
Abstract
The study provides data on antibiotic resistance as well as the virulence characteristics of Aeromonas caviae isolated from raw and treated wastewater. The isolates were identified as A. caviae by 16S rRNA gene sequencing. In the analyzed strains, high frequency for the following genes was observed: aac(6′)-Ib-cr, qnrB, and qnrD. The presence of qnrA and ogxB genes was not found in any strain. The higher frequency of the investigated genes was observed in strains from raw wastewater (RW). The strains of A. caviae showed multiple antibiotic resistance evaluated by the disk diffusion method. Multiple antibiotic resistance indices ranged from 0.36 to 0.69. Susceptibility to six heavy metals (Cd+2, Zn+2, Cu+2, Co+2, Mn+2, and Ni+2) was recorded for all the isolates. The order of metal resistance of A. caviae was Co > Cu > Zn > Cd > Ni > Mn. All the strains of A. caviae showed β-hemolytic activity. Enzymes of amylase, cellulase, and lipase were produced by all isolates. Only the strains from RW had the ability to form biofilms and showed motility. The obtained results indicate that wastewater is a potential source and/or reservoir of virulent and multidrug-resistant A. caviae as “high-risk isolates.”
Introduction
A
Aeromonas spp. have demonstrated the increasing levels of quinolone resistance, which is mediated by specific chromosomal and plasmid-mediated quinolone resistance (PMQR) genes. The PMQR determinants include qnr genes, which currently comprise five families: qnrA-, qnrB-, qnrC-, qnrD-, and qnrS-encoding pentapeptide repeat proteins, which provide protection against DNA gyrase and topoisomerase IV. Other PMQR determinants have been reported, such as fluoroquinolone-modifying enzyme aac(6)-Ib-cr, which is a variant of the acetyltransferase of aminoglycosides, and plasmid DNA-encoded efflux pumps qepA and oqxAB.7,8,10–13 PMQR genes not only enhance the adaptability of bacteria to drugs but also facilitate the spread of resistance genes among bacteria from different species.
The wastewater environment provides favorable conditions for the spread of antimicrobial resistance, antimicrobial agents, and antimicrobial resistance of Aeromonas spp.6,14–16 The first report on tracking antimicrobial resistance in Aeromonads in drinking, tap, and waste water and the importance of these bacteria as a vector of resistance in aquatic environments was done by Figueira et al. 17 The authors tried to demonstrate that Aeromonas spp. serve as a vehicle for antibiotic resistance dissemination within the urban water cycle. Wastewater Aeromonads, particularly Aeromonas media and Aeromonas punctate, were confirmed as relevant environmental harbors of quinolone resistance, either chromosomal (gyrA mutation) or plasmid encoded (qnrS and aac(6′)-Ib-cr). The study by Varela et al. 18 highlights the importance of quinolone-resistant Aeromonas spp. as carriers and relevant tracers of antibiotic resistance in hospital and municipal wastewater. The recent publication by Harnisz and Korzeniewska 19 indicated the determinants of antibiotic resistance and virulence in Aeromonas spp. isolated from treated wastewater (TW) and river water samples collected from upstream and downstream of the effluent discharge point. The analyzed isolates were most frequently resistant to beta-lactams, tetracyclines, and aminoglycosides.
The objective of this investigation was to characterize and compare the antibiotic resistance and virulence properties of A. caviae isolated from raw and TW. In particular, the genetic determinants of quinolone resistance were assessed by the detection of plasmid-encoded qnr genes.
Materials and Methods
Sample collection
Samples of the influents (raw wastewater marked as RW) and effluents (treated wastewater marked as TW) were collected from a Polish municipal wastewater treatment plant (WWTP)—geographical coordinates: N 50°5′35.881 and E 19°3′32.202. Detailed description of this WWTP is in publication, Hubeny et al. 20
The wastewater samples were collected into 1 L sterile bottles and transported to the laboratory at 4°C using a cooler box. Bacterial strains were isolated within 24–36 hr of sample collection. Selection and isolation of antibiotic-resistant bacteria were performed in previous studies described in Nowrotek et al. 21 Finally, 38 from the treated and 36 from the raw bacterial strains resistant to antibiotics were isolated from the wastewater.
Identification of antibiotic-resistant bacteria
The genomic DNA from 74 bacteria was isolated with a Roche kit, according to the manufacturer's guidelines. DNA was stored at −20°C until ready for use. The presence of integrons was detected by amplifying an internal fragment of integrase genes intI1 and intI2. The length and sequence of the primers used and the annealing temperature are shown in Table 1. The intI1 gene was detected in all isolates, while intI2 and intI3 were not detected in any of the isolates. More details about this stage were presented in the publication by Nowrotek et al. 21
Qualitative PCR and Sequencing Primers Used in This Study
Antibiotic susceptibility testing
The susceptibility test was performed using the Kirby-Bauer disk diffusion method. 22 One hundred microliters of each bacterial suspension (0.5 density according to the McFarland scale) was placed on Muller-Hinton agar and spread all over the plate. The following antibiotic disks (Oxoid, United Kingdom) at the final concentrations (in brackets) were used: amikacin (AMI: 30 μg/L), amoxicillin (AMO: 30 μg/L), ampicillin (AMP 25 μg/L), azithromycin (AZY 15 μg/L), aztreonam (AZT: 30 μg/L), cefaclor (CEF: 30 μg/L), cefadroxil (CFK: 30 μg/L), cefepime (CFP: 30 μg/L), cefoxitin (CFO: 30 μg/L), ceftaroline (CFR: 5 μg/L), ceftazidime (CFI: 30 μg/L), ciprofloxacin (CYP: 10 μg/L), doripenem (DRI: 10 μg/L), doxycycline (DOX: 30 μg/L), ertapenem (ERA: 10 μg/L), erythromycin (ERT: 30 μg/L), gentamicin (GNT1: 120, GNT2: 200), imipenem (IMP: 10), metronidazole (MET: 50 μg/L), minocycline (MNC: 30 μg/L), mupirocin (MUP: 200 μg/L), nalidixic acid (NAL: 30 μg/L), neomycin (NEO: 30 μg/L), netilmicin (NET: 30 μg/L), nitrofurantoin (NIT: 300 μg/L), norfloxacin (NOR: 10 μg/L), novobiocin (NOV: 30 μg/L), ofloxacin (OFL: 5 μg/L), piperacillin (PIP: 100 μg/L), rifampicin (RYF: 30 μg/L), teicoplanin (TEI: 30 μg/L), ticarcillin (TIC: 75 μg/L), trimethoprim (TRI: 5 μg/L), trimethoprim/sulfamethoxazole (TRI+SMX: 25 μg/L), tobramycin (TOB: 30 μg/L), and vancomycin (VAN: 30 μg/L). The antibiotic discs were placed aseptically onto the surface of the seeded plates. Then, the plates were incubated at 30°C for 24–48 hr. The inhibition zone was measured according to the recommendations of the EUCAST (2018) to estimate the antibiotic resistance. As a quality control strain, Escherichia coli (ATCC 25922) was used. All tests were carried out in triplicate. Multiple antibiotic resistance (MAR) index was calculated, as described by Krumperman. 23 Bacterial isolates were divided into resistant and sensitive based on their resistance profiles.
Polymerase chain reaction and sequence analysis
The oligonucleotide primer sets commercially synthesized by Sigma-Aldrich were used to detect quinolone resistance genes, including aac(6′)-Ib-cr, qnrA, qnrB, qnrD, qnrS, qepA, oqxA, and oqxB. The characteristics of the applied primers and the conditions of PCR reactions are summarized in Table 1. The amplification products were electrophoresed for 15 min at 75 V and 45 min at 90 V in 1 × TBE buffer. Then, they were visualized under a UV trans-illuminator (BioRad) and photographed.
Biofilm formation
The biofilm assay was performed according to a procedure described by Stepanović et al. 24 However, some changes were made to the methodology. The whole part of the experiment was performed based on the methodology presented in the publication by Jałowiecki et al. 25
Motility test
The motility test was performed according to Atkinson et al. 26 with minor modifications, which were presented in the publication by Jałowiecki et al. 25
Detection of biosurfactant production
The method of detecting the production of biosurfactants by bacteria was previously described in Jałowiecki et al. 27
Screening of enzyme activities using plate assays
As part of the research, the following enzymes were analyzed: amylase, cellulase, inulinase, laccase, lipase, and urease. The methodology used to determine the activity of the selected enzymes was previously described in Jałowiecki et al. 27
Determination of hemolytic activity
The strains were tested for hemolytic activity on blood agar base (BioMerieux) containing 5% defibrinated sheep blood with incubation at 30°C for 24–72 hr. Ten microliters of 24-hr culture of isolates was streaked on the blood agar plate. The presence of a clear colorless zone surrounding the colonies indicated hemolytic activity. E. coli (ATCC 25922) and Bacillus subtilis (ATCC 6633) were used as controls.
Testing the minimum inhibitory concentrations of metals
Minimum inhibitory concentrations (MICs) of six metal ions (Cd2+, Zn2+, Ni2+, Cu2+, Co2+, and Mn2+) were determined using the microdilution assay. Sterile 96-well microtiter plates were used in the experiment. The metal chlorides used for the test were CdCl2, ZnCl2, NiCl2, CuCl2, CoCl2, and MnCl2 (Merck, Germany). Metal stock solutions were made in distilled water, sterilized by filtration through cellulose nitrate filters (pore size 0.22 μm and diameter 47 mm, Sartorius), and stored in sterile flasks in the dark at 4°C. The concentrations of metals were chosen on the basis of a preliminary experiment and the literature. All chemicals were of analytical reagent grade.
Each well of the 96-well microplates was filled with 160 μL of sterile Luria-Bertani (LB) broth, 20 μL of 0.5 McFarland bacterial suspension, and 20 μL of metal stock solution of initial concentration 35 mg/mL. The metal concentrations used in the experiment were as follows: 3,500, 1,750, 875, 437.5, 218.75, 109.37, 54.68, 27.34, 13.67, 6.83, 3.4, and 1.7 μg/mL. LB medium without metals and inoculated with test bacteria was used as control. The microplates were incubated at 30°C for 24 and 48 hr. Growth of the bacteria was measured by OD at 600 nm with a microwell plate reader (Eppendorf). The absence of growth was determined by the detection threshold of the plate reader. The MICs were read as the lowest concentrations yielding no visible growth. Susceptibility testing was conducted in triplicate to increase the validity.
Statistical analyses
Statistical analyses were performed using Statistica 13.1 (2018). The principal component analysis (PCA) was performed based on the results obtained for MIC and MAR indices at the significance level p = 0.05.
Results
The criterion used to select strains for further analyses was the presence of the intI1 and intI2 genes. IntI1 gene was detected in 16 isolates and they were subjected to further characterization. The intI2 gene was not found in any of the analyzed strains. The 16S rRNA sequencing analysis revealed that all identified strains belonged to A. caviae. In addition, plasmid DNA was detected in all strains.
Analyzing the obtained results, high frequency of the aac(6′)-Ib-cr gene in the isolates was observed. This gene was found in all analyzed strains, with the exception of the RW_2 strain derived from RW (Table 2). In our study, four of the qnr genes were also analyzed, for example, qnrA, qnrB, qnrD, and qnrS. Two genes, qnrB and qnrD, occurred with equally high frequency; the presence of the qnrD gene was observed in all the analyzed strains, whereas the qnrB gene was present in all strains, except for RW_1 (derived from RW). The presence of qnrA and ogxB genes was not detected in any of the investigated strains. As for qepA gene, it was found only in strains isolated from RW, except for the RW_2 strain where this gene was not observed.
Summary of the Results from the Experimental Work
RW, raw wastewater; TW, treated wastewater; “−” no PCR products or no biofilm producers or generally “no activity”; “+” weak biofilm producer, “++” moderate biofilm producer; “+” presence of PCR products/activity.
Figure 1 presents the phenotypic pattern of antibiotic resistance of A. caviae strains isolated from wastewater. The strains harbored the highest prevalence of resistance to teicoplanin (100%), vancomycin (100%), metronidazole (100%), and ampicillin (100%). Four strains isolated from RW (named RW_2, RW_3, RW_4, and RW_5) were resistant to 15 antibiotics, mostly belonging to fluoroquinolones and β-lactams (Fig. 1). Therefore, the genes conferring resistance to fluoroquinolones were screened. Generally, strains isolated from RW showed higher antibiotic resistance than those isolated from the TW. These strains showed resistance to four different classes of antibiotics.
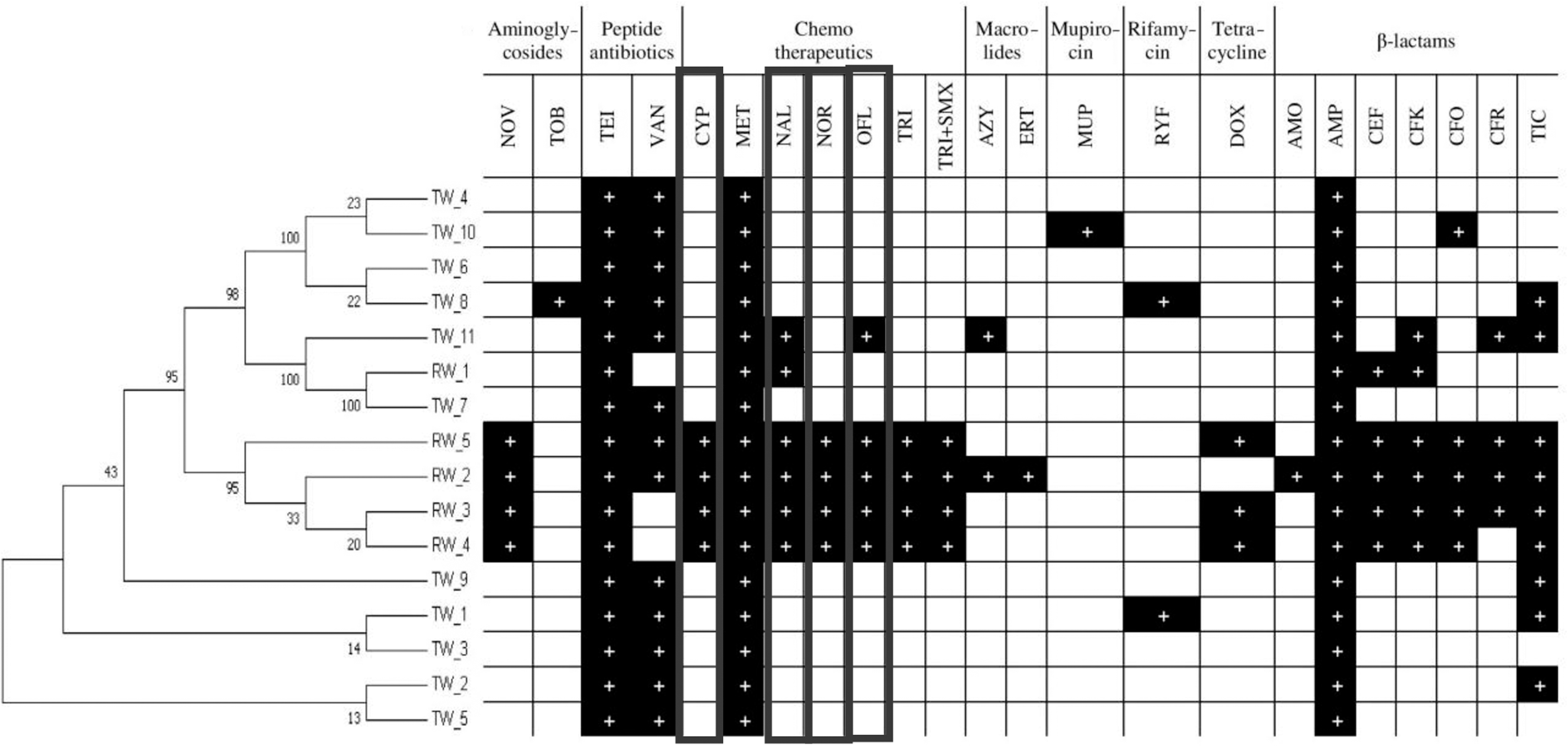
Antibiogram of Aeromonas caviae obtained from the disk diffusion method. Antibiotics belonging to fluoroquinolones are marked by the bold line: amikacin (AMI), amoxicillin (AMO), ampicillin (AMP), azithromycin (AZY), aztreonam (AZT), cefaclor (CEF), cefadroxil (CFK), cefepime (CFP), cefoxitin (CFO), ceftaroline (CFR), ceftazidime (CFI), ciprofloxacin (CYP), doripenem (DRI), doxycycline (DOX), ertapenem (ERA), erythromycin (ERT), gentamicin (GNT), imipenem (IMP), metronidazole (MET), minocycline (MNC), mupirocin (MUP), nalidixic acid (NAL), neomycin (NEO), netilmicin (NET), nitrofurantoin (NIT), norfloxacin (NOR), novobiocin (NOV), ofloxacin (OFL), piperacillin (PIP), rifampicin (RYF), teicoplanin (TEI), ticarcillin (TIC), trimethoprim (TRI), trimethoprim/sulfamethoxazole (TRI+SMX), tobramycin (TOB), and vancomycin (VAN).
The MAR index was determined for each isolate according to Krumperman. 23 The MAR index values of A. caviae isolates are displayed in Fig. 2. All isolates show MAR index values ranging from 0.36 to 0.69. However, some differences can be observed between strains isolated from RW and TW. In the case of strains isolated from RW, the MAR index values ranged from 0.37 to 0.69, with an average value of 0.58. In the TW, the MAR index values ranged from 0.36 to 0.63, with an average value of 0.43.
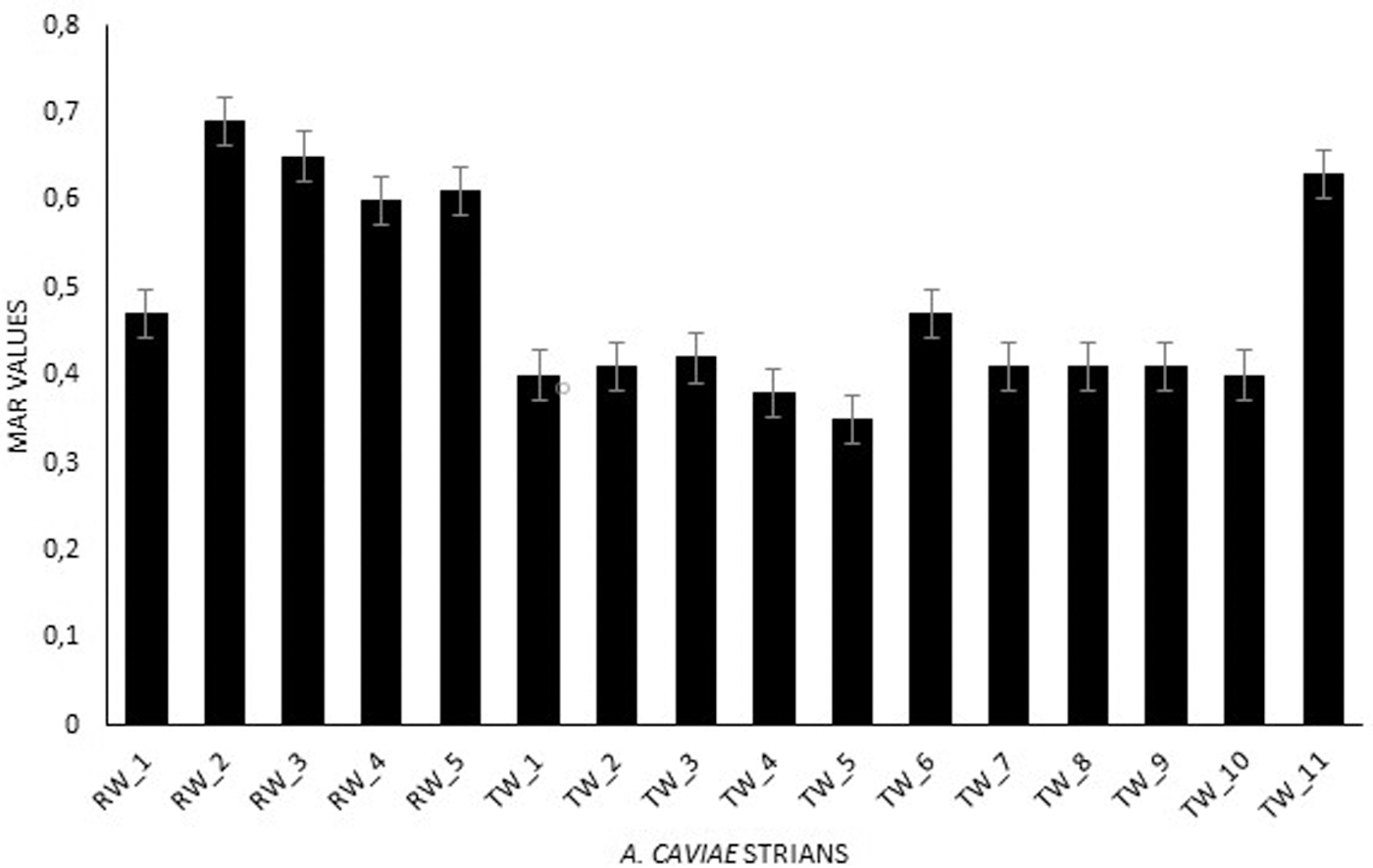
MAR indices of Aeromonas caviae. MAR, multiple antimicrobial resistance.
It is known that heavy metals may enhance selection for antibiotic resistance and vice versa.28,29 In our study, susceptibility to six heavy metals (Cd+2, Zn+2, Cu+2, Co+2, Mn+2, and Ni+2) at the range of concentrations was recorded for all isolates. PCA differentiated all isolates into two groups: those from RW and those from TW (Fig. 3). High resistance of isolates was noted toward the tested metals. However, much higher resistance was recorded for isolates from RW. The MIC averages of the isolates ranged from 0.7 μM for manganese to 26.7 μM for cobalt. Trends in metal resistance varied depending on the isolates from both raw and TW. However, the study showed the same order of metal resistance of A. caviae isolates from both raw and TWs: Co > Cu > Zn > Cd > Ni > Mn. In this study, an attempt was made to determine the relationship between metal MIC values and MAR indices. PCA showed that the strains of A. caviae were divided into two groups (Fig. 3). The arrangement of the vectors indicates that two homogeneous groups were formed. The selection criterion for the first group was the MAR index and for the second, the MIC value. According to the obtained results, no relationship was found between the values of MIC and MAR indices. This was due to the significant distance between points on the chart. The most significant factors affecting the spread of data were the MIC values for Zn and Cu.
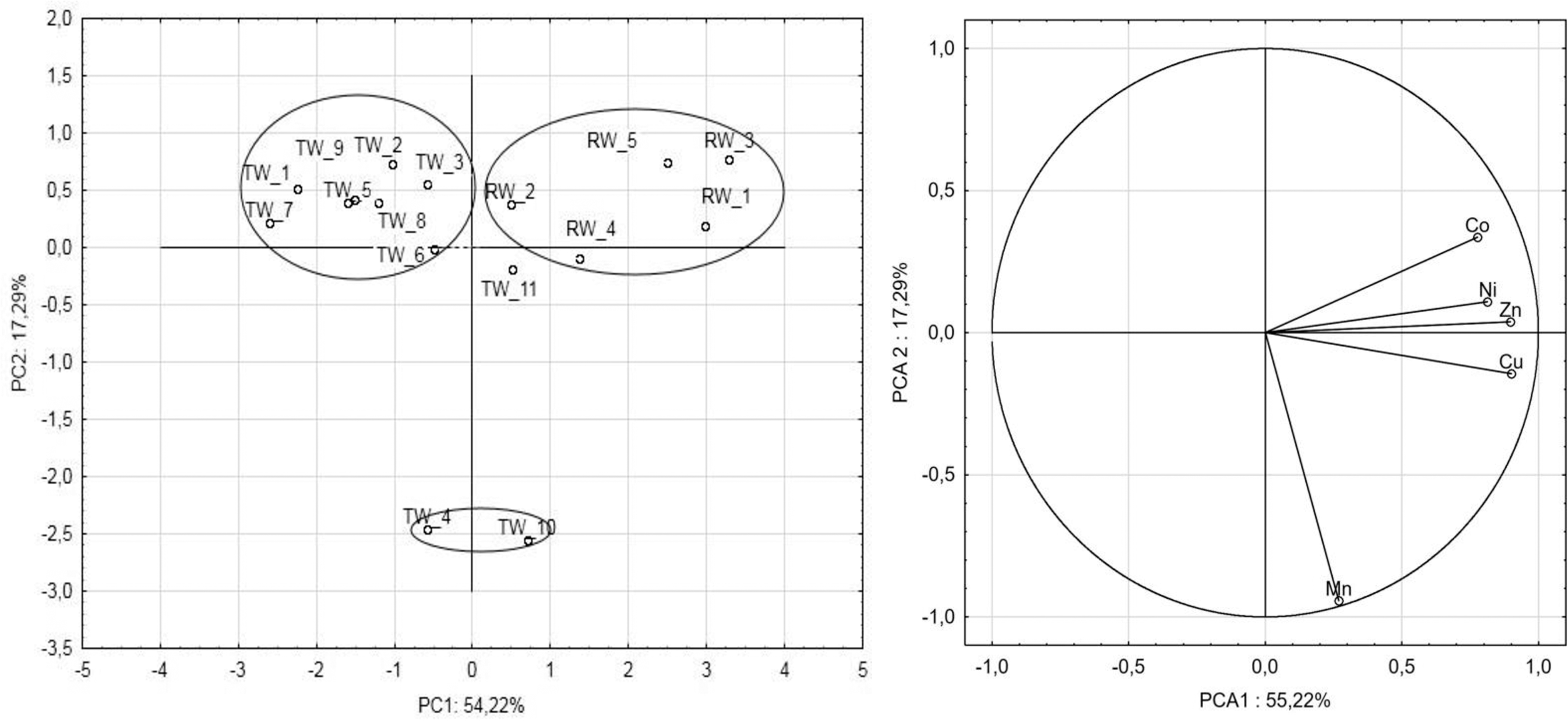
Bidimensional plot of principal component analysis. PC1 and PC2 refer to the first two principal components, accounting for 54.22% and 17.29% of the total variance, respectively.
Also, some phenotypic virulence traits connected with pathogenicity were observed using in vitro tests, including biofilm formation, motility, and enzyme production (Table 2). For most pathogens, secretion of virulence factors and initiation of biofilm formation are important properties during the infections.
A. caviae showed a decrease in surface tension, with values between 46 and 57 mN/m compared with the values of surface tension of water (72 ± 0.23 mN/m) and LB medium (58.34 ± 0.86 mN/m). The reduction of the surface tension confirms that there is production of specific biomolecules having surface-active properties, such as biosurfactants known to cause the lowering of the surface tension. The studied isolates are not biosurfactant producers and no differences were observed between their surface tension values. All the strains of A. caviae showed β-hemolytic activity. A. caviae strains had the ability to produce the selected enzymes. The activities of the following enzymes were determined: urease, laccase, amylase, inulinase, cellulase, and lipase. Among these enzymes, production of three enzymes was not observed in the analyzed strains: urease, laccase, and inulinase. The strains isolated from RW produced the remaining three enzymes, that is, amylase, cellulase, and lipase. More diversified production of these enzymes (amylase, cellulase, and lipase) was observed for strains isolated from TW. Amylase and cellulase production were demonstrated for eight strains, while the lipase enzyme was produced by nine isolated strains. Only the TW_6 strain did not produce any of these enzymes (Table 2).
All strains isolated from RW had the ability to form biofilms. In strains RW_1 and RW_2, this ability was classified as weak, whereas in the case of other strains isolated from RW (RW_3, RW_4, and RW_5), the ability to form biofilm was qualified as moderate. In strains isolated from the TW, weak biofilm capacity was observed in almost all strains, with the exception of two strains, TW_6 and TW_11, which were classified as “non-biofilm producers” (Table 2). Motility was observed only in the strains isolated from RW, whereas in the strains isolated from the TW, motility was noted only in one strain named TW_6. In other strains, motility was not observed.
Discussion
Integrons as mobile elements are involved in the dissemination of antibiotic resistance in the air and water environment. 30 Among three classes of integrons (intI1, intI2, and intI3), class 1 (intI1) is the most common. The integrons were identified in a wide range of Gram-negative bacteria isolated from various sources. 31 In our study, the presence of intI1 was identified in all tested isolates.
In the literature, there is little information about antibiotic resistance genes (ARGs) in A. caviae isolated from WWTP.17,14 ARGs recently found in Aeromonas strains from wastewater encoded resistance to four major groups of antibiotics, that is, the tetracyclines, aminoglycosides, β-lactams, and quinolones. Quinolone resistance, which presents high perspicacity in wastewater environments, is a promising monitoring tag to access antibiotic resistance dissemination.14,17,32–34 Plasmid-borne qnr genes currently constitute five families, qnrA, qnrB, qnrC, qnrD, and qnrS. In our study, two genes, qnrB and qnrD, were determined with equally high frequency. However, the presence of qnrA and ogxB genes was not found in any of the analyzed strains. Generally, higher frequency of qnr genes was observed in strains isolated from RW than in strains from the TW. While analyzing the obtained results, high frequency of the aac(6′)-Ib-cr gene was also observed. This gene was found in all analyzed strains, with the exception of one strain named RW_2 derived from RW. Widespread occurrence of the aac(6′)-Ib-cr gene was also found in other studies. 18 Varela et al. suggest that the gene aac(6′)-Ib-cr in Aeromonas spp. may represent an indicator of multidrug resistance in wastewater and related environments. 18 Furthermore, genes such as qnrS2 and/or aac(6′)-Ib-cr were reported to play a part in the maintenance of resistance levels in the microbial population in the presence of high concentrations of antibiotics.34–36 In other studies, two PMQR genes (aacA-cr and qnrS) were found only in A. media strains. 37 The widespread occurrence of these resistance genes is particularly disadvantageous since quinolones are first-line drugs against Aeromonas-induced infections. 38 Due to this fact, effluents from urban WWTPs are suspected to be one of the most important anthropogenic sources for antibiotics as well as ARGs and antibiotic resistance bacteria spread into the environments.17,39,40
Antibiotic resistance profiles estimated in our study showed that all the isolates were resistant to at least four antibiotics belonging to more than three structural classes and their MAR indices were higher than 0.2. According to the literature, it was assumed that MAR values higher than 0.2 suggested the origin of strains from a high-risk antibiotic-contaminated source. 41 However, the MAR index lower than or equal to 0.2 was considered to indicate original strain resistance to the antibiotic without previous exposure to it. Multidrug-resistant strains were determined as those that had at least simultaneous resistance to three or more classes of antibiotics. All the isolates showed varying degrees of multiple resistant patterns to more than one antibiotic and they had MAR indices above 0.2.
The majority of A. caviae isolates showed high-level resistance to β-lactams (penicillins), trimethoprim, and sulfamethoxazole, and were susceptible to second- and third-generation cephalosporins, carbapenems, macrolides, tetracyclines, fluoroquinolones, and aminoglycosides, and appeared to have similar susceptibility profiles. Most of the isolates were resistant to antibiotics that have been extensively used in hospitals and veterinary or agriculture sectors, contributing to a high level of resistance in bacteria, especially in widely occurring Aeromonas species. 42
Metal and antibiotic resistance are the most common features that bacteria gain in the environments contaminated by metals and/or antibiotics.28,43 Some authors suggest that the presence of metal contamination in natural environments can play a role in the maintenance and proliferation of antibiotic resistance.44–47 Many metal-antibiotic correlations have been observed in samples retrieved from heavy metal-contaminated environments, and cross-resistance mechanisms have been confirmed.46–48 Although there are a lot of data on metal-antibiotic correlations, the mechanisms of the relationship remain scant. It is expected that in environments with a high concentration of metals, bacteria activate their resistance mechanisms, which are selected within the population and serve as an important indicator of environmental contamination. As a whole, our study implies relatively high antimicrobial and heavy metal resistance in A. caviae isolated from wastewater, and this relationship was observed mainly in RW. Trends in metal resistance varied depending on the isolates from both raw and TW. Nevertheless, a similar order of metal resistance of A. caviae isolated from raw and TW was observed: Co > Cu > Zn > Cd > Ni > Mn. However, in contrast to our observations, the study by Matyar et al. 49 showed a different order of resistance for Aeromonas isolates: Cu > Mn > Cr > Cd = Zn > Pb. Therefore, the wastewater should not be underrated as a source of multidrug- and heavy metal-resistant Aeromonads, especially in the context of potential public health implications. The mobile genetic elements such as integrons or plasmids containing genes of metal or antibiotic resistance are the cause of horizontal transfer between different bacteria.
To the best of our knowledge, there is no report on the ability of A. caviae to produce biosurfactants, and our results confirm this fact. However, several studies on the production of biosurfactants by various Aeromonas spp. have been reported. The study by Ilori et al. 50 shows that oil degrading Aeromonas spp. produces a glycolipid that probably represents a new type of biosurfactant with a strong emulsifying ability. Priya 51 described six Aeromonas species with the hydrocarbon biodegradation and emulsifying abilities. The strains also lowered the surface tension between 20 and 30 mN/m. In the study by Kamal et al., 52 Aeromonas salmonicida reduced the surface tension to the value of 34.5 mN/m. Production of biosurfactants by A. salmonicida was confirmed by the following methods: calculation of the emulsification index, methylene blue test, thin-layer chromatography analysis, hemolysis, and cell hydrophobicity tests. This result, which was obtained in the case of all applied methods, confirmed that the A. salmonicida strain had the highest potential in the production of biosurfactants. 53 Also, the analysis of the chemical structure by Fourier transform infrared (FT-IR) spectroscopy confirmed that the functional groups of biosurfactants produced by A. salmonicida were lipids.
Hemolytic proteins are commonly isolated from pathogenic bacteria, and β-hemolysins are classified as indirect bacterial virulence factors. A. caviae has recently been recognized as an important enteropathogen, and its hemolysin is purported to be one of the virulence factors. Singh and Sanyal demonstrated that most of these β-hemolytic strains were either A. hydrophila or Aeromonas sobria, but rarely A. caviae. 54 Similar results were presented by Nakano et al., where a majority of the A. hydrophila and A. sobria isolates were highly hemolytic, whereas only 11% of the A. caviae isolates were capable of lysing sheep erythrocytes. 55 In contrast, all strains of A. caviae tested in our study showed β-hemolytic activity. Karunakarani and Devi found that A. caviae strain named NRRL B 966 produced an extracellular β-hemolytic toxin. To characterize this hemolytic activity, the bacterial kinetics of the growth and hemolytic toxin production were determined. 56 The hemolytic toxin activity steadily increased in the medium with time parallelly to growth and reached a maximum activity in the stationary phase. Nacescu et al. 57 postulated a correlation between the pathogenic potential and the hemolytic activity of Aeromonas species. 58 Sometimes, the hemolytic activity of A. caviae was not observed. In this case, it might be the potential for virulence, but the toxin and/or hemolytic activity is not detected phenotypically because of underexpression or nonexpression of the toxin/hemolytic genes related to growth conditions.
Our analysis revealed that lipases were the enzyme most frequently produced by A. caviae. As many as 87% strains showed lipase activity. Among them, there were all those isolated from RW. Also, around 80% strains showed amylase and cellulase activities. The lipases (or triacylglycerol hydrolases) as virulence factors interact with human leukocytes and modulate host immune systems. Lipases are a specific class of enzymes, which are very important in hydrolysis of triglycerides to glycerol and free fatty acids. However, Aeromonas lipases belonging to the superfamily of microbial lipases with significant sequence homology differ in substrate specificity, thermostability, and pH optima. 59 Lipases are typical enzymes mainly produced by pathogenic organisms. As reported, they also play a major role in pathogenicity toward warm-blooded animals. 60 In the case of proteases, their activity contributes to the process of bacterial multiplication, as well as acts on host nutrient sequestration and immune response. 61 The activity of another enzyme—amylase—was also observed in human infections caused by Aeromonas strains, but there is still insufficient evidence that its activity was related to the infection. 62
In medicine, it has been estimated that 65% of infections are biofilm associated. As estimated, these biofilm infections are 10–1,000 times more resistant to the effects of antimicrobial agents. Biofilm formation is considered to be one of the vital virulence factors, aiding bacterial colonization by cell adhesion to epithelial cells and intestinal villi, reducing bacterial sensitivity to antimicrobial agents, and reducing recognition of bacteria by the immunologic system.63,64 Bechet and Blondeau 65 observed that A. caviae were capable of forming biofilms on the glass surface of culture flasks in nutrient broth. Our results revealed that the A. caviae isolated from wastewater were capable of biofilm forming with various degrees of intensity. The strains isolated from RW were stronger biofilm producers than those from the TW. The intensity of biofilm production depended on the source of the Aeromonas species. For example, Aeromonas isolates from a river can form the biofilm with a high percentage of weak biofilm producers and strong producers trailing as the second occurrence.66,67 The study of biofilm formation by a variety of A. caviae isolated from different wastewater provides more relevant information on the behavior of species from diverse sources.
Motility is known to facilitate rapid colonization of surfaces and is linked to virulence factors such as enzyme production, biofilm formation, antimicrobial resistance, and pathogenicity. 68 Among Aeromonas species, two major motility types were observed according to the sort of flagellum produced—swimming motility is associated with liquid environments 69 and promoted mainly by the polar flagellum, while the lateral type of motility promotes swarming movement on solid surfaces. 59 The majority of the isolates in this study showed swimming motility. In our study, a significant positive correlation was observed between swimming motility and biofilm formation in isolates from RW. In literature, nonmotile Aeromonas spp. with optimal growth temperature between 22°C and 25°C are also discussed, as well as mesophilic species with motility, where the optimal growth temperature was between 35°C and 37°C.61,70 Mesophilic Aeromonas express a single polar flagellum for swimming in all culture conditions and produce lateral flagella for swarming on solid media. The role of motility and other surface and virulence properties requires further investigation, mainly genetic analysis of the whole genomes.
Conclusions
This study is only one of the few studies that describe both the antibiotic and metal resistance and virulence factors in A. caviae strains isolated from wastewater. The World Health Organization lists Aeromonas spp. as a potential water-borne pathogen. This study gives new data and perspectives of research on such virulence factors as, for example, enzyme activities, mobility, biofilm formation, and spread of antibiotic resistance. The pathogenicity of A. caviae can be characterized by both phenotypic virulence traits and antibiotic resistance. Our results imply the existence of potentially pathogenic and multiple antibiotic-resistant A. caviae strains in wastewater, especially in the TW, indicating that wastewater is a quite significant source and/or reservoir of antibiotic resistance determinants.
In conclusion, the carried out study shows that in the wastewater impacted by human activities, antibiotic-and heavy metal-resistant bacteria with virulence properties can be found, posing a risk to public health. The study indicates that WWTPs represent a source and/or reservoir of multidrug-resistant genetic elements carrying new associations of resistance genes into the receiving water bodies.
Authors' Contributions
All authors contributed equally to this study. All authors read and approved the final version of the article.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was financed by the National Science Center (project no: 2017/26/M/NZ9/00071).