising fragments of transposons, insertion sequences, integrons, and other mobile genetic elements, interspersed with a wide variety of antibiotic resistance genes. Indeed, many of these regions have arisen through evolution in situ from a single original insertion of a resistance transposon at a particular location (e.g., Refs.24,42–44). The relationship between pZM3 and pRSB225 is a clear example of this process. Recovered 43 years apart, they share a resistance island in the same location (Fig. 2A, B) in an almost identical plasmid backbone. The major difference between the resistance islands results from a recombination event that has exchanged the fragment between intI1 and IS26, introducing additional transposons and a different gene cassette into pRSB225 (Fig. 2B). The specific IS26-tniA boundary seen in pZM3 is a genetic signature that appears in many different resistance islands in different plasmid families. The genetic content on one side of this IS26 often varies, for example, in plasmids carrying sul3-associated integrons 45 and in plasmids carrying a derivative of Tn2670 such as the IncC plasmids, pRMH760, 46 pDGO100, 47 and pB2-1. 29
The large resistance island in pZM3 also shares the same backbone boundary on the right hand side as Tn2670 in NR1 (i.e., 3,845 bp downstream of the FII-1 repA gene), giving rise to the possibility that the resistance island in pZM3 may have initially evolved in situ in NR1 or a related plasmid that contained Tn2670. Movement into a different plasmid backbone is likely to have occurred by homologous recombination at one end in the case of the sul3-containing plasmids related to pCERC3. 45 However, the region adjacent to the NR1-derived backbone segment in pZM3 and pRSB225 is not related to the corresponding segment in NR1.
In NR1, Tn2670 and Tn10 are separated by a 14-kb backbone region that is absent from pZM3 (Fig. 3). The IS1 at one extremity of Tn2670 in NR1 may have deleted the intervening 14-kb backbone region and the IS10 at the end of Tn10. To produce the configuration seen in pZM3, this was followed by a second IS1-mediated event that simultaneously inverted part of the Tn10 remnant and introduced an additional copy of IS1 in inverse orientation (Fig. 3, middle panel). The presence of the separated TSD (denoted by flags in Fig. 3) is characteristic of an IS-mediated inversion event. The structure in pRSB225, in which the Tn10 derivative is in the same orientation as in Tn10 in NR1, could have been generated by homologous recombination-mediated inversion between the two inversely oriented copies of IS1 (Fig. 3, bottom).
It is clear that IS1 is playing an important role, second only to IS26, in shaping the evolution of plasmids and in moving antibiotic resistance genes and virulence determinants in Gram-negative bacteria. It is interesting to note that a number of plasmids belonging to the ColV-ColIa FII-2:FIB-1 group of virulence plasmids 21 share 99.5% nucleotide identity with pZM3 in the region spanning from the IS1 adjacent to sitA to a region ∼2 kb downstream of the FIB-1 repA gene, although the entire aerobactin and sit region between the inversely oriented copies of IS1 is inverted relative to pZM3. The IS1-sit-aerobactin-IS1 transposon-like structure in pZM3 is likely to be mobile as a search of the GenBank nonredundant database (last searched June 27, 2019) revealed that it is found in at least three different genetic contexts, and a previous study demonstrated that both of the copies of IS1 were active and able to mediate the formation of cointegrates. 19 IS1 has also played a role in the rearrangement, and possible mobilization, of remnants of Tn10, and the IS1 copies flanking Tn2670 enhance the ability of this important transposon to mobilize.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work and C.J.H. were supported by the National Health and Medical Research Council, grant no. 1141540.